A Comprehensive Review of Our Current Understanding of Red Blood Cell (RBC) Glycoproteins

Abstract
:1. Introduction
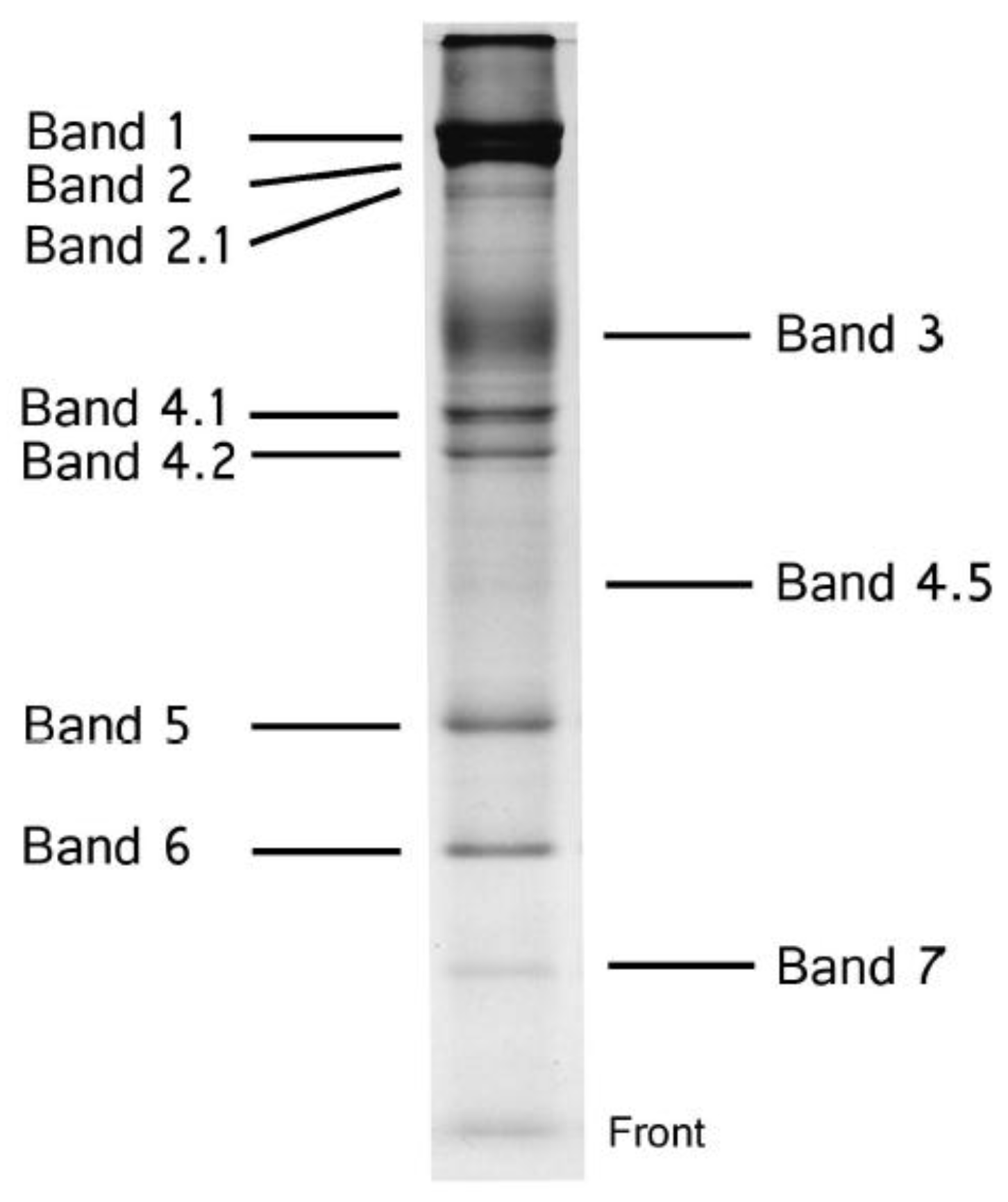
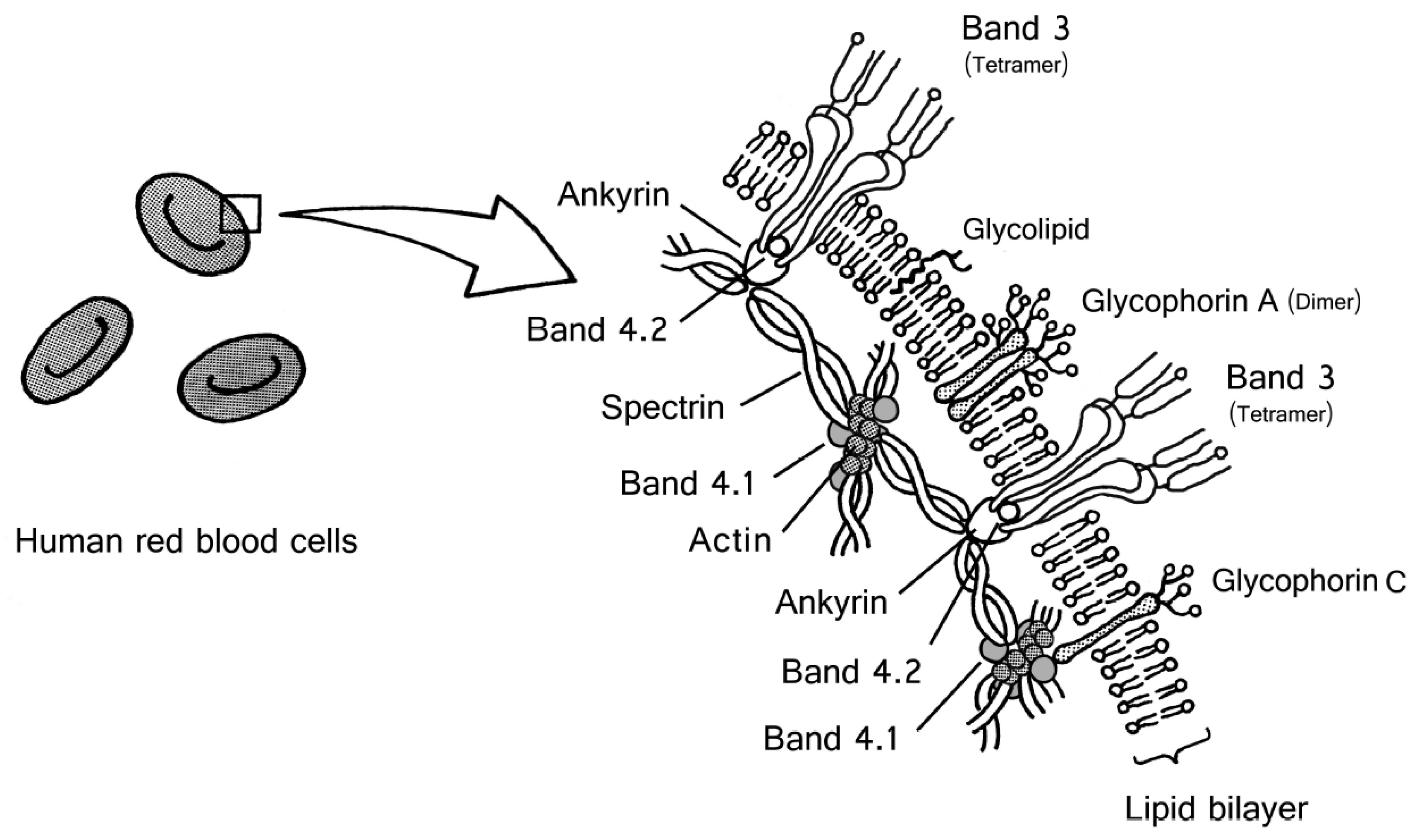
2. Structure of the Human Red Blood Cell Membrane
3. Structure of Band 3
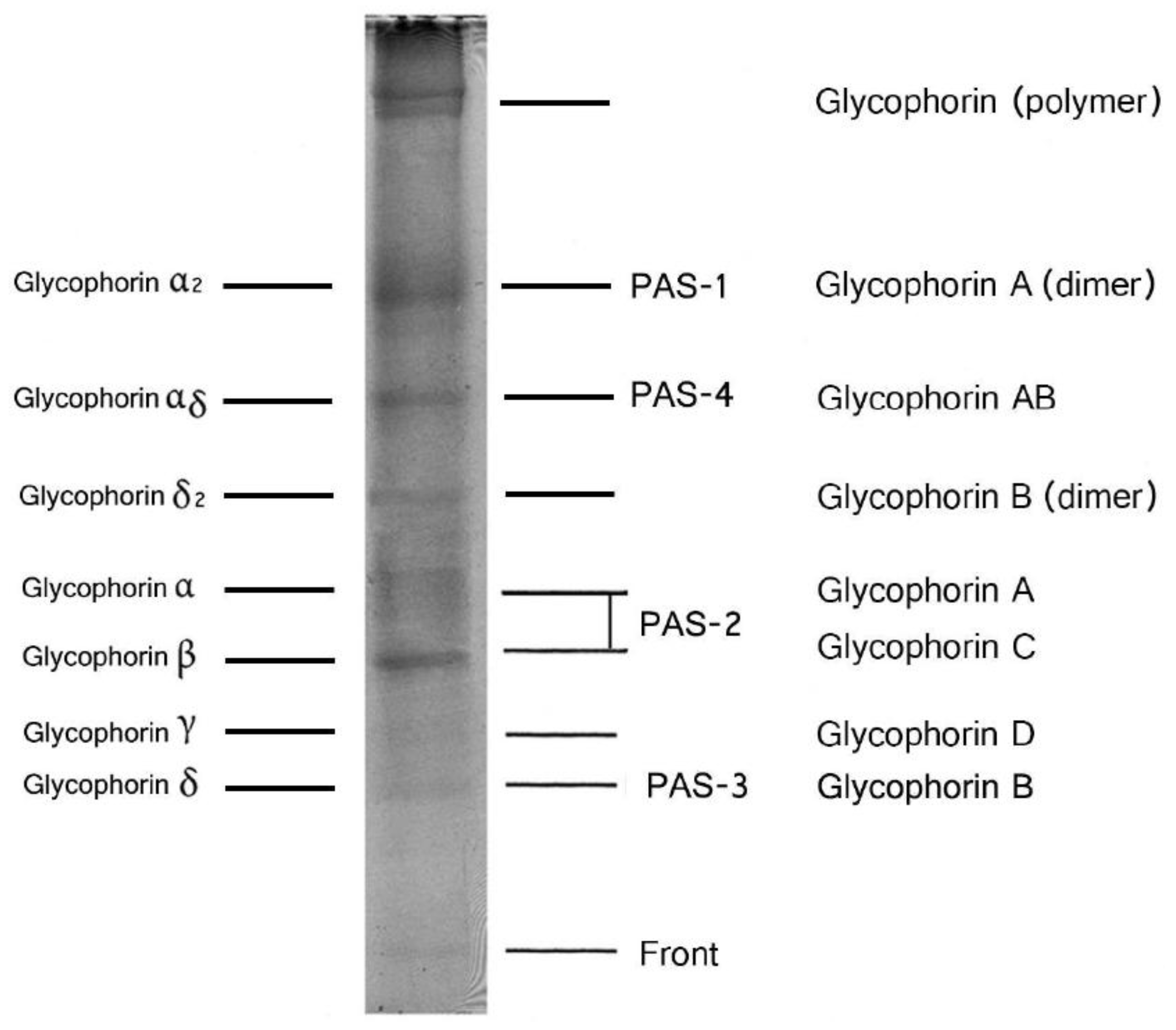
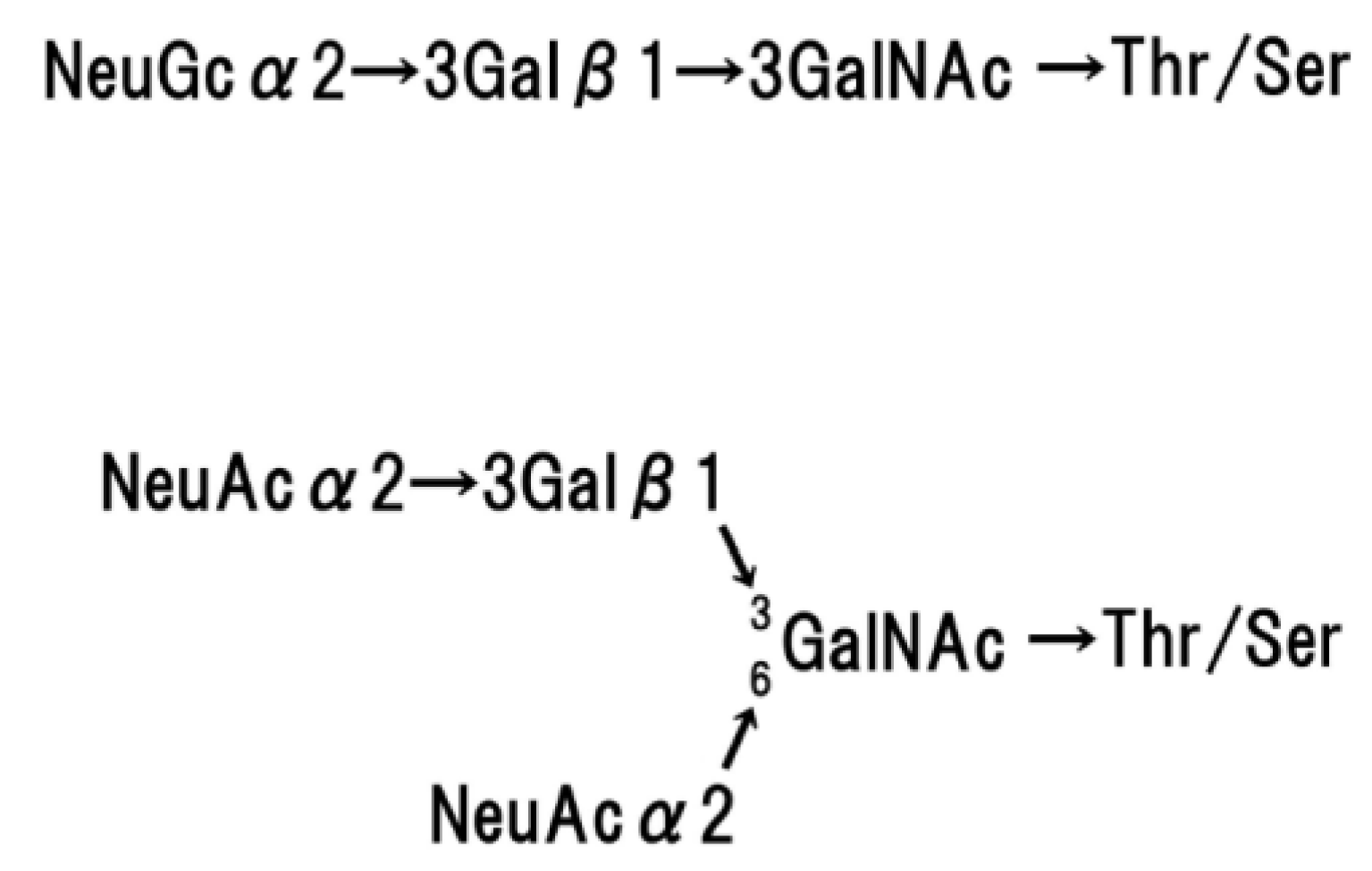
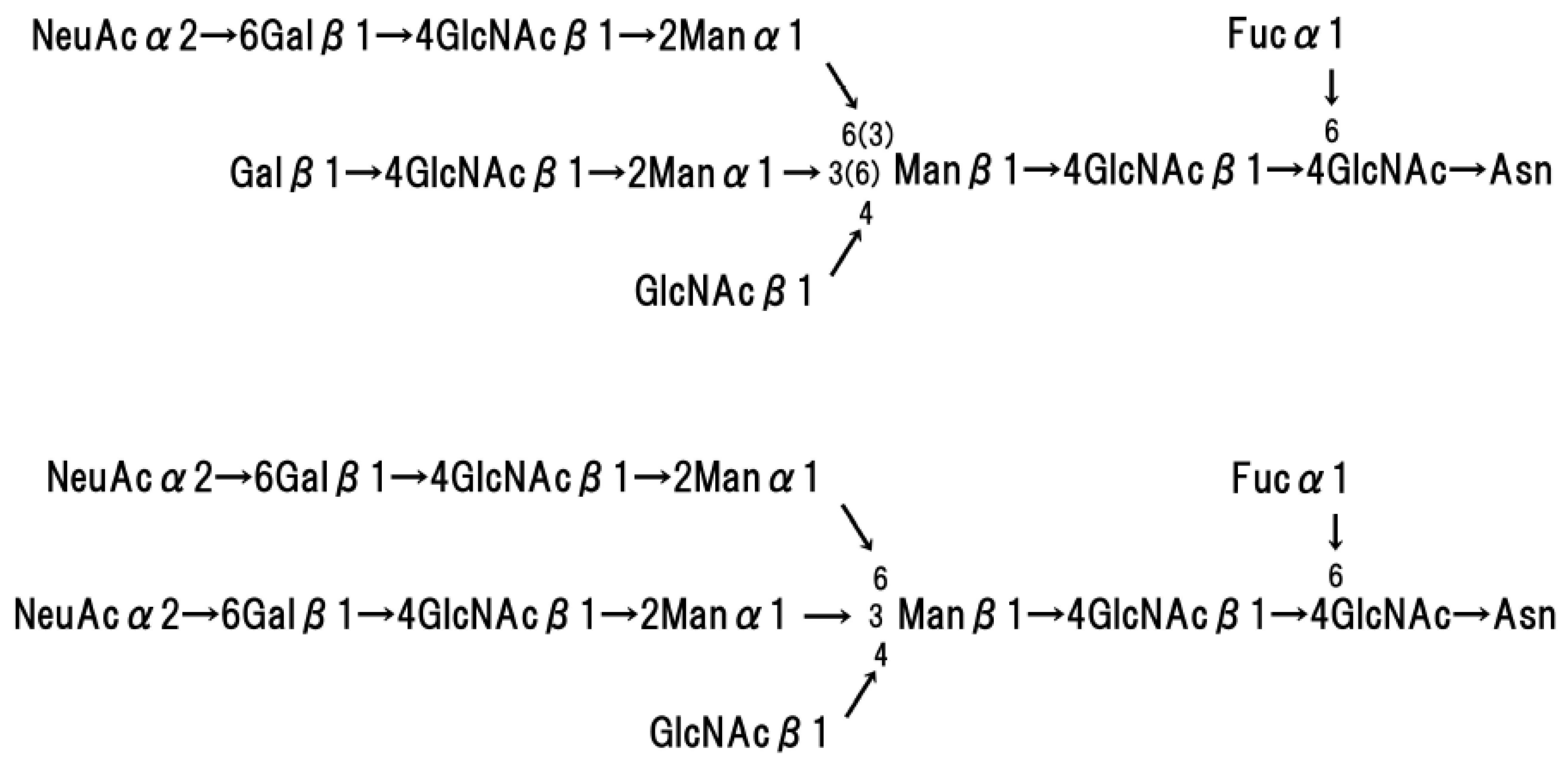
4. Structure of Human Glycophorins
5. Other Glycoprotein in the Human Red Blood Cell Membrane
6. Physiological Function of Band 3
7. Physiological Function of Human Glycophorins
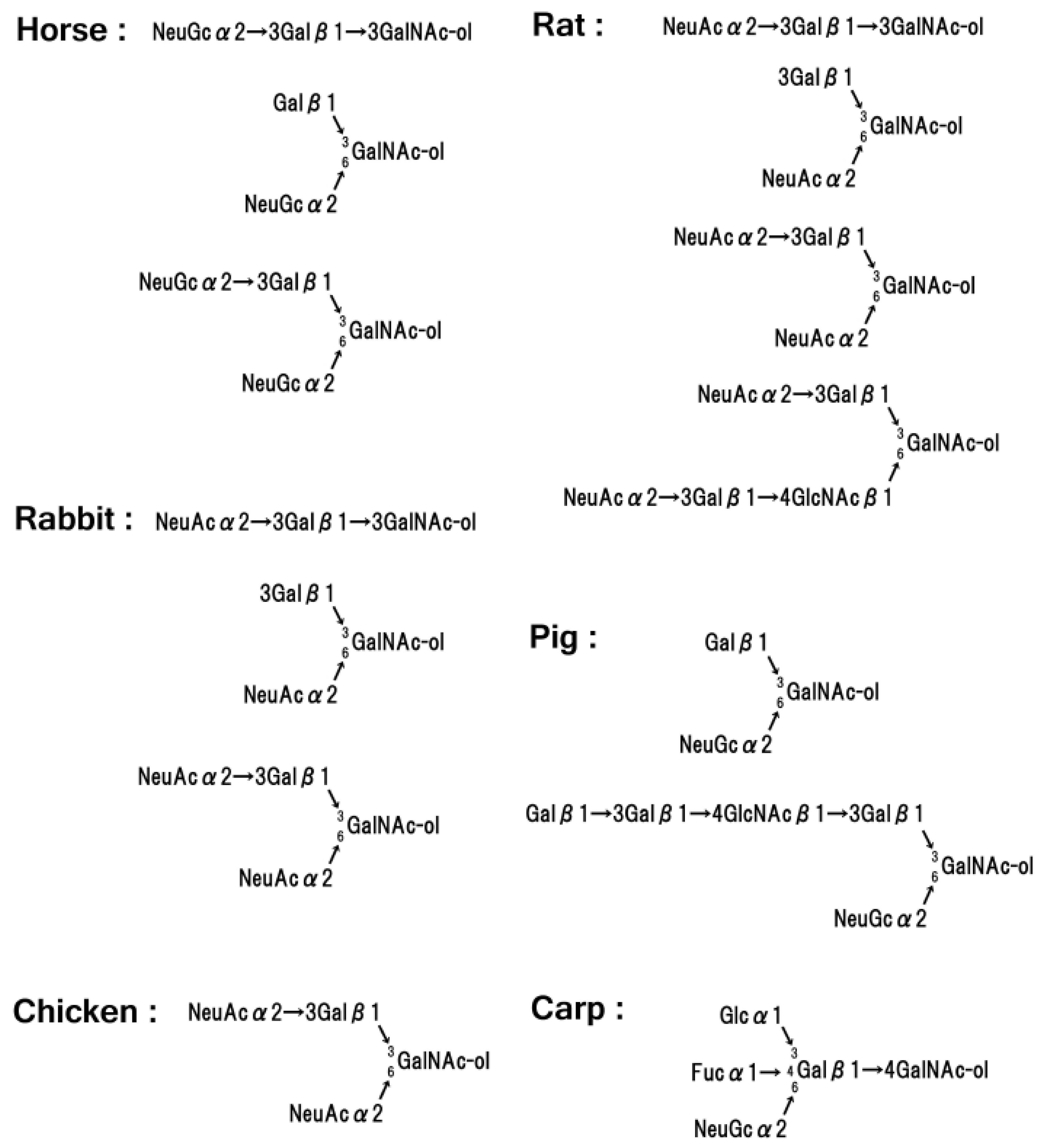
8. Glycoproteins in Red Cell Membranes of Non-Human Origin
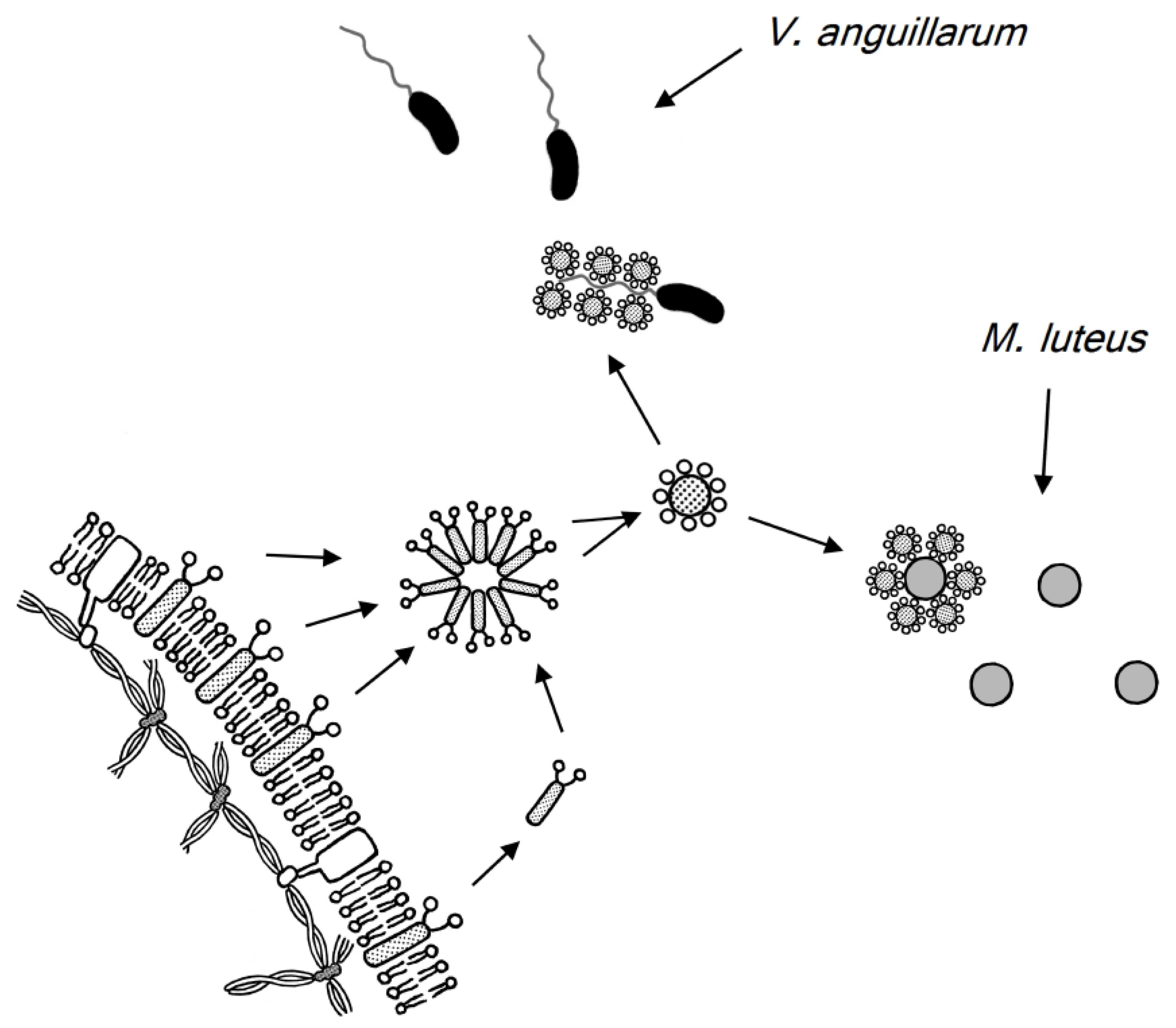
9. Glycophorin in Carp Red Blood Cell Membranes
10. Conclusions
Acknowledgments
Conflicts of Interest
References
- Fairbanks, G.; Steck, T.L.; Wallach, D.F.H. Electrophoretic analysis of the major polypeptides of the human erythrocyte membrane. Biochemistry 1971, 10, 2606–2617. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Tanner, M.J.A. Molecular and cellular biology of the erythrocyte anion exchanger (AE 1). Semin. Hematol. 1993, 30, 34–57. [Google Scholar] [PubMed]
- Steck, T. The organization of proteins in the human red blood cell membrane: A review. J. Cell Biol. 1974, 62, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Doerner, K.C.; White, B.A. Detection of glycoproteins separated by nondenaturing polyacrylamide gel electrophoresis using the periodic acid-Schiff stain. Anal. Biochem. 1990, 187, 147–150. [Google Scholar] [CrossRef]
- Anstee, D.J.; Tanner, M.J.A. Structure and function of the red cell membrane sialoglycoproteins. Br. J. Haematol. 1986, 64, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Anstee, D.J.; Mawby, W.J.; Tanner, M.J.A. Abnormal blood-group-Ss-active sialoglycoproteins in the membrane of Miltenberger class III, IV and V human erythrocytes. Biochem. J. 1979, 183, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Owens, J.W.; Mueller, T.J.; Morrison, M. A minor sialoglycoprotein of the human erythrocyte membrane. Arch. Biochem. Biophys. 1980, 204, 247–254. [Google Scholar] [CrossRef]
- Grefrath, S.P.; Reynolds, J.A. The molecular weight of the major glycoprotein from the human erythrocyte membrane. Proc. Nat. Acad. Sci. USA 1974, 71, 3913–3916. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, M. Molecular-genetics of the glycophorin A gene cluster. Semin. Hematol. 1993, 30, 138–151. [Google Scholar] [PubMed]
- Blanchard, D.; Dahr, W.; Hummel, M.; Lartron, F.; Beyreuther, K.; Cartron, J.-P. Glycophorins B and C from human erythrocyte membranes. Purification and sequence analysis. J. Biol. Chem. 1987, 262, 5808–5811. [Google Scholar] [PubMed]
- Mohandas, N.; Chasis, J.A. Red blood cell deformability, membrane material properties and shape: Regulation by transmembrane, skeletal and cytosolic proteins and lipids. Semin. Hematol. 1993, 30, 171–192. [Google Scholar] [PubMed]
- Bennet, V. The membrane skeleton of human erythrocytes and its implications for more complex cells. Ann. Rev. Biochem. 1985, 54, 273–304. [Google Scholar] [CrossRef] [PubMed]
- Lux, S.E., IV. Anatomy of the red cell membrane skeleton: Unanswered questions. Blood 2016, 127, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Cordat, E.; Reithmeier, R.A.F. Structure, function, and trafficking of SLC4 and SLC26 anion transporters. Curr. Top. Membr. 2014, 73, 3–15. [Google Scholar]
- Hemming, N.J.; Anstee, D.J.; Mawby, W.J.; Reid, M.E.; Tanner, M.J.A. Localization of the protein 4.1-binding site on human erythrocyte glycophorins C and D. Biochem. J. 1994, 299, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Hemming, N.J.; Anstee, D.J.; Staricoff, M.A.; Tanner, M.J.A.; Mohandas, N. Identification of the membrane attachment sites for protein 4.1 in the human erythrocyte. J. Biol. Chem. 1995, 270, 5360–5366. [Google Scholar] [CrossRef] [PubMed]
- Reid, M.E.; Takakuwa, Y.; Conboy, J.; Tchernia, G.; Mohandas, N. Glycophorin C content of human erythrocyte membrane is regulated by protein 4.1. Blood 1990, 11, 2229–2234. [Google Scholar]
- Alloisio, N.; Dalla Venezia, N.; Rana, A.; Andrabi, K.; Texier, P.; Gilsanz, F.; Cartron, J.-P.; Delaunay, J.; Chishti, A.H. Evidence that red blood cell protein p55 may participate in the skeleton-membrane linkage that involves protein 4.1 and glycophorin C. Blood 1993, 82, 1323–1327. [Google Scholar] [PubMed]
- Staricoff, M.A.; Tanner, M.J.A. Role of band 3 and glycophorin C in the maintenance of the shape and mechanical properties of the human red blood cell. Cell. Mol. Biol. Lett. 1996, 1, 151–161. [Google Scholar]
- Lovrien, R.E.; Anderson, R.A. Glycophorin is linked by band 4.1 protein to the human erythrocyte membrane skeleton. Nature 1984, 307, 655–658. [Google Scholar]
- Mohandas, N.; Gallagher, P.G. Red cell membrane: Past, present, and future. Blood 2008, 112, 3939–3948. [Google Scholar] [CrossRef] [PubMed]
- Okubo, K.; Hamasaki, N. Recent progress on band 3 protein. Seikagaku 1992, 64, 1116–1120. (In Japanese) [Google Scholar] [PubMed]
- Steck, T.L.; Koziarz, J.J.; Singh, M.K.; Reddy, G.; Köhler, H. Preparation and analysis of seven major, topographically defined fragments of band 3, the predominant transmembrane polypeptide of human erythrocyte membranes. Biochemistry 1978, 17, 1216–1222. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, T.; Kobayashi-Yurugi, T.; Alguel, Y.; Iwanari, H.; Hatae, H.; Iwata, M.; Abe, Y.; Hino, T.; Ikeda-Suno, C.; Kuma, H.; et al. Crystal structure of the anion exchanger domain of human erythrocyte band 3. Science 2015, 350, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, T.; Irimura, T.; Osawa, T. The carbohydrate moiety of band 3 glycoprotein of human erythrocyte membranes. Structures of lower molecular weight oligosaccharides. J. Biol. Chem. 1981, 256, 10497–10502. [Google Scholar] [PubMed]
- Fukuda, M.; Dell, A.; Oates, J.E.; Fukuda, M.N. Structure of branched lactosaminoglycan, the carbohydrate moiety of band 3 isolated from adult human erythrocytes. J. Biol. Chem. 1984, 259, 8260–8273. [Google Scholar] [PubMed]
- Chasis, J.A.; Mohandas, N. Red blood cell glycophorins. Blood 1992, 80, 1869–1879. [Google Scholar] [PubMed]
- Cartron, J.-P.; Colin, Y.; Kudo, S.; Fukuda, M. Molecular Genetics of Human Erythrocyte Sialoglycoproteins Glycophorins A, B, C, and D; Springer: Boston, MA, USA, 1990; pp. 299–335. [Google Scholar]
- Blanchard, D. Human red cell glycophorins: Biochemical and antigenic properties. Transfus. Med. Rev. 1990, 4, 170–186. [Google Scholar] [CrossRef]
- Furthmayer, H. Glycophorins A, B, and C: A family of sialoglycoproteins. Isolation and preliminary characterization of trypsin derived peptides. J. Supramol. Struct. 1978, 9, 79–95. [Google Scholar] [CrossRef] [PubMed]
- De Isla, N.G.; Riquelme, B.D.; Rasia, R.J.; Valverde, J.R.; Stoltz, J.F. Quantification of glycophorin A and glycophorin B on normal human RBCs by flow cytometry. Transfusion 2003, 43, 1145–1152. [Google Scholar] [CrossRef] [PubMed]
- Tomita, M.; Marchesi, V.T. Amino-acid sequence and oligosaccharide attachment sites of human erythrocyte glycophorin. Proc. Natl. Acad. Sci. USA 1975, 72, 2964–2968. [Google Scholar] [CrossRef] [PubMed]
- Tomita, M.; Furthmayr, H.; Marchesi, V.T. Primary structure of human erythrocyte glycophorin A. Isolation and characterization of peptides and complete amino acid sequence. Biochemistry 1978, 17, 4756–4770. [Google Scholar] [CrossRef] [PubMed]
- Bormann, B.-J.; Knowles, W.J.; Marchesi, V.T. Synthetic peptides mimic the assembly of transmembrane glycoproteins. J. Biol. Chem. 1989, 264, 4033–4037. [Google Scholar] [PubMed]
- Lemmon, M.A.; Flanagan, J.M.; Hunt, J.F.; Adair, B.D.; Bormann, B.J.; Dempsey, C.E.; Engelman, D.M. Glycophorin A dimerization is driven by specific interactions between transmembrane α-helices. J. Biol. Chem. 1992, 267, 7683–7689. [Google Scholar] [PubMed]
- Brosig, B.; Langosch, D. The dimerization motif of the glycophorin A transmembrane segment in membranes: Importance of glycine residues. Protein Sci. 1998, 7, 1052–1056. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, K.R.; Prestegard, J.H.; Engelman, D.M. A transmembrane helix dimer: Structure and implications. Science 1997, 276, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.O.; Song, D.; Shekar, S.; Groesbeek, M.; Ziliox, M.; Aimoto, S. Structure of the transmembrane dimer interface of glycophorin A in membrane bilayers. Biochemistry 2001, 40, 6553–6558. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.O.; Eilers, M.; Song, D.; Crocker, E.; Ying, W.; Groesbeek, M.; Metz, G.; Ziliox, M.; Aimoto, S. Implications of threonine hydrogen bonding in the glycophorin A transmembrane helix dimer. Biophys. J. 2002, 82, 2476–2486. [Google Scholar] [CrossRef]
- Anbazhagan, V.; Schneider, D. The membrane environment modulates self-association of the human GpA TM domain—Implications for membrane protein folding and transmembrane signaling. Biochim. Biophys. Acta 2010, 1798, 1899–1907. [Google Scholar] [CrossRef] [PubMed]
- Shan, X.; Davis, J.H.; Chu, J.W.K.; Sharom, F.J. 2H-NMR investigation of DMPC/glycophorin bilayers. Biochim. Biophys. Acta 1994, 1193, 127–137. [Google Scholar] [CrossRef]
- Thomas, D.B.; Winzler, R.J. Structural studies on human erythrocyte glycoproteins. Alkali-labile oligosaccharides. J. Biol. Chem. 1969, 244, 5943–5946. [Google Scholar] [PubMed]
- Fukuda, M.; Lauffenburger, M.; Sasaki, H.; Rogers, M.E.; Dell, A. Structures of novel sialylated O-linked oligosaccharides isolated from human erythrocyte glycophorins. J. Biol. Chem. 1987, 262, 11952–11957. [Google Scholar] [PubMed]
- Pisano, A.; Redmond, J.W.; Williams, K.L.; Gooley, A.A. Glycosylation sites identified by solid-phase Edman degradation: O-linked glycosylation motifs on human glycophorin A. Glycobiology 1993, 3, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Yoshima, H.; Furthmayr, H.; Kobata, A. Structures of the asparagine-linked sugar chains of glycophorin A. J. Biol. Chem. 1980, 255, 9713–9718. [Google Scholar] [PubMed]
- Marchesi, V.T.; Furthmayr, H.; Tomita, M. The red cell membrane. Ann. Rev. Biochem. 1976, 45, 667–698. [Google Scholar] [CrossRef] [PubMed]
- Furthmayr, H.; Tomita, M.; Marchesi, V.T. Fractionation of the major sialoglycopeptides of the human red blood cell membrane. Biochem. Biophys. Res. Commun. 1975, 65, 113–121. [Google Scholar] [CrossRef]
- Marchesi, V.T.; Andrews, E.P. The use of lithium diiodosalicylate (LIS) to isolate glycoproteins from cell membranes. Science 1971, 174, 1247–1248. [Google Scholar] [CrossRef] [PubMed]
- Furthmayr, H. Structural comparison of glycophorins and immunochemical analysis of genetic variants. Nature 1978, 271, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Siebert, P.D.; Fukuda, M. Isolation and characterization of human glycophorin A cDNA clones by a synthetic oligonucleotide approach: Nucleotide sequence and mRNA structure. Proc. Natl. Acad. Sci. USA 1986, 83, 1665–1669. [Google Scholar] [CrossRef] [PubMed]
- Siebert, P.D.; Fukuda, M. Molecular cloning of a human glycophorin B cDNA: Nucleotide sequence and genomic relationship to glycophorin A. Proc. Natl. Acad. Sci. USA 1987, 84, 6735–6739. [Google Scholar] [CrossRef] [PubMed]
- Tate, C.G.; Tanner, M.J.A. Isolation of cDNA clones for human erythrocyte membrane sialoglycoproteins α and δ. Biochem. J. 1988, 254, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Mattei, M.G.; Colin, Y.; Le Van Kim, C.; Mattei, J.F.; Cartron, J.-P. Localization of the gene for human erythrocyte glycophorin C to chromosome 2, q14–q21. Hum. Genet. 1986, 74, 420–422. [Google Scholar] [CrossRef] [PubMed]
- Le Van Kim, C.; Colin, Y.; Blanchard, D.; Dahr, W.; London, J.; Cartron, J.-P. Gerbich blood group deficiency of the Ge:-1,-2,-3 and Ge:-1,-2, 3 types. Immunochemical study and genomic analysis with cDNA probes. Eur. J. Biochem. 1987, 165, 571–579. [Google Scholar] [PubMed]
- Cartron, J.-P.; Le Van Kim, C.; Colin, Y. Glycophorin C and related glycophorins: Structure, function, and regulation. Semin. Hematol. 1993, 30, 152–168. [Google Scholar] [PubMed]
- Dahr, W.; Beyreuther, K.; Kordowicz, M.; Krüger, J. N-terminal amino acid sequence of sialoglycoprotein D (glycophorin C) from human erythrocyte membranes. Eur. J. Biochem. 1982, 125, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Kudo, S.; Fukuda, M. Identification of a novel human glycophorin, glycophorin E, by isolation of genomic clones and complementary DNA clones utilizing polymerase chain reaction. J. Biol. Chem. 1990, 265, 1102–1110. [Google Scholar] [PubMed]
- Vignal, A.; Rahuel, C.; London, J.; Zahar, B.C.; Schaff, S.; Hattab, C.; Okubo, Y.; Cartron, J.-P. A novel gene member of the human glycophorin A and B gene family. Eur. J. Biochem. 1990, 191, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Kudo, S.; Fukuda, M. Contribution of gene conversion to the retention of the sequence for M blood group type determinant in glycophorin E gene. J. Biol. Chem. 1994, 269, 22969–22974. [Google Scholar] [PubMed]
- Onetti, R.; Baulida, J.; Bassols, A. Increased glucose transport in ras-transformed fibroblasts: A possible role for N-glycosylation of GLUT1. FEBS Lett. 1997, 407, 267–270. [Google Scholar] [CrossRef]
- Counillon, L.; Pouysségur, J.; Reithmeier, R.A.F. The Na+/H+ exchanger NHF-1 possesses N- and O-linked glycosylation restricted the first N-terminal extracellular domain. Biochemistry 1994, 33, 10463–10469. [Google Scholar] [CrossRef] [PubMed]
- Tse, C.-M.; Levine, S.A.; Yun, C.H.C.; Khurana, S.; Donowitz, M. The plasma membrane Na+/H+ exchanger 2 is an O-linked but not an N-linked sialoglycoprotein: Use of a polyclonal antibody to identify and characterize glycosylation. Biochemistry 1994, 33, 12954–12961. [Google Scholar] [CrossRef] [PubMed]
- Orlowski, J.; Grinstein, S. Na+/H+ exchangers of mammalian cells. J. Biol. Chem. 1997, 272, 22373–22376. [Google Scholar] [CrossRef] [PubMed]
- Denker, S.P.; Huang, D.C.; Orlowski, J.; Furthmayr, H.; Barber, D.L. Direct binding of the Na–H exchanger NHE1 to ERM proteins regulates the cortical cytoskeleton and cell shape independently of H+ translocation. Mol. Cell 2000, 6, 1425–1436. [Google Scholar] [CrossRef]
- Ponta, H.; Wainwright, D.; Herrlich, P. Molecules in focus The CD44 protein family. Int. J. Biochem. Cell Biol. 1998, 30, 299–305. [Google Scholar] [CrossRef]
- Skelton, T.P.; Zeng, C.; Nocks, A.; Stamenkovic, I. Glycosylation provides both stimulatory and inhibitory effects on cell surface and soluble CD44 binding to hyaluronan. J. Cell Biol. 1998, 140, 431–446. [Google Scholar] [CrossRef] [PubMed]
- Isacke, C.M.; Yarwood, H. The hyaluronan receptor, CD44. Int. J. Biochem. Cell Biol. 2002, 34, 718–721. [Google Scholar] [CrossRef]
- Mawby, W.J.; Holmes, C.H.; Anstee, D.J.; Spring, F.A.; Tanner, M.J.A. Isolation and characterization of CD47 glycoprotein: A multispanning membrane protein which is the same as integrin-associated protein (IAP) and the ovarian tumour marker OA3. Biochem. J. 1994, 304, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, S.; Boder, E.T.; Discher, D.E. Phylogenetic divergence of CD47 interactions with human signal regulatory protein α reveals locus of species specificity. Implications for the binding site. J. Biol. Chem. 2007, 282, 1805–1818. [Google Scholar] [CrossRef] [PubMed]
- Anstee, D.J. Blood group-active surface molecules of the human red blood cell. Vox Sang. 1990, 58, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Grodecka, M.; Bertrand, O.; Karolak, E.; Lisowski, M.; Waśniowska, K. One-step immunopurification and lectinochemical characterization of the Duffy atypical chemokine receptor from human erythrocytes. Glycoconj. J. 2012, 29, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Ryan, J.R.; Stoute, J.A.; Amon, J.; Dunton, R.F.; Mtalib, R.; Koros, J.; Owour, B.; Luckhart, S.; Wirtz, R.A.; Barnwell, J.W.; et al. Evidence for transmission of Plasmodium vivax among a Duffy antigen negative population in western Kenya. Am. J. Trop. Med. Hyg. 2006, 75, 575–581. [Google Scholar] [PubMed]
- Daniels, G.; Khalid, G. Identification, by immunoblotting, of the structures carrying Lutheran and para-Lutheran blood group antigens. Vox Sang. 1989, 57, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Parson, S.F.; Mallinson, G.; Judson, P.A.; Anstee, D.J.; Tanner, M.J.; Daniels, G.L. Evidence that the Lub blood group antigen is located on red cell membrane glycoproteins of 85 and 78 kd. Transfussion 1987, 27, 61–63. [Google Scholar] [CrossRef]
- Redman, C.M.; Avellino, G.; Pfeffer, S.R.; Mukherjee, T.K.; Nichols, M.; Rubinstein, P.; Marsh, W.L. Kell blood group antigens are part of a 93,000-dalton red cell membrane protein. J. Biol. Chem. 1986, 261, 9521–9525. [Google Scholar] [PubMed]
- Jaber, A.; Blanchard, D.; Goossens, D.; Bloy, C.; Lambin, P.; Rouger, P.; Salmon, C.; Cartron, J.-P. Characterization of the blood group Kell (K1) antigen with a human monoclonal antibody. Blood 1989, 73, 1597–1602. [Google Scholar] [PubMed]
- Lee, S.; Zambas, E.D.; Marsh, W.L.; Redman, C.M. Molecular cloning and primary structure of Kell blood group protein. Proc. Natl. Acad. Sci. USA 1991, 88, 6353–6357. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Russo, D.; Redman, C. Functional and structural aspects of the Kell blood group system. Transfus. Med. Rev. 2000, 14, 93–103. [Google Scholar] [CrossRef]
- Yu, L.-C.; Twu, Y.-C.; Chou, M.-L.; Reid, M.E.; Gray, A.R.; Moulds, J.M.; Chang, C.-Y.; Lin, M. The molecular genetics of the human I locus and molecular background explain the partial association of the adult i phenotype with congenital cataracts. Blood 2003, 101, 2081–2088. [Google Scholar] [CrossRef] [PubMed]
- Huflejt, M.E.; Vuskovic, M.; Vasiliu, D.; Xu, H.; Obukhova, P.; Shilova, N.; Tuzikov, A.; Galanina, O.; Arun, B.; Lu, K.; et al. Anti-carbohydrate antibodies of normal sera: Findings, surprises and challenges. Mol. Immunol. 2009, 46, 3037–3049. [Google Scholar] [CrossRef] [PubMed]
- Beppu, M.; Kikugawa, K. Senescent cell antigens in the clearance of senescent cells. Seikagaku 1995, 67, 303–307. (In Japanese) [Google Scholar] [PubMed]
- Reithmeier, R.A.F.; Casey, J.R.; Kalli, A.C.; Sansom, M.S.P.; Alguel, Y.; Iwata, S. Band 3, the human red cell chloride/bicarbonate anion exchanger (AE1, SLC4A1), in a structural context. Biochim. Biophys. Acta 2016, 1858, 1507–1532. [Google Scholar] [CrossRef] [PubMed]
- Alper, S.L.; Kopito, R.R.; Libresco, S.M.; Lodish, H.F. Cloning and characterization of a murine band 3-related cDNA from kidney and from a lymphoid cell line. J. Biol. Chem. 1988, 263, 17092–17099. [Google Scholar] [PubMed]
- Kopito, R.R.; Lee, B.S.; Simmons, D.M.; Lindsey, A.E.; Morgans, C.W.; Schneider, K. Regulation of intracellular pH by a neuronal homolog of the erythrocyte anion exchanger. Cell 1989, 59, 927–937. [Google Scholar] [CrossRef]
- Casey, J.R.; Pirraglia, C.A.; Reithmeier, R.A.F. Enzymatic deglycosylation of human Band 3, the anion transport protein of the erythrocyte membrane. Effect on protein structure and transport properties. J. Biol. Chem. 1992, 267, 11940–11948. [Google Scholar] [PubMed]
- Kay, M.M.B. Location of senescent cell antigen on band 3. Proc. Natl. Acad. Sci. USA 1984, 81, 5753–5757. [Google Scholar] [CrossRef] [PubMed]
- Kay, M.M.B. Isolation of the phagocytosis-inducing IgG-binding antigen on senescent somatic cells. Nature 1981, 289, 491–494. [Google Scholar] [CrossRef] [PubMed]
- Kay, M.M.B.; Goodman, S.R.; Sorensen, K.; Whitfield, C.L.; Wong, P.; Zaki, L.; Rudloff, V. Senescent cell antigen is immunologically related to band 3. Proc. Natl. Acad. Sci. USA 1983, 80, 1631–1635. [Google Scholar] [CrossRef] [PubMed]
- Beppu, M.; Mizukami, A.; Nagoya, M.; Kikugawa, K. Binding of anti-band 3 autoantibody to oxidatively damaged erythrocytes. Formation of senescent antigen on erythrocyte surface by an oxidative mechanism. J. Biol. Chem. 1990, 265, 3226–3233. [Google Scholar] [PubMed]
- Beppu, M.; Mizukami, A.; Nagoya, M.; Kikugawa, K. Generation of senescent antigen on erythrocytes by partial blocking of SH groups of the membrane proteins. J. Pharmacobio-Dyn. 1992, 15, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Beppu, M.; Mizukami, A.; Ando, K.; Kikugawa, K. Antigenic determinants of senescent antigen of human erythrocytes are located in sialylated carbohydrate chains of Band 3 glycoprotein. J. Biol. Chem. 1992, 267, 14691–14696. [Google Scholar] [PubMed]
- Ando, K.; Kikugawa, K.; Beppu, M. Involvement of sialylated poly-N-acetyllactosaminyl sugar chains of band 3 glycoprotein on senescent erythrocytes in anti-band 3 autoantibody binding. J. Biol. Chem. 1994, 269, 19394–19398. [Google Scholar] [PubMed]
- Kay, M.M.B.; Marchalonis, J.J.; Hughes, J.; Watanabe, K.; Schluter, S.F. Definition of a physiologic aging autoantigen by using synthetic peptides of membrane protein band 3: Localization of the active antigenic sites. Proc. Natl. Acad. Sci. USA 1990, 87, 5734–5738. [Google Scholar] [CrossRef] [PubMed]
- Lutz, H.U.; Gianora, O.; Nater, M.; Schweizer, E.; Stammler, P. Naturally occurring anti-band 3 antibodies bind to protein rather than to carbohydrate on band 3. J. Biol. Chem. 1993, 268, 23562–23566. [Google Scholar] [PubMed]
- Blumenfeld, O.O.; Huang, C.-H. Molecular genetics of the glycophorin gene family, the antigens for MNSs blood groups: Multiple gene rearrangements and modulation of splice site usage result in extensive diversification. Hum. Mutat. 1995, 6, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Hamaguchi, H.; Cleve, H. Solubilization of human erythrocyte membrane glycoproteins and separation of the MN glycoprotein from a glycoprotein with I, S, and A activity. Biochim. Biophys. Acta 1972, 278, 271–280. [Google Scholar] [CrossRef]
- Blumenfeld, O.O.; Zvilichovsky, B. Isolation of glycoproteins from red cell membranes using pyridine. Methods Enzymol. 1972, 28, 245–252. [Google Scholar]
- Schulte, T.H.; Marchesi, V.T. Self-association of human erythrocyte glycophorin A. Appearance of low mobility bands on sodium dodecyl sulfate gels. Biochim. Biophys. Acta 1978, 508, 425–430. [Google Scholar] [CrossRef]
- Furthmayr, H.; Marchesi, V.T. Subunit structure of human erythrocyte glycophorin A. Biochemistry 1976, 15, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Janado, M.; Azuma, J.; Onodera, K. Heterogeneity of a human erythrocyte membrane glycoprotein. J. Biochem. 1973, 74, 881–887. [Google Scholar] [PubMed]
- Lemmon, M.A.; Flanagan, J.M.; Treutlein, H.R.; Zhang, J.; Engelman, D.M. Sequence specificity in the dimerization of transmembrane α-helixes. Biochemistry 1992, 31, 12719–12725. [Google Scholar] [CrossRef] [PubMed]
- Marton, L.S.G.; Garvin, J.E. Subunit structure of the major human erythrocyte glycoprotein: Depolymerization by heating ghosts with sodium dodecyl sulfate. Biochem. Biophys. Res. Commun. 1973, 52, 1457–1462. [Google Scholar] [CrossRef]
- Tuech, J.K.; Morrison, M. Human erythrocyte membrane sialoglycoproteins: A study of interconversion. Biochem. Biophys. Res. Commun. 1974, 59, 352–360. [Google Scholar] [CrossRef]
- Johe, K.K.; Smith, A.J.; Vengelen-Tyler, V.; Blumenfeld, O.O. Amino acid sequence of an α-δ-glycophorin hybrid. A structure reciprocal to Sta δ-α-glycophorin hybrid. J. Biol. Chem. 1989, 264, 17486–17493. [Google Scholar] [PubMed]
- Merry, A.H.; Hodson, C.; Thomson, E.; Mallinson, G.; Anstee, D.J. The use of monoclonal antibodies to quantify the levels of sialoglycoproteins α and δ and variant sialoglycoproteins in human erythrocyte membranes. Biochem. J. 1986, 233, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Ada, G.L.; Stone, J.D. Electrophoretic studies of virus-red cell interaction: Mobility gradient of cells treated with viruses of the influenza group and the receptor-destroying enzyme of V. cholerae. Brit. J. Exptl. Pathol. 1950, 31, 263–274. [Google Scholar]
- Dahr, W.; Beyreuther, K.; Steinbach, H.; Gielen, W.; Krüger, J. Structure of the Ss blood group antigens, II. A methionine/threonine polymorphism within the N-terminal sequence of the Ss glycoprotein. Hoppe-Seyler Z. Physiol. Chem. 1980, 361, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Prohaska, R.; Koerner, T.A.W., Jr.; Armitage, I.M.; Furthmayr, H. Chemical and carbon-13 nuclear magnetic resonance studies of the blood group M and N active sialoglycopeptides from human glycophorin A. J. Biol. Chem. 1981, 256, 5781–5791. [Google Scholar] [PubMed]
- Pang, A.J.; Reithmeier, R.A.F. Interaction of anion exchanger 1 and glycophorin A in human erythroleukaemic K562 cells. Biochem. J. 2009, 421, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Anstee, D.J.; Ridgwell, K.; Tanner, M.J.A.; Daniels, G.L.; Parsons, S.F. Individuals lacking the Gerbich blood-group antigen have alterations in the human erythrocyte membrane sialoglycoproteins β and γ. Biochem. J. 1984, 221, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Dahr, W.; Moulds, J.; Baumeister, G.; Moulds, M.; Kiedrowski, S.; Hummel, M. Altered membrane sialoglycoproteins in human erythrocytes lacking the Gerbich blood group antigens. Biol. Chem. Hoppe-Seyler 1985, 366, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Lisowska, E. Antigenic properties of human glycophorins—An update. In The Molecular Immunology of Complex Carbohydrates—2; Springer: Boston, MA, USA, 2001; pp. 155–169. [Google Scholar]
- Takasaki, S.; Kobata, A. Chemical characterization and distribution of ABO blood group active glycoprotein in human erythrocyte membrane. J. Biol. Chem. 1976, 251, 3610–3615. [Google Scholar] [PubMed]
- Wilczyńska, Z.; Miller-Podraza, H.; Kościelak, J. The contribution of different glycoconjugates to the total ABH blood group activity of human erythrocytes. EFBS Lett. 1980, 112, 277–279. [Google Scholar]
- Podbielska, M.; Fredriksson, S.-Å.; Nilsson, B.; Lisowska, E.; Krotkiewski, H. ABH blood group antigens in O-glycans of human glycophorin A. Arch. Biochem. Biophys. 2004, 429, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Sueyoshi, S.; Tsuji, T.; Osawa, T. Carbohydrate-binding specificities of five lectins that bind to O-glycosyl-linked carbohydrate chains. Quantitative analysis by frontal-affinity chromatography. Carbohydr. Res. 1988, 178, 213–224. [Google Scholar] [CrossRef]
- Krotkiewska, B.; Pasek, M.; Krotkiewski, H. Interaction of glycophorin A with lectins as measured by surface plasmon resonance (SPR). Acta Biochim. Pol. 2002, 49, 481–490. [Google Scholar] [PubMed]
- Anderson, R.; Paquette, S.; Lovrien, R. Lectin-erythrocyte interaction with external transmembrane glycophorin saccharides controlling membrane internal cytoskeleta. J. Agric. Food Chem. 2002, 50, 6599–6604. [Google Scholar] [CrossRef] [PubMed]
- Ellens, H.; Bentz, J.; Mason, D.; Zhang, F.; White, J.M. Fusion of influenza hemagglutinin-expressing fibroblasts with glycophorin-bearing liposomes: Role of hemagglutinin surface density. Biochemistry 1990, 29, 9697–9707. [Google Scholar] [CrossRef] [PubMed]
- Kanaseki, T.; Kawasaki, K.; Murata, M.; Ikeuchi, Y.; Ohnishi, S. Structural features of membrane fusion between influenza virus and liposome as revealed by quick-freezing electron microscopy. J. Cell Biol. 1997, 137, 1041–1056. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Suzuki, T.; Matsumoto, M. Isolation and characterization of receptor sialoglycoprotein for hemagglutinating virus of Japan (Sendai virus) from bovine erythrocyte membrane. J. Biochem. 1983, 93, 1621–1633. [Google Scholar] [CrossRef] [PubMed]
- Wybenga, L.E.; Epand, R.F.; Nir, S.; Chu, J.W.K.; Sharom, F.J.; Flanagan, T.D.; Epand, R.M. Glycophorin as a receptor for Sendai virus. Biochemistry 1996, 35, 9513–9518. [Google Scholar] [CrossRef] [PubMed]
- Templeton, T.J.; Keister, D.B.; Muratova, O.; Procter, J.; Kaslow, D.C. Adherence of erythrocytes during exflagellation of Plasmodium falciparum microgametes is dependent on erythrocyte surface sialic acid and glycophorins. J. Exp. Med. 1998, 187, 1599–1609. [Google Scholar] [CrossRef] [PubMed]
- Pasvol, G. How many pathways for invasion of the red blood cell by the malaria parasite? Trends. Parasitol. 2003, 19, 430–432. [Google Scholar] [CrossRef] [PubMed]
- Lobo, C.-A. Babesia divergens and Plasmodium falciparum use common receptors, glycophorins A and B, to invade the human red blood cell. Infect. Immun. 2005, 73, 649–651. [Google Scholar] [CrossRef] [PubMed]
- Ghislaine Mayer, D.C.; Jiang, L.; Achur, R.N.; Kakizaki, I.; Gowda, D.C.; Miller, L.H. The glycophorin C N-linked glycan is a critical component of the ligand for the Plasmodium falciparum erythrocyte receptor BAEBL. PNAS 2006, 103, 2358–2362. [Google Scholar] [CrossRef] [PubMed]
- Cortajarena, A.L.; Goñi, F.M.; Ostolaza, H. Glycophorin as a receptor for Escherichia coli α-hemolysin in erythrocytes. J. Biol. Chem. 2001, 276, 12513–12519. [Google Scholar] [CrossRef] [PubMed]
- Hirst, G.K. The agglutination of red cells by allantoic fluid of chick embryos infected with influenza virus. Science 1941, 94, 22–23. [Google Scholar] [CrossRef] [PubMed]
- Cochet, S.; Volet, G.; Cartron, J.-P.; Bertrand, O. New procedures for glycophorin A purification with high yield and high purity. J. Chromatogr. B 2001, 750, 109–119. [Google Scholar] [CrossRef]
- Lee, S.; Lu, W. Using elastic light scattering of red blood cells to detect infection of malaria parasite. IEEE Trans. Biomed. Eng. 2012, 59, 150–155. [Google Scholar] [PubMed]
- Giger, K.; Habib, I.; Ritchie, K.; Low, P.S. Diffusion of glycophorin A in human erythrocytes. Biochim. Biophys. Acta 2016, 1858, 2839–2845. [Google Scholar] [CrossRef] [PubMed]
- Yiangou, L.; Montandon, R.; Modrzynska, K.; Rosen, B.; Bushell, W.; Hale, C.; Billker, O.; Rayner, J.C.; Pance, A. A stem cell strategy identifies glycophorin C as a major erythrocyte receptor for the rodent malaria parasite Plasmodium berghei. PLoS ONE 2016, 11, e0158238. [Google Scholar] [CrossRef] [PubMed]
- Hasssoun, H.; Hanada, T.; Lutchman, M.; Sahr, K.E.; Palek, J.; Hanspal, M.; Chishti, A.H. Complete deficiency of glycophorin A in red blood cells from mice with targeted inactivation of the band 3 (AE1) gene. Blood 1998, 91, 2146–2151. [Google Scholar]
- Lesley, J.B.; Groves, J.D.; Okubo, Y.; Thilaganathan, B.; Tanner, J.A. Altered band 3 structure and function in glycophorin A- and B-deficient (MkMk) red blood cells. Blood 1994, 84, 916–922. [Google Scholar]
- Hamaguchi, H.; Cleve, H. Solubilization and comparative analysis of mammalian erythrocyte membrane glycoproteins. Biochem. Biophys. Res. Commun. 1972, 47, 459–464. [Google Scholar] [CrossRef]
- Ballas, S.K. Comparative distribution of glyceraldehyde 3-phosphate dehydrogenase activity in human, guinea-pig, rabbit and mouse erythrocytes. Comp. Biochem. Physiol. B 1987, 87, 837–842. [Google Scholar] [CrossRef]
- Matsuyama, R.; Ueda, T.; Inoue, F. SDS-PAGE analysis of membrane proteins on domestic cat (Felis catus) erythrocytes. Med. Biol. 1999, 138, 79–82. (In Japanese) [Google Scholar]
- Fukuda, K.; Tomita, M.; Hamada, A. Isolation and characterization of alkali-labile oligosaccharide units from horse glycophorin. J. Biochem. 1980, 87, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, K.; Kawashima, I.; Tomita, M.; Hamada, A. Structural studies of the acidic oligosaccharide units from bovine glycophorin. Biochim. Biophys. Acta 1982, 717, 278–288. [Google Scholar] [CrossRef]
- Kawashima, I.; Fukuda, K.; Tomita, M.; Hamada, A. Isolation and characterization of alkali-labile oligosaccharide units from porcine erythrocyte glycophorin. J. Biochem. 1982, 91, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Klimas, N.G.; Caldwell, K.E.; Whitney, P.L.; Fletcher, M.A. Comparison of receptor properties of erythrocyte membrane glycoproteins. Dev. Comp. Immunol. 1981, 6, 765–774. [Google Scholar]
- Yamashita, T.; Murayama, J.; Utsumi, H.; Hamada, A. Structural studies of O-glycosidic oligosaccharide units of dog erythrocyte glycophorin. Biochim. Biophys. Acta 1985, 839, 26–31. [Google Scholar] [CrossRef]
- Angel, A.-S.; Grönberg, G.; Krotkiewski, H.; Lisowska, E.; Nilsson, B. Structural analysis of the N-linked oligosaccharides from murine glycophorin. Arch. Biochem. Biophys. 1991, 291, 76–88. [Google Scholar] [CrossRef]
- Murayama, J.-I.; Utsumi, H.; Hamada, A. Amino acid sequence of monkey erythrocyte glycophorin MK. Its amino acid sequence has a striking homology with that of human glycophorin A. Biochim. Biophys. Acta 1989, 999, 273–280. [Google Scholar] [CrossRef]
- Krotkiewski, H. The structure of glycophorins of animal erythrocytes. Glycoconj. J. 1988, 5, 35–48. [Google Scholar] [CrossRef]
- Dockham, P.A.; Vidaver, G.A. Comparison of human and pigeon erythrocyte membrane proteins by one-and two-dimensional gel electrophoresis. Comp. Biochem. Physiol. 1987, 87B, 171–177. [Google Scholar] [CrossRef]
- Weise, M.J.; Ingram, V.M. Proteins and glycoproteins of membranes from developing chick red cells. J. Biol. Chem. 1976, 251, 6667–6673. [Google Scholar] [PubMed]
- Caldwell, A.B. Proteins of the turkey erythrocyte membrane. Biochemistry 1976, 15, 2711–2718. [Google Scholar] [CrossRef] [PubMed]
- Romano, L.; Passow, H. Characterization of anion transport system in trout red blood cell. Am. J. Physiol. 1984, 246, C330–C338. [Google Scholar] [PubMed]
- Jay, D.G. Characterization of the chicken erythrocyte anion exchange protein. J. Biol. Chem. 1983, 258, 9431–9436. [Google Scholar] [PubMed]
- Michel, F.; Rudloff, V. Isolation and characterization of the rainbow trout erythrocyte band-3 protein. FEBS J. 1989, 181, 181–187. [Google Scholar] [CrossRef]
- Hübner, S.; Michel, F.; Rudloff, V.; Appelhans, H. Amino acid sequence of band-3 protein from rainbow trout erythrocytes derived from cDNA. Biochem. J. 1992, 285, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.C. The exterior surface of the chicken erythrocyte. J. Biol. Chem. 1975, 250, 617–622. [Google Scholar] [PubMed]
- Duk, M.; Krotkiewski, H.; Stasyk, T.V.; Lutsik-Kordovsky, M.; Syper, D.; Lisowska, E. Isolation and characterization of glycophorin from nucleated (chicken) erythrocytes. Arch. Biochem. Biophys. 2000, 375, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Aoki, T.; Fukai, M.; Ueno, R. Glycoproteins in red cell membranes from carp and rainbow trout. Fish. Sci. 1996, 62, 498–499. [Google Scholar] [CrossRef]
- Aoki, T.; Chimura, K.; Nakao, N.; Mizuno, Y. Isolation and characterization of glycophorin from carp red blood cell membranes. Membranes 2014, 4, 491–508. [Google Scholar] [CrossRef] [PubMed]
- Aoki, T.; Inoue, T. Glycophorin in red blood cell membranes of healthy and diseased carp, Cyprinus carpio L. J. Fish Dis. 2011, 34, 573–576. [Google Scholar] [CrossRef] [PubMed]
- Aoki, T.; Chimura, K.; Sugiura, H.; Mizuno, Y. Structure of a sialo-oligosaccharide from glycophorin in carp red blood cell membranes. Membranes 2014, 4, 764–777. [Google Scholar] [CrossRef] [PubMed]
- Araki, T.; Aoki, T.; Kitamikado, M. Isolation and identification of a β-1,3-xylanase-producing bacterium. Nippon Suisan Gakkaishi 1987, 53, 2077–2081. [Google Scholar] [CrossRef]
- Uchida, Y. Application of Chitin, Chitosan; Chitin, Chitosan research association, Ed.; Gihodo Shuppan Co., Ltd.: Tokyo, Japan, 1990; pp. 71–98. (In Japanese) [Google Scholar]
- Wilson, M.; Bengtén, E.; Miller, N.W.; Clem, L.W.; Du Pasquier, L.; Warr, G.W. A novel chimeric Ig heavy chain from a teleost fish shares similarities to IgD. Proc. Natl. Acad. Sci. USA 1997, 94, 4593–4597. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Band | Molecular Mass (kDa) | Designation | Function |
|---|---|---|---|
| 1 | 240 | spectrin (α chain) | components of cytoskeleton |
| 2 | 220 | spectrin (β chain) | |
| 2.1 | 210 | ankylin | |
| 3 | 100 | band 3 (AE1) | anion transporter |
| 4.1 | 82 | band 4.1 | components of cytoskeleton |
| 4.2 | 76 | band 4.2 | |
| 4.5 | 55 | band 4.5 (GLUT1) | glucose transporter |
| 5 | 43 | actin | components of cytoskeleton |
| 6 | 35 | glyceraldehyde-3-phoshate dehydrogenase (GAPDH) | glycolytic enzyme |
| 7 | 31 | stomatin | – |
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aoki, T. A Comprehensive Review of Our Current Understanding of Red Blood Cell (RBC) Glycoproteins. Membranes 2017, 7, 56. https://doi.org/10.3390/membranes7040056
Aoki T. A Comprehensive Review of Our Current Understanding of Red Blood Cell (RBC) Glycoproteins. Membranes. 2017; 7(4):56. https://doi.org/10.3390/membranes7040056
Chicago/Turabian StyleAoki, Takahiko. 2017. "A Comprehensive Review of Our Current Understanding of Red Blood Cell (RBC) Glycoproteins" Membranes 7, no. 4: 56. https://doi.org/10.3390/membranes7040056
APA StyleAoki, T. (2017). A Comprehensive Review of Our Current Understanding of Red Blood Cell (RBC) Glycoproteins. Membranes, 7(4), 56. https://doi.org/10.3390/membranes7040056