
Anaerobic Membrane Bioreactor for Continuous Lactic Acid Fermentation




Abstract
:1. Introduction
2. Materials and Methods
2.1. Ceramic Membranes
2.2. Bacterial Culture
2.3. Batch Fermentation
2.4. Optical Sensor
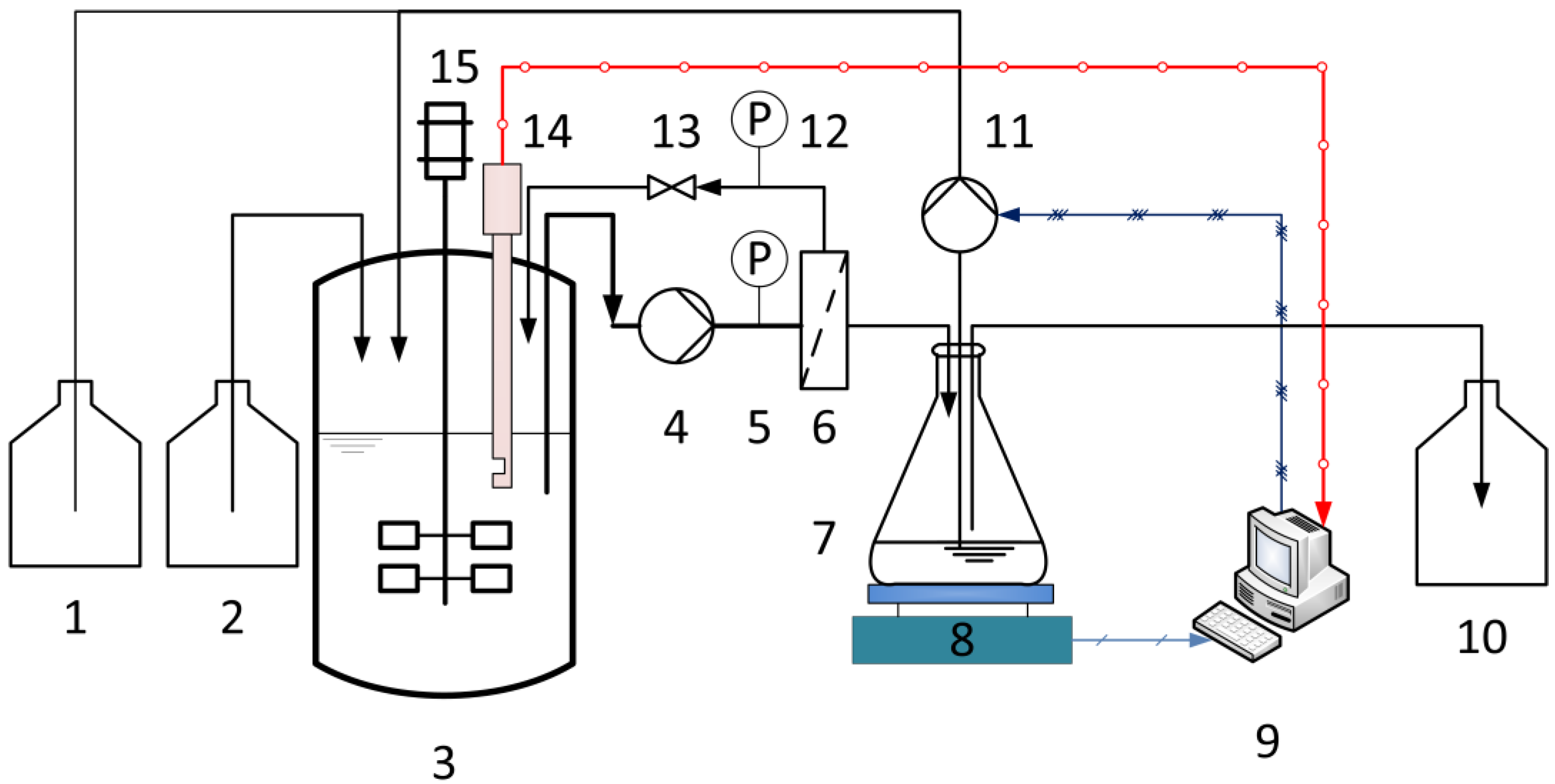
2.5. Experimental Equipment
2.6. Analytical Method
2.7. Calculation
3. Results
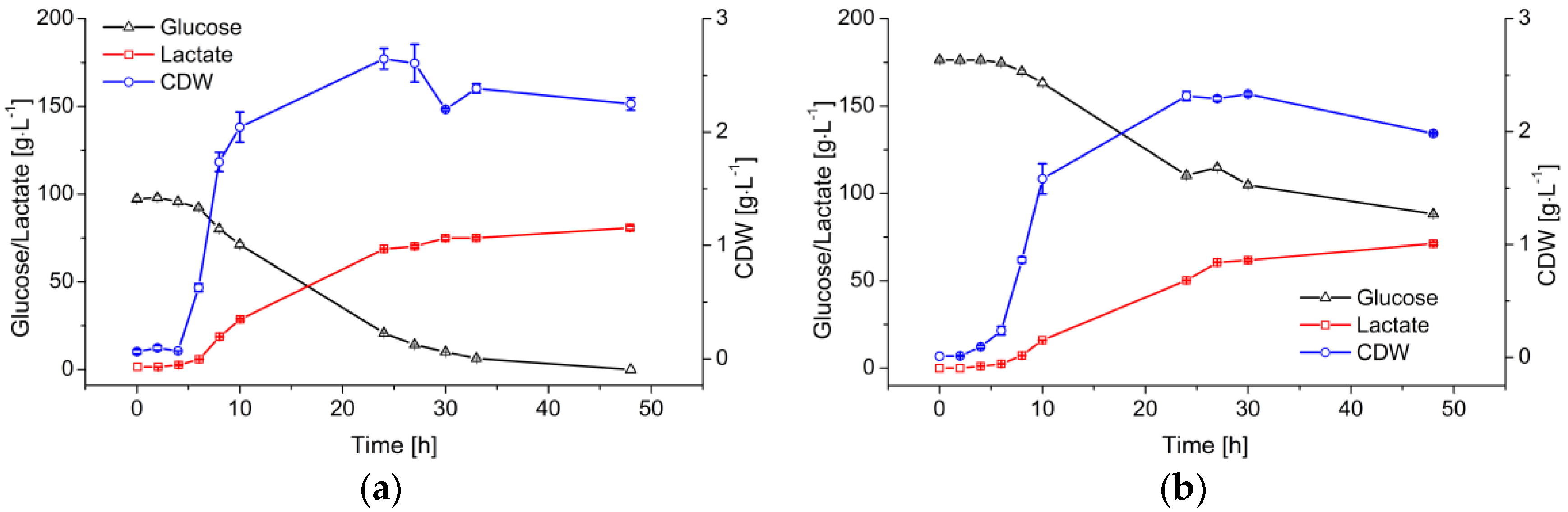
3.1. Batch Fermentation for LA Production
3.2. Filtration Performance of Ceramic Membranes
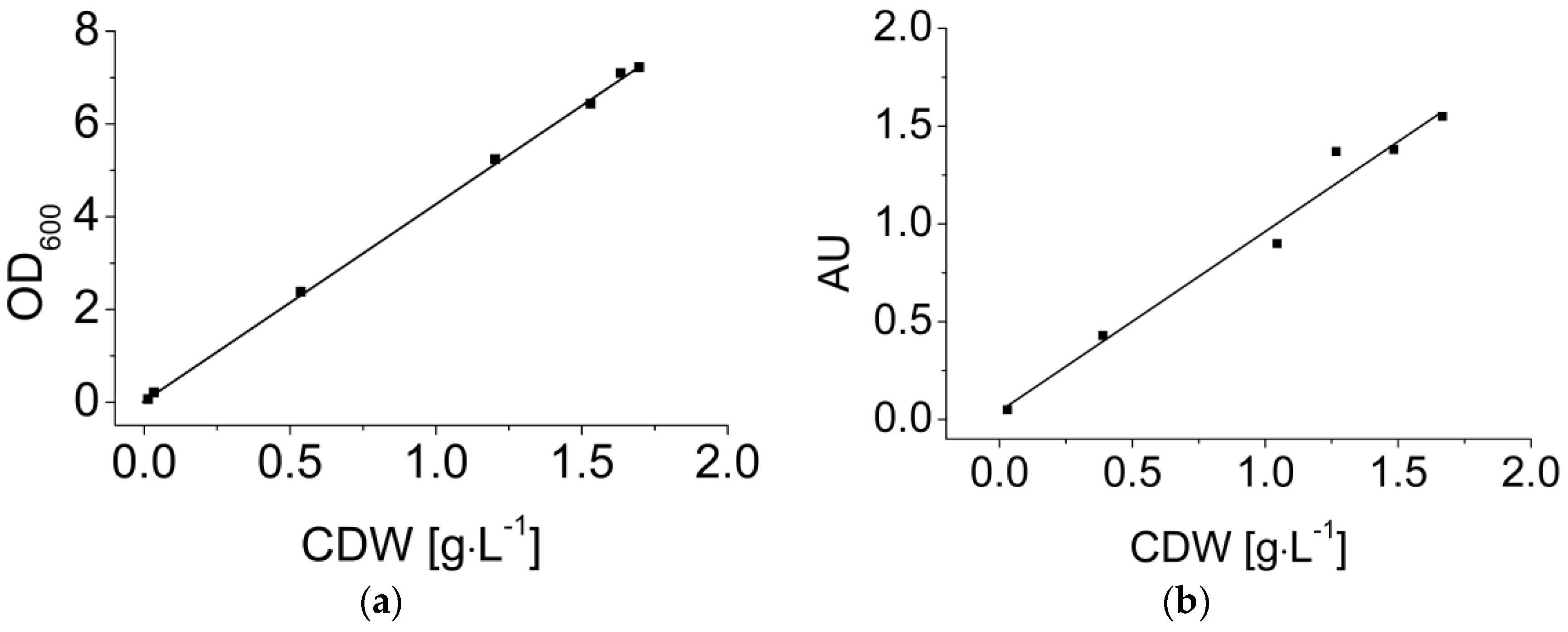
3.3. Dynamic Calibration of the EXcell Sensor
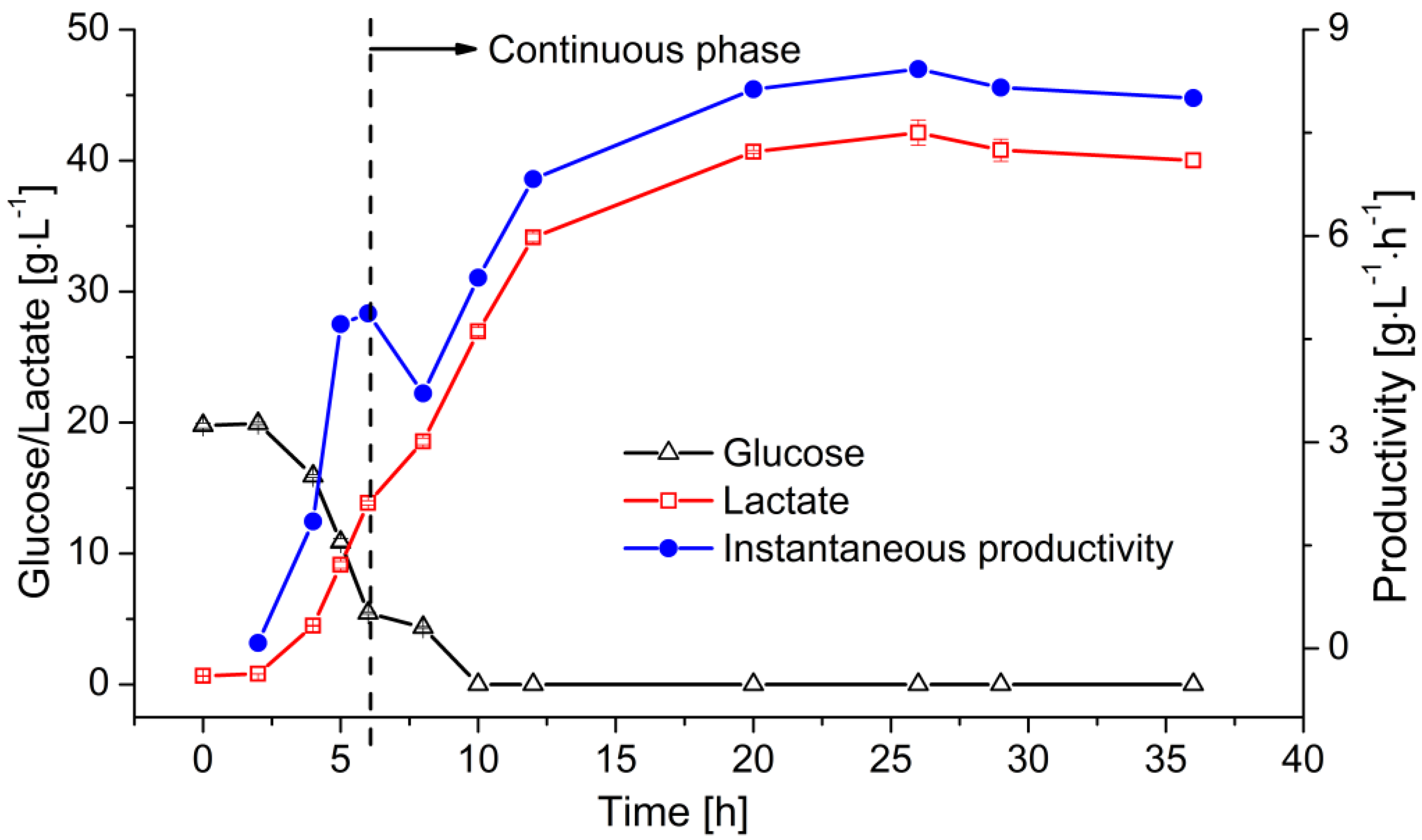
3.4. Continuous Fermentation of LA in the MBR
4. Discussion
4.1. Batch Fermentation
4.2. Filtration of the Fermentation Broth
4.3. Dynamic Calibration of the Optical Sensor
4.4. Continuous Fermentation of LA in the MBR
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AU | Arbitrary units |
| CDW | Cell dry weight |
| CFV | Crossflow velocity |
| F | Flow rate |
| LA | Lactic acid |
| μ | Specific growth rate |
| M | Mass of biomass represented by offline measurements |
| MBR | Membrane bioreactor |
| MWCO | Molecular weight cutoff |
| MO | Microorganism |
| N | Mass of biomass represented by online measurements |
| NIR | Near infrared |
| ODx | Optical density at λ = x nm |
| P | Pressure |
| PLA | Polylactic acid |
| Q | Productivity of lactic acid |
| RO | Reverse osmosis |
| t | Time |
| T | Temperature |
| TMP | Transmembrane pressure |
References
- Le-Clech, P.; Chen, V.; Fane, T.A. Fouling in membrane bioreactors used in wastewater treatment. J. Membr. Sci. 2006, 284, 17–53. [Google Scholar] [CrossRef]
- Smith, C.V.; Di Gregorio, D.; Talcott, R.M. (Eds.) The use of ultrafiltration membranes for activated sludge separation. In Proceedings of the 24th Annual Purdue Industrial Waste Conference, West Lafayette, IN, USA, 6–8 May 1969. [Google Scholar]
- Iorhemen, O.T.; Hamza, R.A.; Tay, J.H. Membrane Bioreactor (MBR) Technology for Wastewater Treatment and Reclamation: Membrane Fouling. Membranes 2016, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Chae, S.-R.; Drews, A.; Kraume, M.; Shin, H.-S.; Yang, F. Recent advances in membrane bioreactors (MBRs): Membrane fouling and membrane material. Water Res. 2009, 43, 1489–1512. [Google Scholar] [CrossRef] [PubMed]
- Mutamim, N.S.A.; Noor, Z.Z.; Hassan, M.A.A.; Yuniarto, A.; Olsson, G. Membrane bioreactor: Applications and limitations in treating high strength industrial wastewater. Chem. Eng. J. 2013, 225, 109–119. [Google Scholar] [CrossRef]
- Judd, S. The status of membrane bioreactor technology. Trends Biotechnol. 2008, 26, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Hiasa, M.; Mahmood, T.; Matsuo, T. Direct Solid-Liquid Separation Using Hollow Fiber Membrane in an Activated Sludge Aeration Tank. Water Sci. Technol. 1989, 21, 43–54. [Google Scholar]
- Busse, N.; Kraume, M.; Czermak, P. Optimal permeate flux for an enzymatic oxidation of technical lignins in a membrane reactor. Sep. Sci. Technol. 2016, 52, 374–380. [Google Scholar] [CrossRef]
- Ebrahimi, M.; Busse, N.; Kerker, S.; Schmitz, O.; Hilpert, M.; Czermak, P. Treatment of the Bleaching Effluent from Sulfite Pulp Production by Ceramic Membrane Filtration. Membranes 2016, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- Ur Rehman, A.; Kovacs, Z.; Quitmann, H.; Ebrahimi, M.; Czermak, P. Enzymatic production of fructo-oligosaccharides from inexpensive and abundant substrates using a membrane reactor system. Sep. Sci. Technol. 2016, 51, 1537–1545. [Google Scholar] [CrossRef]
- Carstensen, F.; Klement, T.; Büchs, J.; Melin, T.; Wessling, M. Continuous production and recovery of itaconic acid in a membrane bioreactor. Bioresour. Technol. 2013, 137, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Colomban, A.; Roger, L.; Boyaval, P. Production of propionic acid from whey permeate by sequential fermentation, ultrafiltration, and cell recycling. Biotechnol. Bioeng. 1993, 42, 1091–1098. [Google Scholar] [CrossRef] [PubMed]
- Pal, P.; Dey, P. Process intensification in lactic acid production by three stage membrane integrated hybrid reactor system. Chem. Eng. Process. 2013, 64, 1–9. [Google Scholar] [CrossRef]
- Lu, Z.; Wei, M.; Yu, L. Enhancement of pilot scale production of l(+)-lactic acid by fermentation coupled with separation using membrane bioreactor. Process. Biochem. 2012, 47, 410–415. [Google Scholar] [CrossRef]
- Subramanian, M.R.; Talluri, S.; Christopher, L.P. Production of lactic acid using a new homofermentative Enterococcus faecalis isolate. Microb. Biotechnol. 2015, 8, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Wee, Y.-J.; Kim, J.-N.; Ryu, H.-W. Biotechnological Production of Lactic Acid and Its Recent Applications. Food Technol. Biotechnol. 2006, 44, 163–172. [Google Scholar]
- Rosenberg, M.; Rebroš, M.; Krištofíková, L.; Malátová, K. High Temperature Lactic Acid Production by Bacillus coagulans Immobilized in LentiKats. Biotechnol. Lett. 2005, 27, 1943–1947. [Google Scholar] [CrossRef] [PubMed]
- Castillo Martinez, F.A.; Balciunas, E.M.; Salgado, J.M.; Domínguez González, J.M.; Converti, A.; de Souza Oliveira, R.P. Lactic acid properties, applications and production: A review. Trends Food Sci. Technol. 2013, 30, 70–83. [Google Scholar] [CrossRef]
- Quitmann, H.; Fan, R.; Czermak, P. Acidic organic compounds in beverage, food and feed production. Adv. Biochem. Eng. Biotechnol. 2014, 143, 91–141. [Google Scholar] [PubMed]
- Chahal, S.P.; Starr, J.N. (Eds.) Lactic Acid, Ullmann’s Encyclopedia of Industrial Chemistry; Wiley-VCH, GmbH & Co. KGaA: Weinheim, Germany, 2006; pp. 1–8. [Google Scholar]
- Narayanan, N.; Roychoudhury, P.K.; Srivastava, A. l(+) lactic acid fermentation and its product polymerization. Electron. J. Biotechnol. 2004, 7, 167–179. [Google Scholar]
- Zhao, B.; Wang, L.; Li, F.; Hua, D.; Ma, C.; Ma, Y.; Xu, P. Kinetics of d-lactic acid production by Sporolactobacillus sp. strain CASD using repeated batch fermentation. Bioresour. Technol. 2010, 101, 6499–6505. [Google Scholar] [CrossRef] [PubMed]
- De Vecchi, E.; Drago, L. Lactobacillus sporogenes or Bacillus coagulans: Misidentification or Mislabelling? Int. J. Probiotics Prebiotics 2006, 1, 3–10. [Google Scholar]
- Xu, P.; Wang, L.; Zhao, B.; Ma, C.; Su, F.; Tao, F.; Tang, H. Bacillus coagulans Strains and their Applications in l-Lactic Acid Production. U.S. Patent No. 8,492,127, 23 July 2013. Available online: http://www.google.com/patents/US8492127 (accessed on 15 January 2017).
- Michelson, T.; Kask, K.; Jõgi, E.; Talpsep, E.; Suitso, I.; Nurk, A. l(+)-Lactic acid producer Bacillus coagulans SIM-7 DSM 14043 and its comparison with Lactobacillus delbrueckii ssp. lactis DSM 20073. Enzyme Microb. Technol. 2006, 39, 861–867. [Google Scholar] [CrossRef]
- Walton, S.L.; Bischoff, K.M.; Heiningen, A.R.P.; Walsum, G.P. Production of lactic acid from hemicellulose extracts by Bacillus coagulans MXL-9. J. Ind. Microbiol. Biotechnol. 2010, 37, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Luedeking, R.; Piret, E.L. A kinetic study of the lactic acid fermentation. Batch process at controlled pH. Biotechnol. Bioeng. 1959, 1, 393–412. [Google Scholar] [CrossRef]
- Zacharof, M.-P.; Lovitt, R.W. Modelling and simulation of cell growth dynamics, substrate consumption, and lactic acid production kinetics of Lactococcus lactis. Biotechnol. Bioprocess. Eng. 2013, 18, 52–64. [Google Scholar] [CrossRef]
- Nandasana, A.D.; Kumar, S. Kinetic modeling of lactic acid production from molasses using Enterococcus faecalis RKY1. Biochem. Eng. J. 2008, 38, 277–284. [Google Scholar] [CrossRef]
- Ou, M.S.; Ingram, L.O.; Shanmugam, K.T. l(+)-Lactic acid production from non-food carbohydrates by thermotolerant Bacillus coagulans. J. Ind. Microbiol. Biotechnol. 2011, 38, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Ye, L.; Wu, J.C. Efficient production of l-lactic acid by newly isolated thermophilic Bacillus coagulans WCP10-4 with high glucose tolerance. Appl. Microbiol. Biotechnol. 2013, 97, 4309–4314. [Google Scholar] [CrossRef] [PubMed]
- Dey, P.; Pal, P. Direct production of l(+) lactic acid in a continuous and fully membrane-integrated hybrid reactor system under non-neutralizing conditions. J. Membr. Sci. 2012, 389, 355–362. [Google Scholar] [CrossRef]
- Xu, G.-Q.; Chu, J.; Wang, Y.-H.; Zhuang, Y.-P.; Zhang, S.-L.; Peng, H.-Q. Development of a continuous cell-recycle fermentation system for production of lactic acid by Lactobacillus paracasei. Process. Biochem. 2006, 41, 2458–2463. [Google Scholar] [CrossRef]
- Cirilo, N.H.; Toshiyuki, M.; Genta, K.; Kenji, S.; Ayaaki, I. Synchronized fresh cell bioreactor system for continuous l-(+)-lactic acid production using Lactococcus lactis IO-1 in hydrolysed sago starch. J. Biosci. Bioeng. 2002, 93, 281–287. [Google Scholar]
- Fan, R.; Ebrahimi, M.; Quitmann, H.; Czermak, P. Lactic acid production in a membrane bioreactor system with thermophilic Bacillus coagulans: Fouling analysis of the used ceramic membranes. Sep. Sci. Technol. 2015, 48, 2177–2189. [Google Scholar]
- Carrère, H.; Blaszkow, F.; de Balmann, H.R. Modelling the clarification of lactic acid fermentation broths by cross-flow microfiltration. J. Membr. Sci. 2001, 186, 219–230. [Google Scholar] [CrossRef]
- Gao, T.; Wong, Y.; Ng, C.; Ho, K. l-lactic acid production by Bacillus subtilis MUR1. Bioresour. Technol. 2012, 121, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Hu, G.; Pan, L.; Wang, Z.; Zhou, Y.; Wang, Y.; Ruan, Z.; He, M. Highly efficient production of optically pure l-lactic acid from corn stover hydrolysate by thermophilic Bacillus coagulans. Bioresour. Technol. 2016, 219, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Zhou, X.; Hudari, M.S.B.; Li, Z.; Wu, J.C. Highly efficient production of l-lactic acid from xylose by newly isolated Bacillus coagulans C106. Bioresour. Technol. 2013, 132, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Vincent-Vela, M.C.; Cuartas-Uribe, B.; Álvarez-Blanco, S.; Lora-García, J. Analysis of an ultrafiltration model: Influence of operational conditions. Desalination 2012, 284, 14–21. [Google Scholar] [CrossRef]
- Hwang, K.J.; Wang, C.Y. Microfiltration characteristics of Bacillus subtilis fermentation broths. J. Taiwan Inst. Chem. Eng. 2012, 43, 347–353. [Google Scholar] [CrossRef]
- Ude, C.; Schmidt-Hager, J.; Findeis, M.; John, G.T.; Scheper, T.; Beutel, S. Application of an online-biomass sensor in an optical multisensory platform prototype for growth monitoring of biotechnical relevant microorganism and cell lines in single-use shake flasks. Sensors 2014, 14, 17390–17405. [Google Scholar] [CrossRef] [PubMed]
- Giorno, L.; Chojnacka, K.; Donato, L.; Drioli, E. Study of a Cell-Recycle Membrane Fermentor for the Production of Lactic Acid by Lactobacillus bulgaricus. Ind. Eng. Chem. Res. 2002, 41, 433–440. [Google Scholar] [CrossRef]
- Fan, R.; Ebrahimi, M.; Quitmann, H.; Czermak, P. Lactic acid production in a membrane bioreactor system with thermophilic Bacillus coagulans: Online monitoring and process control using an optical sensor. Sep. Sci. Technol. 2016, 52, 1–12. [Google Scholar] [CrossRef]
- Suripto, D.Y.; Abdul, G.; Takao, K. Effect of Product Inhibitions on l-Lactic Acid Fermentation from Fresh Cassava Roots in Tofu Liquid Waste by Streptococcus bovis. J. Ferment Bioeng. 2008, 9, 59–65. [Google Scholar]
- Zhang, Y.; Chen, X.; Luo, J.; Qi, B.; Wan, Y. An efficient process for lactic acid production from wheat straw by a newly isolated Bacillus coagulans strain IPE22. Bioresour. Technol. 2014, 158, 396–399. [Google Scholar] [CrossRef] [PubMed]
- Ahring, B.K.; Traverso, J.J.; Murali, N.; Srinivas, K. Continuous fermentation of clarified corn stover hydrolysate for the production of lactic acid at high yield and productivity. Biochem. Eng. J. 2016, 109, 162–169. [Google Scholar] [CrossRef]
- Yuwono, S.D.; Mulyono; Widiarto, S.; Hadi, S.; Kokugan, T. Improvement of Lactic Acid Production from Cassava by Streptococcus bovis Using Two-Stages Membrane Bioreactor. Asian J. Chem. 2014, 26, 6249–6252. [Google Scholar]
- Van Hecke, W.; Verhoef, S.; Groot, W.; Sarić, M.; van de Bunt, B.; de Haan, A.; de Wever, H. Investigation of lactate productivity in membrane bioreactors on C5/C6 carbohydrates. J. Membr. Sci. 2017, 528, 336–345. [Google Scholar] [CrossRef]
- Tang, J.; Wang, X.C.; Hu, Y.; Ngo, H.H.; Li, Y. Dynamic membrane-assisted fermentation of food wastes for enhancing lactic acid production. Bioresour. Technol. 2017, 234, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Wang, X.; Hu, Y.; Zhang, Y.; Li, Y. Lactic acid fermentation from food waste with indigenous microbiota: Effects of pH, temperature and high OLR. Waste Manag. 2016, 52, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Marques, S.; Matos, C.T.; Gírio, F.M.; Roseiro, J.C.; Santos, J. Lactic acid production from recycled paper sludge: Process intensification by running fed-batch into a membrane-recycle bioreactor. Biochem. Eng. J. 2017, 120, 63–72. [Google Scholar] [CrossRef]
- Mimitsuka, T.; Sawai, K.; Kobayashi, K.; Tsukada, T.; Takeuchi, N.; Yamada, K.; Ogino, H.; Yonehara, T. Production of d-lactic acid in a continuous membrane integrated fermentation reactor by genetically modified Saccharomyces cerevisiae: Enhancement in d-lactic acid carbon yield. J. Biosci. Bioeng. 2015, 119, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.K.; Ryu, H.W.; Oh, H.; Kim, M.; Wee, Y.J. Cell-recycle continuous fermentation of Enterococcus faecalis RKY1 for economical production of lactic acid by reduction of yeast extract supplementation. J. Microbiol. Biotechnol. 2014, 24, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Min-tian, G.; Koide, M.; Gotou, R.; Takanashi, H.; Hirata, M.; Hano, T. Development of a continuous electrodialysis fermentation system for production of lactic acid by Lactobacillus rhamnosus. Process. Biochem. 2005, 40, 1033–1036. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Y.; Zhang, X.; Xu, T. In situ combination of fermentation and electrodialysis with bipolar membranes for the production of lactic acid: Operational compatibility and uniformity. Bioresour. Technol. 2012, 125, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Li, W.; Fan, Y.; Xing, W. Integrated Membrane Process for the Purification of Lactic Acid from a Fermentation Broth Neutralized with Sodium Hydroxide. Ind. Eng. Chem. Res. 2013, 52, 2412–2417. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Operation Mode | MO 1 | Substrate Conc. 2 (g·L−1) | LA Conc. (g·L−1) | Dilution Rate (h−1) | Yield (g·g−1) | Productivity (g·L−1·h−1) | Ref. |
|---|---|---|---|---|---|---|---|
| Continuous with total cell recycle | B. coagulans PS5 | 50 | 42 | 0.2 | 0.84 | 8.4 | This work |
| Batch | B. coagulans WCP 10-4 | 240 | 210 | – | 0.95 | 3.5 | [31] |
| Fed-batch | B. coagulans C106 | 120 + 80 + 60 | 215.7 | – | 0.95 | 4.0 | [39] |
| Membrane integrated repeated batch | B. coagulans IPE22 | 60 | 56.5 | – | 0.96 | 2.35 | [46] |
| Membrane integrated repeated batch | B. coagulans NBRC 12714 | 110 | 91.4 | – | 0.90 | 3.9 | [38] |
| Continuous with cell recycle | B. coagulans NBRC 12714 | 100 | 92 | 0.15 | 0.92 | 13.8 | [38] |
| Continuous with cell recycle | B. coagulans AD | 51 | 35.2 | 0.167 | 0.95 | 3.69 | [47] |
| Continuous with cell recycle in two-stages MBR | S. bovis | 30 | 20.1 | 0.25 | 0.67 | 5.03 | [48] |
| Continuous in MBR with cell bleeding | B. coagulans ATCC 23498 | 99.7 | 85.4 | 0.357 | 0.86 | 30.5 | [49] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, R.; Ebrahimi, M.; Czermak, P. Anaerobic Membrane Bioreactor for Continuous Lactic Acid Fermentation. Membranes 2017, 7, 26. https://doi.org/10.3390/membranes7020026
Fan R, Ebrahimi M, Czermak P. Anaerobic Membrane Bioreactor for Continuous Lactic Acid Fermentation. Membranes. 2017; 7(2):26. https://doi.org/10.3390/membranes7020026
Chicago/Turabian StyleFan, Rong, Mehrdad Ebrahimi, and Peter Czermak. 2017. "Anaerobic Membrane Bioreactor for Continuous Lactic Acid Fermentation" Membranes 7, no. 2: 26. https://doi.org/10.3390/membranes7020026
APA StyleFan, R., Ebrahimi, M., & Czermak, P. (2017). Anaerobic Membrane Bioreactor for Continuous Lactic Acid Fermentation. Membranes, 7(2), 26. https://doi.org/10.3390/membranes7020026