
Molecular Pathogenesis of Psoriasis and Biomarkers Reflecting Disease Activity

Abstract
:1. Introduction
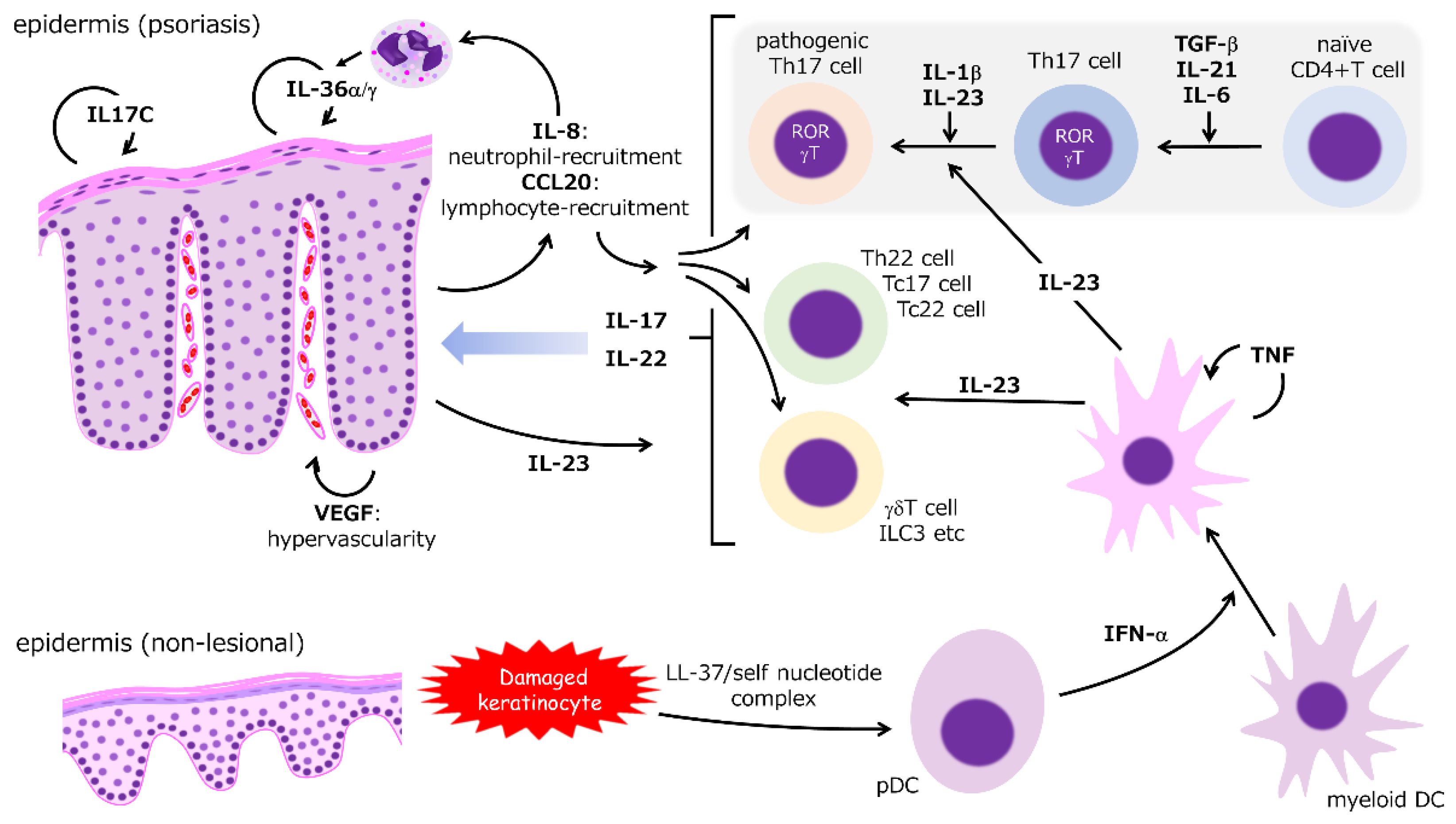
2. Molecular Pathogenesis of Psoriasis
2.1. IL-23
2.2. IL-17
2.3. CCL20/CCR6 Axis
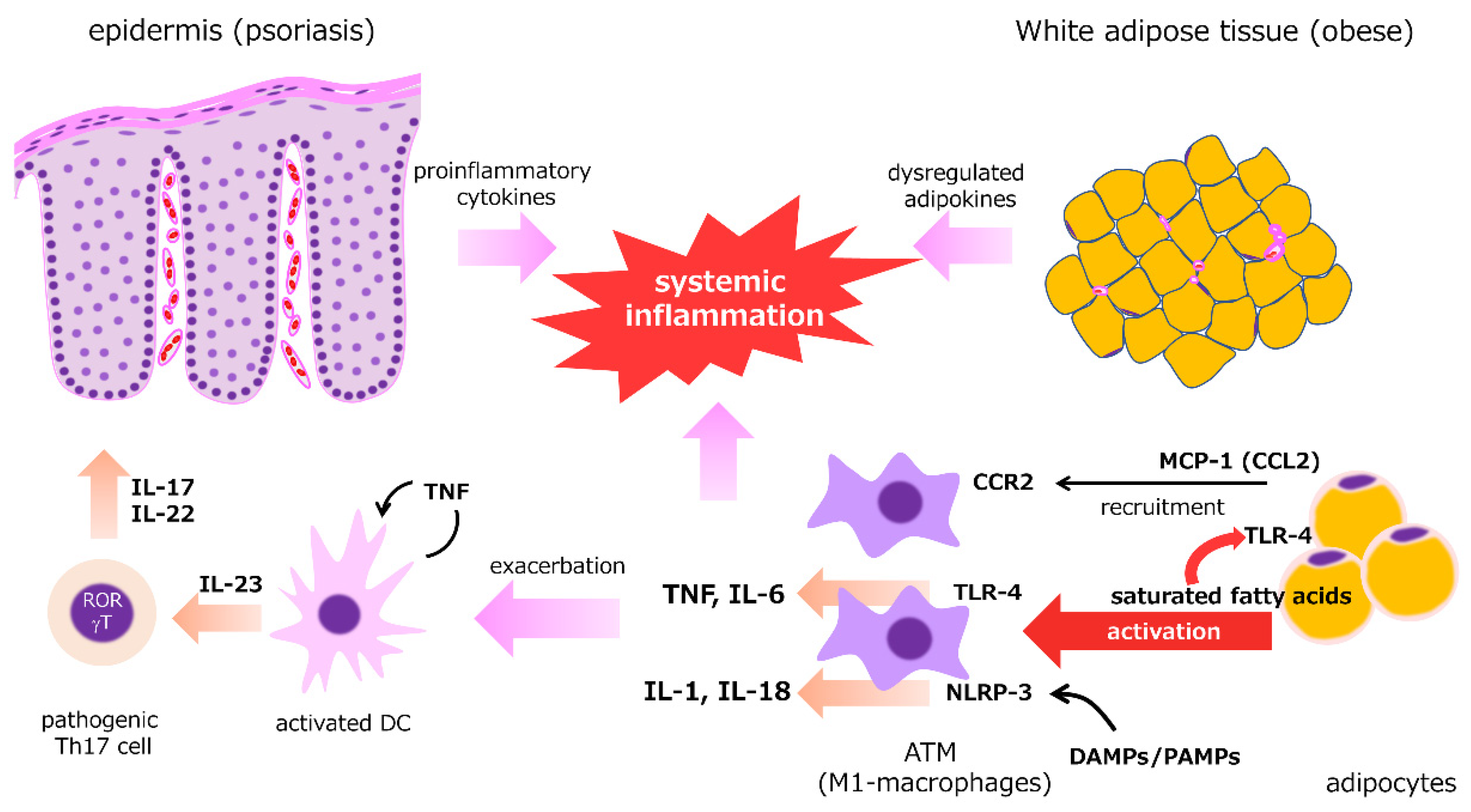
2.4. Adipose Tissue
3. Biomarkers in Psoriasis Treatment
3.1. Peripheral Blood Cell Counts
Neutrophil-to-Lymphocyte Ratio and Platelet-to-Lymphocyte Ratio
3.2. Cytokines and Chemokines
3.2.1. IL-17
3.2.2. IL-22
3.2.3. IL-36
3.2.4. Fractalkine
3.2.5. Thymus and Activation-Regulated Chemokine
3.3. Adipokines
3.3.1. Resistin
3.3.2. Adiponectin
3.4. Antimicrobial Peptides
3.4.1. Defensin 2
3.4.2. S100A
3.5. Protease Inhibitors
3.5.1. Squamous Cell Carcinoma Antigen
3.5.2. Elafin
3.6. Glycoproteins
3.6.1. Leucin-Rich Alpha-2-Glycoprotein
3.6.2. YKL-40
3.7. Fatty Acid-Binding Protein
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nestle, F.O.; Kaplan, D.H.; Barker, J. Psoriasis. N. Engl. J. Med. 2009, 361, 496–509. [Google Scholar] [CrossRef]
- Boehncke, W.H.; Schön, M.P. Psoriasis. Lancet 2015, 386, 983–994. [Google Scholar] [CrossRef]
- Takahashi, H.; Iinuma, S.; Tsuji, H.; Honma, M.; Iizuka, H. Biologics are more potent than other treatment modalities for improvement of quality of life in psoriasis patients. J. Dermatol. 2014, 41, 686–689. [Google Scholar] [CrossRef] [PubMed]
- Imafuku, S.; Kanai, Y.; Murotani, K.; Nomura, T.; Ito, K.; Ohata, C.; Yamazaki, F.; Miyagi, T.; Takahashi, H.; Okubo, Y.; et al. Utility of the Dermatology Life Quality Index at initiation or switching of biologics in real-life Japanese patients with plaque psoriasis: Results from the ProLOGUE study. J. Dermatol. Sci. 2021, 101, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Honma, M.; Cai, Z.; Burge, R.; Zhu, B.; Yotsukura, S.; Torisu-Itakura, H. Relationship Between Rapid Skin Clearance and Quality of Life Benefit: Post Hoc Analysis of Japanese Patients with Moderate-to-Severe Psoriasis Treated with Ixekizumab (UNCOVER-J). Dermatol. Ther. 2020, 10, 1397–1404. [Google Scholar] [CrossRef] [PubMed]
- Meneguin, S.; de Godoy, N.A.; Pollo, C.F.; Miot, H.A.; de Oliveira, C. Quality of life of patients living with psoriasis: A qualitative study. BMC Dermatol. 2020, 20, 4–9. [Google Scholar] [CrossRef]
- Hrehorów, E.; Salomon, J.; Matusiak, L.; Reich, A.; Szepietowski, J.C. Patients with psoriasis feel stigmatized. Acta Derm. Venereol. 2012, 92, 67–72. [Google Scholar] [PubMed] [Green Version]
- Alpsoy, E.; Polat, M.; FettahlıoGlu-Karaman, B.; Karadag, A.S.; Kartal-Durmazlar, P.; YalCın, B.; Emre, S.; Didar-Balcı, D.; Bilgic-Temel, A.; Arca, E.; et al. Internalized stigma in psoriasis: A multicenter study. J. Dermatol. 2017, 44, 885–891. [Google Scholar] [CrossRef]
- Pearl, R.L.; Wan, M.T.; Takeshita, J.; Gelfand, J.M. Stigmatizing attitudes toward persons with psoriasis among laypersons and medical students. J. Am. Acad. Dermatol. 2019, 80, 1556–1563. [Google Scholar] [CrossRef]
- Armstrong, A.W.; Read, C. Pathophysiology, Clinical Presentation, and Treatment of Psoriasis: A Review. JAMA 2020, 323, 1945–1960. [Google Scholar] [CrossRef]
- Ogawa, E.; Sato, Y.; Minagawa, A.; Okuyama, R. Pathogenesis of psoriasis and development of treatment. J. Dermatol. 2018, 45, 264–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harden, J.L.; Krueger, J.G.; Bowcock, A.M. The immunogenetics of Psoriasis: A comprehensive review. J. Autoimmun. 2015, 64, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Egeberg, A.; Gisondi, P.; Carrascosa, J.M.; Warren, R.B.; Mrowietz, U. The role of the interleukin-23/Th17 pathway in cardiometabolic comorbidity associated with psoriasis. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 1695–1706. [Google Scholar] [CrossRef]
- Boehncke, W.; Boehncke, S.; Tobin, A.; Kirby, B. The ‘psoriatic march’: A concept of how severe psoriasis may drive cardiovascular comorbidity. Exp. Dermatol. 2011, 20, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Honma, M.; Hayashi, K. Psoriasis: Recent progress in molecular-targeted therapies. J. Dermatol. 2021, 48, 761–777. [Google Scholar] [CrossRef]
- Robb, M.A.; McInnes, P.M.; Califf, R.M. Biomarkers and surrogate endpoints: Developing common terminology and definitions. JAMA 2016, 315, 1107–1108. [Google Scholar] [CrossRef]
- Brembilla, N.C.; Senra, L.; Boehncke, W.H. The IL-17 family of cytokines in psoriasis: IL-17A and beyond. Front. Immunol. 2018, 9, 1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilliet, M.; Lande, R. Antimicrobial peptides and self-DNA in autoimmune skin inflammation. Curr. Opin. Immunol. 2008, 20, 401–407. [Google Scholar] [CrossRef]
- Takahashi, T.; Yamasaki, K. Psoriasis and antimicrobial peptides. Int. J. Mol. Sci. 2020, 21, 6791. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Bai, Y.P. Dendritic cells: The driver of psoriasis. J. Dermatol. 2020, 47, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Collamer, A.N.; Guerrero, K.T.; Henning, J.S.; Battafarano, D.F. Psoriatic skin lesions induced by tumor necrosis factor antagonist therapy: A literature review and potential mechanisms of action. Arthritis Rheum. 2008, 59, 996–1001. [Google Scholar] [CrossRef] [PubMed]
- Sakkas, L.I.; Mavropoulos, A.; Perricone, C.; Bogdanos, D.P. IL-35: A new immunomodulator in autoimmune rheumatic diseases Treg Breg. Immunol. Res. 2018, 66, 305–312. [Google Scholar] [CrossRef]
- Teng, M.W.L.; Bowman, E.P.; Mcelwee, J.J.; Smyth, M.J.; Casanova, J.; Cooper, A.M.; Cua, D.J. IL-12 and IL-23 cytokines: From discovery to targeted therapies for immune-mediated inflammatory diseases. Nat. Med. 2015, 21, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Trepicchio, W.L.; Oestreicher, J.L.; Pittman, D.; Wang, F.; Chamian, F.; Dhodapkar, M.; Krueger, J.G. Increased Expression of Interleukin 23 p19 and p40 in Lesional Skin of Patients with Psoriasis Vulgaris. J. Exp. Med. 2004, 199, 125–130. [Google Scholar] [CrossRef]
- Shibata, A.; Sugiura, K.; Furuta, Y.; Mukumoto, Y.; Kaminuma, O.; Akiyama, M. Toll-like receptor 4 antagonist TAK-242 inhibits autoinflammatory symptoms in DITRA. J. Autoimmun. 2017, 80, 28–38. [Google Scholar] [CrossRef]
- Yoon, J.; Leyva Castillo, J.M.; Wang, G.; Galand, C.; Oyoshi, M.K.; Kumar, L.; Hoff, S.; He, R.; Chervonsky, A.; Oppenheim, J.J.; et al. IL-23 induced in keratinocytes by endogenous TLR4 ligands polarizes dendritic cells to drive IL-22 responses to skin immunization. J. Exp. Med. 2016, 213, 2147–2166. [Google Scholar] [CrossRef]
- Li, H.; Yao, Q.; Mariscal, A.G.; Wu, X.; Hülse, J.; Pedersen, E.; Helin, K.; Waisman, A.; Vinkel, C.; Thomsen, S.F.; et al. Epigenetic control of IL-23 expression in keratinocytes is important for chronic skin inflammation. Nat. Commun. 2018, 9, 1420. [Google Scholar] [CrossRef]
- Koenen, H.J.P.M.; Smeets, R.L.; Vink, P.M.; Van Rijssen, E.; Boots, A.M.H.; Joosten, I. Human CD25 high Foxp3 pos regulatory T cells differentiate into IL-17—Producing cells. Blood 2008, 112, 2340–2353. [Google Scholar] [CrossRef] [Green Version]
- Bovenschen, H.J.; Van De Kerkhof, P.C.; Van Erp, P.E.; Woestenenk, R.; Joosten, I.; Koenen, H.J.P.M. Foxp3 regulatory T cells of psoriasis patients easily differentiate into IL-17A-producing cells and are found in lesional skin. J. Investig. Dermatol. 2011, 131, 1853–1860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mylonas, A.; Conrad, C. Psoriasis: Classical vs. Paradoxical. the yin-yang of TNF and Type i interferon. Front. Immunol. 2018, 9, 2746. [Google Scholar] [CrossRef] [PubMed]
- Krueger, G.; Callis, K. Potential of Tumor Necrosis Factor Inhibitors in Psoriasis and Psoriatic Arthritis. Arch. Dermatol. 2004, 140, 218–225. [Google Scholar] [CrossRef]
- Kalliolias, G.D.; Ivashkiv, L.B.; Program, T.D. TNF biology, pathogenic mechanisms and emerging therapeutic strategies. Nat. Rev. Rheumatol. 2016, 12, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, K.; Takeuchi, Y.; Hirota, K. The pathogenicity of Th17 cells in autoimmune diseases. Semin. Immunopathol. 2019, 41, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, N.; Ueno, H. Regulation of human helper T cell subset differentiation by cytokines. Curr. Opin. Immunol. 2015, 34, 130–136. [Google Scholar] [CrossRef] [Green Version]
- Amatya, N.; Garg, A.V.; Gaffen, S.L. IL-17 Signaling: The Yin and the Yang. Trends Immunol. 2017, 38, 310–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGeachy, M.J.; Cua, D.J.; Gaffen, S.L. The IL-17 Family of Cytokines in Health and Disease. Immunity 2019, 50, 892–906. [Google Scholar] [CrossRef]
- Monin, L.; Gaffen, S.L. Interleukin 17 Family Cytokines: Signaling and Therapeutic Implications. Cold Spring Harb. Perspect. Biol. 2018, 10, a028522. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Huang, J.; Zhao, X.; Lu, H.; Wang, W.; Yang, X.O.; Shi, Y.; Wang, X.; Lai, Y.; Dong, C. Interleukin-17 receptor D constitutes an alternative receptor for interleukin-17A important in psoriasis-like skin inflammation. Sci. Immunol. 2019, 4, eaau9657. [Google Scholar] [CrossRef]
- Matsuzaki, G.; Umemura, M. Interleukin-17 family cytokines in protective immunity against infections: Role of hematopoietic cell-derived and non-hematopoietic cell-derived interleukin-17s. Microbiol. Immunol. 2018, 62, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.S.; Tato, C.M.; Joyce-Shaikh, B.; Gulan, F.; Cayatte, C.; Chen, Y.; Blumenschein, W.M.; Judo, M.; Ayanoglu, G.; McClanahan, T.K.; et al. Interleukin-23-Independent IL-17 Production Regulates Intestinal Epithelial Permeability. Immunity 2015, 43, 727–738. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, J.R.; Zhang, Y.; Brown, W.A.; Smith, C.L.; Byrne, F.R.; Fiorino, M.; Stevens, E.; Bigler, J.; Davis, J.A.; Rottman, J.B.; et al. Differential Roles for Interleukin-23 and Interleukin-17 in Intestinal Immunoregulation. Immunity 2015, 43, 739–750. [Google Scholar] [CrossRef] [Green Version]
- McGonagle, D.G.; McInnes, I.B.; Kirkham, B.W.; Sherlock, J.; Moots, R. The role of IL-17A in axial spondyloarthritis and psoriatic arthritis: Recent advances and controversies. Ann. Rheum. Dis. 2019, 78, 1167–1178. [Google Scholar] [CrossRef] [Green Version]
- Cheuk, S.; Wiken, M.; Blomqvist, L.; Nylen, S.; Talme, T.; Stahle, M.; Eidsmo, L.; Wikén, M.; Blomqvist, L.; Nylén, S.; et al. Epidermal Th22 and Tc17 Cells Form a Localized Disease Memory in Clinically Healed Psoriasis. J. Immunol. 2014, 192, 3111–3120. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Tian, T.; Park, C.O.; Lofftus, S.Y.; Mei, S.; Liu, X.; Luo, C.; O’Malley, J.T.; Gehad, A.; Teague, J.E.; et al. Survival of tissue-resident memory T cells requires exogenous lipid uptake and metabolism. Nature 2017, 543, 252–256. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Oh, C.H.; Jeon, J.; Baek, Y.; Ahn, J.; Kim, D.J.; Lee, H.S.; Correa, J.; Sua, M.; Lowes, M.A.; et al. Molecular phenotyping small (Asian) versus large (Western) plaque psoriasis shows common activation of IL-17 pathway genes but different regulatory gene sets. J. Investig. Dermatol. 2016, 136, 161–172. [Google Scholar] [CrossRef] [Green Version]
- Imafuku, S.; Honma, M.; Okubo, Y.; Komine, M.; Ohtsuki, M.; Morita, A.; Seko, N.; Kawashima, N.; Ito, S.; Shima, T.; et al. Efficacy and safety of secukinumab in patients with generalized pustular psoriasis: A 52-week analysis from phase III open-label multicenter Japanese study. J. Dermatol. 2016, 43, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Mrowietz, U.; Bachelez, H.; Burden, A.D.; Rissler, M.; Sieder, C.; Orsenigo, R.; Chaouche-Teyara, K. Secukinumab for moderate-to-severe palmoplantar pustular psoriasis: Results of the 2PRECISE study. J. Am. Acad. Dermatol. 2019, 80, 1344–1352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honma, M.; Nozaki, H.; Hayashi, K.; Iinuma, S.; Ishida-Yamamoto, A. Palmoplantar pustulosis emerged on a case of generalized pustular psoriasis successfully treated by secukinumab. J. Dermatol. 2019, 46, e468–e469. [Google Scholar] [CrossRef] [PubMed]
- Zlotnik, A.; Yoshie, O. The Chemokine Superfamily Revisited. Immunity 2012, 36, 705–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schutyser, E.; Struyf, S.; Van Damme, J. The CC chemokine CCL20 and its receptor CCR6. Cytokine Growth Factor Rev. 2003, 14, 409–426. [Google Scholar] [CrossRef]
- Meitei, H.T.; Jadhav, N.; Lal, G. CCR6-CCL20 axis as a therapeutic target for autoimmune diseases. Autoimmun. Rev. 2021, 20, 102846. [Google Scholar] [CrossRef]
- Harper, E.G.; Guo, C.; Rizzo, H.; Lillis, J.V.; Kurtz, S.E.; Skorcheva, I.; Purdy, D.; Fitch, E.; Iordanov, M.; Blauvelt, A. Th17 cytokines stimulate CCL20 expression in keratinocytes in vitro and in vivo: Implications for psoriasis pathogenesis. J. Investig. Dermatol. 2009, 129, 2175–2183. [Google Scholar] [CrossRef] [Green Version]
- Chiricozzi, A.; Guttman-Yassky, E.; Suárez-Farĩas, M.; Nograles, K.E.; Tian, S.; Cardinale, I.; Chimenti, S.; Krueger, J.G. Integrative responses to IL-17 and TNF-α in human keratinocytes account for key inflammatory pathogenic circuits in psoriasis. J. Investig. Dermatol. 2011, 131, 677–687. [Google Scholar] [CrossRef]
- Furue, K.; Ito, T.; Tanaka, Y.; Yumine, A.; Hashimoto-Hachiya, A.; Takemura, M.; Murata, M.; Yamamura, K.; Tsuji, G.; Furue, M. Cyto/chemokine profile of in vitro scratched keratinocyte model: Implications of significant upregulation of CCL20, CXCL8 and IL36G in Koebner phenomenon. J. Dermatol. Sci. 2019, 94, 244–251. [Google Scholar] [CrossRef] [Green Version]
- Mabuchi, T.; Singh, T.P.; Takekoshi, T.; Jia, G.F.; Wu, X.; Kao, M.C.; Weiss, I.; Farber, J.M.; Hwang, S.T. CCR6 is required for epidermal trafficking of γδ-T cells in an IL-23-Induced model of psoriasiform dermatitis. J. Investig. Dermatol. 2013, 133, 164–171. [Google Scholar] [CrossRef] [Green Version]
- Hedrick, M.N.; Lonsdorf, A.S.; Shirakawa, A.K.; Lee, C.C.R.; Liao, F.; Singh, S.P.; Zhang, H.H.; Grinberg, A.; Love, P.E.; Hwang, S.T.; et al. CCR6 is required for IL-23-induced psoriasis-like inflammation in mice. J. Clin. Investig. 2009, 119, 2317–2329. [Google Scholar] [CrossRef] [Green Version]
- Getschman, A.E.; Imai, Y.; Larsen, O.; Peterson, F.C.; Wu, X.; Rosenkilde, M.M.; Hwang, S.T.; Volkman, B.F. Protein engineering of the chemokine CCL20 prevents psoriasiform dermatitis in an IL-23–dependent murine model. Proc. Natl. Acad. Sci. USA 2017, 114, 12460–12465. [Google Scholar] [CrossRef] [Green Version]
- Furue, K.; Ito, T.; Tsuji, G.; Nakahara, T.; Furue, M. The CCL20 and CCR6 axis in psoriasis. Scand. J. Immunol. 2020, 91, e12846. [Google Scholar] [CrossRef] [Green Version]
- Mabuchi, T.; Chang, T.W.; Quinter, S.; Hwang, S.T. Chemokine receptors in the pathogenesis and therapy of psoriasis. J. Dermatol. Sci. 2012, 65, 4–11. [Google Scholar] [CrossRef]
- Jensen, P.; Skov, L. Psoriasis and Obesity. Dermatology 2017, 232, 633–639. [Google Scholar] [CrossRef]
- Huang, Y.H.; Yang, L.C.; Hui, R.Y.; Chang, Y.C.; Yang, Y.W.; Yang, C.H.; Chen, Y.H.; Chung, W.H.; Kuan, Y.Z.; Chiu, C.S. Relationships between obesity and the clinical severity of psoriasis in Taiwan. J. Eur. Acad. Dermatol. Venereol. 2010, 24, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Naldi, L.; Conti, A.; Cazzaniga, S.; Patrizi, A.; Pazzaglia, M.; Lanzoni, A.; Veneziano, L.; Pellacani, G.; Miglietta, R.; Padalino, C.; et al. Diet and physical exercise in psoriasis: A randomized controlled trial. Br. J. Dermatol. 2014, 170, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Jensen, P.; Christensen, R.; Zachariae, C.; Geiker, N.R.W.; Schaadt, B.K.; Stender, S.; Hansen, P.R.; Astrup, A.; Skov, L. Long-term effects of weight reduction on the severity of psoriasis in a cohort derived from a randomized trial: A prospective observational follow-up study. Am. J. Clin. Nutr. 2016, 104, 259–265. [Google Scholar] [CrossRef]
- Mahil, S.K.; McSweeney, S.M.; Kloczko, E.; McGowan, B.; Barker, J.N.; Smith, C.H. Does weight loss reduce the severity and incidence of psoriasis or psoriatic arthritis? A Critically Appraised Topic. Br. J. Dermatol. 2019, 181, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Castaldo, G.; Rastrelli, L.; Galdo, G.; Molettieri, P.; Rotondi Aufiero, F.; Cereda, E. Aggressive weight-loss program with a ketogenic induction phase for the treatment of chronic plaque psoriasis: A proof-of-concept, single-arm, open-label clinical trial. Nutrition 2020, 74, 110757. [Google Scholar] [CrossRef] [PubMed]
- Jensen, P.; Zachariae, C.; Christensen, R.; Geiker, N.R.W.; Schaadt, B.K.; Stender, S.; Hansen, P.R.; Astrup, A.; Skov, L. Effect of weight loss on the severity of psoriasis: A randomized clinical study. JAMA Dermatol. 2013, 149, 795–801. [Google Scholar] [CrossRef] [Green Version]
- Debbaneh, M.; Millsop, J.W.; Bhatia, B.K.; Koo, J.; Liao, W. Diet and psoriasis, part I: Impact of weight loss interventions. J. Am. Acad. Dermatol. 2014, 71, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Sako, E.Y.; Famenini, S.; Wu, J.J. Bariatric surgery and psoriasis. J. Am. Acad. Dermatol. 2014, 70, 774–779. [Google Scholar] [CrossRef]
- Coimbra, S.; Catarino, C.; Santos-Silva, A. The triad psoriasis–obesity–adipokine profile. J. Eur. Acad. Dermatol. Venereol. 2016, 30, 1876–1885. [Google Scholar] [CrossRef]
- Gerdes, S.; Rostami-Yazdi, M.; Mrowietz, U. Adipokines and psoriasis. Exp. Dermatol. 2011, 20, 81–87. [Google Scholar] [CrossRef]
- Wong, Y.; Nakamizo, S.; Tan, K.J.; Kabashima, K. An update on the role of adipose tissues in psoriasis. Front. Immunol. 2019, 10, 1507. [Google Scholar] [CrossRef]
- Fantuzzi, G. Adipose tissue, adipokines, and inflammation. J. Allergy Clin. Immunol. 2005, 115, 911–919. [Google Scholar] [CrossRef]
- Pou, K.M.; Massaro, J.M.; Hoffmann, U.; Vasan, R.S.; Maurovich-Horvat, P.; Larson, M.G.; Keaney, J.F.; Meigs, J.B.; Lipinska, I.; Kathiresan, S.; et al. Visceral and subcutaneous adipose tissue volumes are cross-sectionally related to markers of inflammation and oxidative stress: The Framingham Heart Study. Circulation 2007, 116, 1234–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexopoulos, N.; Katritsis, D.; Raggi, P. Visceral adipose tissue as a source of inflammation and promoter of atherosclerosis. Atherosclerosis 2014, 233, 104–112. [Google Scholar] [CrossRef]
- Carvalheira, J.B.C.; Qiu, Y.; Chawla, A. Blood spotlight on leukocytes and obesity. Blood 2013, 122, 3263–3267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, D.M.; Caldas, A.P.; Oliveira, L.L.; Bressan, J.; Hermsdorff, H.H. Saturated fatty acids trigger TLR4-mediated inflammatory response. Atherosclerosis 2016, 244, 211–215. [Google Scholar] [CrossRef]
- Kunz, M.; Simon, J.C.; Saalbach, A. Psoriasis: Obesity and Fatty Acids. Front. Immunol. 2019, 10, 1807. [Google Scholar] [CrossRef]
- Barra, N.G.; Henriksbo, B.D.; Anhê, F.F.; Schertzer, J.D. The NLRP3 inflammasome regulates adipose tissue metabolism. Biochem. J. 2020, 477, 1089–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheuk, S.; Eidsmo, L. The Skinny on Fat TRM Cells. Immunity 2017, 47, 1012–1014. [Google Scholar] [CrossRef] [Green Version]
- Han, S.J.; Glatman Zaretsky, A.; Andrade-Oliveira, V.; Collins, N.; Dzutsev, A.; Shaik, J.; Morais da Fonseca, D.; Harrison, O.J.; Tamoutounour, S.; Byrd, A.L.; et al. White Adipose Tissue Is a Reservoir for Memory T Cells and Promotes Protective Memory Responses to Infection. Immunity 2017, 47, 1154–1168. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, G.A.; Manfredi, A.A.; Maugeri, N. Misunderstandings between platelets and neutrophils build in chronic inflammation. Front. Immunol. 2019, 10, 2491. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.C.; Cheng, W.J.; Korinek, M.; Lin, C.Y.; Hwang, T.L. Neutrophils in Psoriasis. Front. Immunol. 2019, 10, 2376. [Google Scholar] [CrossRef] [PubMed]
- Sylman, J.L.; Mitrugno, A.; Atallah, M.; Tormoen, G.W.; Shatzel, J.J.; Yunga, S.T.; Wagner, T.H.; Leppert, J.T.; Mallick, P.; McCarty, O.J.T. The predictive value of inflammation-related peripheral blood measurements in cancer staging and prognosis. Front. Oncol. 2018, 8, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, X.; Li, D.; Wu, D.; Zhang, N. The Relationship between Hematological Indices and Autoimmune Rheumatic Diseases (ARDs), a Meta-Analysis. Sci. Rep. 2017, 7, 10833. [Google Scholar] [CrossRef] [Green Version]
- Asahina, A.; Kubo, N.; Umezawa, Y.; Honda, H.; Yanaba, K. Neutrophil-lymphocyte ratio, platelet-lymphocyte ratio and mean platelet volume in Japanese patients with psoriasis and psoriatic arthritis: Response to therapy with biologics. J. Dermatol. 2017, 44, 1112–1121. [Google Scholar] [CrossRef]
- Najar Nobari, N.; Shahidi Dadras, M.; Nasiri, S.; Abdollahimajd, F.; Gheisari, M. Neutrophil/platelet to lymphocyte ratio in monitoring of response to TNF-α inhibitors in psoriatic patients. Dermatol. Ther. 2020, 33, e13457. [Google Scholar] [CrossRef]
- Paliogiannis, P.; Satta, R.; Deligia, G.; Farina, G.; Bassu, S.; Mangoni, A.A.; Carru, C.; Zinellu, A. Associations between the neutrophil-to-lymphocyte and the platelet-to-lymphocyte ratios and the presence and severity of psoriasis: A systematic review and meta-analysis. Clin. Exp. Med. 2019, 19, 37–45. [Google Scholar] [CrossRef]
- Yilmaz, S.B.; Cicek, N.; Coskun, M.; Yegin, O.; Alpsoy, E. Serum and tissue levels of IL-17 in different clinical subtypes of psoriasis. Arch. Dermatol. Res. 2012, 304, 465–469. [Google Scholar] [CrossRef]
- Kolbinger, F.; Loesche, C.; Valentin, M.A.; Jiang, X.; Cheng, Y.; Jarvis, P.; Peters, T.; Calonder, C.; Bruin, G.; Polus, F.; et al. β-Defensin 2 is a responsive biomarker of IL-17A–driven skin pathology in patients with psoriasis. J. Allergy Clin. Immunol. 2017, 139, 923–932. [Google Scholar] [CrossRef] [Green Version]
- Gordon, K.B.; Armstrong, A.W.; Foley, P.; Song, M.; Shen, Y.K.; Li, S.; Muñoz-Elías, E.J.; Branigan, P.; Liu, X.; Reich, K. Guselkumab Efficacy after Withdrawal Is Associated with Suppression of Serum IL-23-Regulated IL-17 and IL-22 in Psoriasis: VOYAGE 2 Study. J. Investig. Dermatol. 2019, 139, 2437–2446. [Google Scholar] [CrossRef]
- Fitz, L.; Zhang, W.; Soderstrom, C.; Fraser, S.; Lee, J.; Quazi, A.; Wolk, R.; Mebus, C.A.; Valdez, H.; Berstein, G. Association between serum interleukin-17A and clinical response to tofacitinib and etanercept in moderate to severe psoriasis. Clin. Exp. Dermatol. 2018, 43, 790–797. [Google Scholar] [CrossRef] [PubMed]
- Garcet, S.; Nograles, K.; Correa da Rosa, J.; Schafer, P.H.; Krueger, J.G. Synergistic cytokine effects as apremilast response predictors in patients with psoriasis. J. Allergy Clin. Immunol. 2018, 142, 1010–1013. [Google Scholar] [CrossRef] [PubMed]
- Ishigame, H.; Kakuta, S.; Nagai, T.; Kadoki, M.; Nambu, A.; Komiyama, Y.; Fujikado, N.; Tanahashi, Y.; Akitsu, A.; Kotaki, H.; et al. Differential Roles of Interleukin-17A and -17F in Host Defense against Mucoepithelial Bacterial Infection and Allergic Responses. Immunity 2009, 30, 108–119. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.L.; Teijeiro, A.; Burén, S.; Tummala, K.S.; Yilmaz, M.; Waisman, A.; Theurillat, J.P.; Perna, C.; Djouder, N. Metabolic Inflammation-Associated IL-17A Causes Non-alcoholic Steatohepatitis and Hepatocellular Carcinoma. Cancer Cell 2016, 30, 161–175. [Google Scholar] [CrossRef]
- Von Stebut, E.; Boehncke, W.H.; Ghoreschi, K.; Gori, T.; Kaya, Z.; Thaci, D.; Schäffler, A. IL-17A in Psoriasis and Beyond: Cardiovascular and Metabolic Implications. Front. Immunol. 2020, 10, 3096. [Google Scholar] [CrossRef] [Green Version]
- Ikumi, K.; Odanaka, M.; Shime, H.; Imai, M.; Osaga, S.; Taguchi, O.; Nishida, E.; Hemmi, H.; Kaisho, T.; Morita, A.; et al. Hyperglycemia Is Associated with Psoriatic Inflammation in Both Humans and Mice. J. Investig. Dermatol. 2019, 139, 1329–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elnabawi, Y.A.; Dey, A.K.; Goyal, A.; Groenendyk, J.W.; Chung, J.H.; Belur, A.D.; Rodante, J.; Harrington, C.L.; Teague, H.L.; Baumer, Y.; et al. Coronary artery plaque characteristics and treatment with biologic therapy in severe psoriasis: Results from a prospective observational study. Cardiovasc. Res. 2019, 115, 721–728. [Google Scholar] [CrossRef]
- Rutz, S.; Wang, X.; Ouyang, W. The IL-20 subfamily of cytokines-from host defence to tissue homeostasis. Nat. Rev. Immunol. 2014, 14, 783–795. [Google Scholar] [CrossRef]
- Ouyang, W.; O’Garra, A. IL-10 Family Cytokines IL-10 and IL-22: From Basic Science to Clinical Translation. Immunity 2019, 50, 871–891. [Google Scholar] [CrossRef]
- Eyerich, K.; Dimartino, V.; Cavani, A. IL-17 and IL-22 in immunity: Driving protection and pathology. Eur. J. Immunol. 2017, 47, 607–614. [Google Scholar] [CrossRef] [Green Version]
- Honma, M.; Minami-Hori, M.; Takahashi, H.; Iizuka, H. Podoplanin expression in wound and hyperproliferative psoriatic epidermis: Regulation by TGF-β and STAT-3 activating cytokines, IFN-γ, IL-6, and IL-22. J. Dermatol. Sci. 2012, 65, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Guttman-Yassky, E.; Krueger, J.G. Atopic dermatitis and psoriasis: Two different immune diseases or one spectrum? Curr. Opin. Immunol. 2017, 48, 68–73. [Google Scholar] [CrossRef]
- Shimauchi, T.; Hirakawa, S.; Suzuki, T.; Yasuma, A.; Majima, Y.; Tatsuno, K.; Yagi, H.; Ito, T.; Tokura, Y. Serum interleukin-22 and vascular endothelial growth factor serve as sensitive biomarkers but not as predictors of therapeutic response to biologics in patients with psoriasis. J. Dermatol. 2013, 40, 805–812. [Google Scholar] [CrossRef]
- Philipp, S.; Menter, A.; Nikkels, A.F.; Barber, K.; Landells, I.; Eichenfield, L.F.; Song, M.; Randazzo, B.; Li, S.; Hsu, M.C.; et al. Ustekinumab for the treatment of moderate-to-severe plaque psoriasis in paediatric patients (≥6 to <12 years of age): Efficacy, safety, pharmacokinetic and biomarker results from the open-label CADMUS Jr study. Br. J. Dermatol. 2020, 183, 664–672. [Google Scholar] [CrossRef] [Green Version]
- Kragstrup, T.W.; Andersen, T.; Heftdal, L.D.; Hvid, M.; Gerwien, J.; Sivakumar, P.; Taylor, P.C.; Senolt, L.; Deleuran, B. The IL-20 cytokine family in rheumatoid arthritis and spondyloarthritis. Front. Immunol. 2018, 9, 2226. [Google Scholar] [CrossRef]
- Konrad, R.J.; Higgs, R.E.; Rodgers, G.H.; Ming, W.; Qian, Y.W.; Bivi, N.; Mack, J.K.; Siegel, R.W.; Nickoloff, B.J. Assessment and Clinical Relevance of Serum IL-19 Levels in Psoriasis and Atopic Dermatitis Using a Sensitive and Specific Novel Immunoassay. Sci. Rep. 2019, 9, 5211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Autieri, M.V. IL-19 and other IL-20 family member cytokines in vascular inflammatory diseases. Front. Immunol. 2018, 9, 700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabre, T.; Molina, M.F.; Soucy, G.; Goulet, J.P.; Willems, B.; Villeneuve, J.P.; Bilodeau, M.; Shoukry, N.H. Type 3 cytokines IL-17A and IL-22 drive TGF-–dependent liver fibrosis. Sci. Immunol. 2018, 3, eaar7754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, P.T.; Fallon, P.G. The emergence of the IL-36 cytokine family as novel targets for inflammatory diseases. Ann. N. Y. Acad. Sci. 2016, 1417, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Yi, G.; Ybe, J.A.; Saha, S.S.; Caviness, G.; Raymond, E.; Ganesan, R.; Mbow, M.L.; Kao, C.C. Structural and functional attributes of the interleukin-36 receptor. J. Biol. Chem. 2016, 291, 16597–16609. [Google Scholar] [CrossRef] [Green Version]
- Fields, J.K.; Günther, S.; Sundberg, E.J. Structural basis of IL-1 family cytokine signaling. Front. Immunol. 2019, 10, 1412. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Huard, A.; Mora, J.; da Silva, P.; Brüne, B.; Weigert, A. IL-36 family cytokines in protective versus destructive inflammation. Cell. Signal. 2020, 75, 109773. [Google Scholar] [CrossRef] [PubMed]
- Furue, K.; Yamamura, K.; Tsuji, G.; Mitoma, C.; Uchi, H.; Nakahara, T.; Kido-Nakahara, M.; Kadono, T.; Furue, M. Highlighting interleukin-36 signalling in plaque psoriasis and pustular psoriasis. Acta Derm. Venereol. 2018, 98, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Mercurio, L.; Id, C.M.F.; Capriotti, L.; Scarponi, C.; Facchiano, F.; Morelli, M.; Rossi, S.; Pagnanelli, G.; Id, C.A.; Cavani, A.; et al. Interleukin (IL)-17/IL-36 axis participates to the crosstalk between endothelial cells and keratinocytes during inflammatory skin responses. PLoS ONE 2020, 15, e0222969. [Google Scholar] [CrossRef]
- Buhl, A.L.; Wenzel, J. Interleukin-36 in infectious and inflammatory skin diseases. Front. Immunol. 2019, 10, 1162. [Google Scholar] [CrossRef]
- Madonna, S.; Girolomoni, G.; Dinarello, C.A.; Albanesi, C. The significance of Il-36 hyperactivation and Il-36R targeting in psoriasis. Int. J. Mol. Sci. 2019, 20, 3318. [Google Scholar] [CrossRef] [Green Version]
- Pfaff, C.M.; Marquardt, Y.; Fietkau, K.; Baron, J.M.; Lüscher, B. The psoriasis-associated IL-17A induces and cooperates with IL-36 cytokines to control keratinocyte differentiation and function. Sci. Rep. 2017, 7, 15631. [Google Scholar] [CrossRef] [PubMed]
- D’Erme, A.M.; Wilsmann-Theis, D.; Wagenpfeil, J.; Hölzel, M.; Ferring-Schmitt, S.; Sternberg, S.; Wittmann, M.; Peters, B.; Bosio, A.; Bieber, T.; et al. IL-36γ (IL-1F9) Is a Biomarker for Psoriasis Skin Lesions. J. Investig. Dermatol. 2015, 135, 1025–1032. [Google Scholar] [CrossRef] [Green Version]
- Braegelmann, J.; D’Erme, A.M.; Akmal, S.; Maier, J.; Braegelmann, C.; Wenzel, J. Interleukin-36γ (IL-1F9) identifies psoriasis among patients with erythroderma. Acta Derm. Venereol. 2016, 96, 386–387. [Google Scholar] [CrossRef] [Green Version]
- Raychaudhuri, S.P.; Jiang, W.Y.; Farber, E.M. Cellular localization of fractalkine at sites of inflammation: Antigen-presenting cells in psoriasis express high levels of fractalkine. Br. J. Dermatol. 2001, 144, 1105–1113. [Google Scholar] [CrossRef]
- Fraticelli, P.; Sironi, M.; Bianchi, G.; D’Ambrosio, D.; Albanesi, C.; Stoppacciaro, A.; Chieppa, M.; Allavena, P.; Ruco, L.; Girolomoni, G.; et al. Fractalkine (CX3CL1) as an amplification circuit of polarized Th1 responses. J. Clin. Investig. 2001, 107, 1173–1181. [Google Scholar] [CrossRef]
- Sugaya, M.; Nakamura, K.; Mitsui, H.; Takekoshi, T.; Saeki, H.; Tamaki, K. Human keratinocytes express fractalkine/CX3CL1. J. Dermatol. Sci. 2003, 31, 179–187. [Google Scholar] [CrossRef]
- Plant, D.; Young, H.S.; Watson, R.E.B.; Worthington, J.; Griffiths, C.E.M. The CX3CL1-CX3CR1 system and psoriasis. Exp. Dermatol. 2006, 15, 900–903. [Google Scholar] [CrossRef]
- Morimura, S.; Oka, T.; Sugaya, M.; Sato, S. CX3CR1 deficiency attenuates imiquimod-induced psoriasis-like skin inflammation with decreased M1 macrophages. J. Dermatol. Sci. 2016, 82, 175–188. [Google Scholar] [CrossRef]
- Congjun, J.; Yanmei, Z.; Huiling, J.; Zhen, Y.; Shuo, L. Elevated local and serum CX3CL1(Fractalkine) Expression and its association with disease severity in patients with psoriasis. Ann. Clin. Lab. Sci. 2015, 45, 556–561. [Google Scholar]
- Echigo, T.; Hasegawa, M.; Shimada, Y.; Takehara, K.; Sato, S. Expression of fractalkine and its receptor, CX3CR1, in atopic dermatitis: Possible contribution to skin inflammation. J. Allergy Clin. Immunol. 2004, 113, 940–948. [Google Scholar] [CrossRef]
- Teupser, D.; Pavlides, S.; Tan, M.; Gutierrez-Ramos, J.C.; Kolbeck, R.; Breslow, J.L. Major reduction of antherosclerosis in fractalkine (CX3CL1)-deficient mice is at the brachiocephalic artery, not the aortic root. Proc. Natl. Acad. Sci. USA 2004, 101, 17795–17800. [Google Scholar] [CrossRef] [Green Version]
- Hughes, C.E.; Nibbs, R.J.B. A guide to chemokines and their receptors. FEBS J. 2018, 285, 2944–2971. [Google Scholar] [CrossRef]
- Saeki, H.; Tamaki, K. Thymus and activation regulated chemokine (TARC)/CCL17 and skin diseases. J. Dermatol. Sci. 2006, 43, 75–84. [Google Scholar] [CrossRef]
- Rottman, J.B.; Smith, T.L.; Ganley, K.G.; Kikuchi, T.; Krueger, J.G. Potential role of the chemokine receptors CXCR3, CCR4, and the integrin αEβ7 in the pathogenesis of psoriasis vulgaris. Lab. Investig. 2001, 81, 335–347. [Google Scholar] [CrossRef] [Green Version]
- Kakinuma, T.; Nakamura, K.; Wakugawa, M.; Mitsui, H.; Tada, Y.; Saeki, H.; Torii, H.; Asahina, A.; Onai, N.; Matsushima, K.; et al. Thymus and activation-regulated chemokine in atopic dermatitis: Serum thymus and activation-regulated chemokine level is closely related with disease activity. J. Allergy Clin. Immunol. 2001, 107, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, T.; Honma, M.; Iinuma, S.; Iwasaki, T.; Takahashi, H.; Ishida-Yamamoto, A. Alteration of serum thymus and activation-regulated chemokine level during biologic therapy for psoriasis: Possibility as a marker reflecting favorable response to anti-interleukin-17A agents. J. Dermatol. 2018, 45, 710–714. [Google Scholar] [CrossRef] [PubMed]
- Bernink, J.H.; Ohne, Y.; Teunissen, M.B.M.; Wang, J.; Wu, J.; Krabbendam, L.; Guntermann, C.; Volckmann, R.; Koster, J.; van Tol, S.; et al. c-Kit-positive ILC2s exhibit an ILC3-like signature that may contribute to IL-17-mediated pathologies. Nat. Immunol. 2019, 20, 992–1003. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, Y.; Kamata, M.; Shimizu, T.; Nagata, M.; Fukaya, S.; Hayashi, K.; Fukuyasu, A.; Tanaka, T.; Ishikawa, T.; Ohnishi, T.; et al. Thymus and activation-regulated chemokine (TARC) in patients with psoriasis: Increased serum TARC levels in patients with generalized pustular psoriasis. J. Dermatol. 2020, 47, 1149–1156. [Google Scholar] [CrossRef]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef]
- Fasshauer, M.; Blüher, M. Adipokines in health and disease. Trends Pharmacol. Sci. 2015, 36, 461–470. [Google Scholar] [CrossRef]
- Francisco, V.; Ruiz-Fernández, C.; Pino, J.; Mera, A.; González-Gay, M.A.; Gómez, R.; Lago, F.; Mobasheri, A.; Gualillo, O. Adipokines: Linking metabolic syndrome, the immune system, and arthritic diseases. Biochem. Pharmacol. 2019, 165, 196–206. [Google Scholar] [CrossRef]
- Versini, M.; Jeandel, P.Y.; Rosenthal, E.; Shoenfeld, Y. Obesity in autoimmune diseases: Not a passive bystander. Autoimmun. Rev. 2014, 13, 981–1000. [Google Scholar] [CrossRef]
- Bai, F.; Zheng, W.; Dong, Y.; Wang, J.; Garstka, M.A.; Li, R.; An, J.; Ma, H. Serum levels of adipokines and cytokines in psoriasis patients: A systematic review and meta-analysis. Oncotarget 2018, 9, 1266–1278. [Google Scholar] [CrossRef] [Green Version]
- Steppan, C.M.; Lazar, M.A.; Lazar, M.A. Resistin and obesity-associated insulin resistance. Trends Endocrinol. Metab. 2002, 13, 18–23. [Google Scholar] [CrossRef]
- Takahashi, H.; Tsuji, H.; Honma, M.; Ishida-Yamamoto, A.; Iizuka, H. Increased plasma resistin and decreased omentin levels in Japanese patients with psoriasis. Arch. Dermatol. Res. 2013, 305, 113–116. [Google Scholar] [CrossRef]
- Kyriakou, A.; Patsatsi, A.; Sotiriadis, D.; Goulis, D.G. Effects of treatment for psoriasis on circulating levels of leptin, adiponectin and resistin: A systematic review and meta-analysis. Br. J. Dermatol. 2018, 179, 273–281. [Google Scholar] [CrossRef]
- Pina, T.; Genre, F.; Lopez-Mejias, R.; Armesto, S.; Ubilla, B.; Mijares, V.; Dierssen-Sotos, T.; Gonzalez-Lopez, M.A.; Gonzalez-Vela, M.C.; Blanco, R.; et al. Relationship of Leptin with adiposity and inflammation and Resistin with disease severity in Psoriatic patients undergoing anti-TNF-alpha therapy. J. Eur. Acad. Dermatol. Venereol. 2015, 29, 1995–2001. [Google Scholar] [CrossRef]
- Robati, R.M.; Partovi-Kia, M.; Haghighatkhah, H.R.; Younespour, S.; Abdollahimajd, F. Increased serum leptin and resistin levels and increased carotid intima-media wall thickness in patients with psoriasis: Is psoriasis associated with atherosclerosis? J. Am. Acad. Dermatol. 2014, 71, 642–648. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, B.J.; Scalia, R.G.; Ma, X.L. Protective vascular and myocardial effects of adiponectin. Nat. Clin. Pract. Cardiovasc. Med. 2009, 6, 27–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibata, S.; Tada, Y.; Hau, C.S.; Mitsui, A.; Kamata, M.; Asano, Y.; Sugaya, M.; Kadono, T.; Masamoto, Y.; Kurokawa, M.; et al. Adiponectin regulates psoriasiform skin inflammation by suppressing IL-17 production from γδ-T cells. Nat. Commun. 2015, 6, 7687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coimbra, S.; Oliveira, H.; Reis, F.; Belo, L.; Rocha, S.; Quintanilha, A.; Figueiredo, A.; Teixeira, F.; Castro, E.; Rocha-Pereira, P.; et al. Circulating adipokine levels in Portuguese patients with psoriasis vulgaris according to body mass index, severity and therapy. J. Eur. Acad. Dermatol. Venereol. 2010, 24, 1386–1394. [Google Scholar] [CrossRef]
- Boehncke, S.; Salgo, R.; Garbaraviciene, J.; Beschmann, H.; Hardt, K.; Diehl, S.; Fichtlscherer, S.; Thaçi, D.; Boehncke, W.H. Effective continuous systemic therapy of severe plaque-type psoriasis is accompanied by amelioration of biomarkers of cardiovascular risk: Results of a prospective longitudinal observational study. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 1187–1193. [Google Scholar] [CrossRef]
- Gerdes, S.; Pinter, A.; Biermann, M.; Papavassilis, C.; Reinhardt, M. Adiponectin levels in a large pooled plaque psoriasis study population. J. Dermatolog. Treat. 2020, 31, 531–534. [Google Scholar] [CrossRef] [Green Version]
- Johnson, C.M.; Fitch, K.; Merola, J.F.; Han, J.; Qureshi, A.A.; Li, W.Q. Plasma levels of tumour necrosis factor-α and adiponectin can differentiate patients with psoriatic arthritis from those with psoriasis. Br. J. Dermatol. 2019, 181, 379–380. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Gallo, R.L. Antimicrobial Peptides: Old Molecules with New Ideas. J. Investig. Dermatol. 2011, 132, 887–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazzaro, B.P.; Zasloff, M.; Rolff, J. Antimicrobial peptides: Application informed by evolution. Science 2020, 368, eaau5480. [Google Scholar] [CrossRef]
- Magana, M.; Pushpanathan, M.; Santos, A.L.; Leanse, L.; Fernandez, M.; Ioannidis, A.; Giulianotti, M.A.; Apidianakis, Y.; Bradfute, S.; Ferguson, A.L.; et al. The value of antimicrobial peptides in the age of resistance. Lancet Infect. Dis. 2020, 20, e216–e230. [Google Scholar] [CrossRef]
- Lai, Y.; Gallo, R.L. AMPed up immunity: How antimicrobial peptides have multiple roles in immune defense. Trends Immunol. 2009, 30, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Ali, R.S.; Falconer, A.; Ikram, M.; Bissett, C.E.; Cerio, R.; Quinn, A.G. Expression of the peptide antibiotics human β defensin-1 and human β defensin-2 in normal human skin. J. Investig. Dermatol. 2001, 117, 106–111. [Google Scholar] [PubMed] [Green Version]
- Jansen, P.A.M.; Rodijk-Olthuis, D.; Hollox, E.J.; Kamsteeg, M.; Tjabringa, G.S.; de Jongh, G.J.; van Vlijmen-Willems, I.M.J.J.; Bergboer, J.G.M.; van Rossum, M.M.; de Jong, E.M.G.J.; et al. β-Defensin-2 protein is a serum biomarker for disease activity in psoriasis and reaches biologically relevant concentrations in lesional skin. PLoS ONE 2009, 4, e4725. [Google Scholar] [CrossRef] [Green Version]
- Morita, A.; Tani, Y.; Matsumoto, K.; Yamaguchi, M.; Teshima, R.; Ohtsuki, M. Assessment of serum biomarkers in patients with plaque psoriasis on secukinumab. J. Dermatol. 2020, 47, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.; Sun, Z.; Chen, X.; Wang, Y.; Li, R.; Ji, S.; Zhao, Y. Serum Human Beta-Defensin-2 Is a Possible Biomarker for Monitoring Response to JAK Inhibitor in Psoriasis Patients. Dermatology 2017, 233, 164–169. [Google Scholar] [CrossRef]
- Gonzalez, L.L.; Garrie, K.; Turner, M.D. Role of S100 proteins in health and disease. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118677. [Google Scholar] [CrossRef]
- Büchau, A.S.; Gallo, R.L. Innate immunity and antimicrobial defense systems in psoriasis. Clin. Dermatol. 2007, 25, 616–624. [Google Scholar] [CrossRef] [Green Version]
- D’Amico, F.; Skarmoutsou, E.; Granata, M.; Trovato, C.; Rossi, G.A.; Mazzarino, M.C. S100A7: A rAMPing up AMP molecule in psoriasis. Cytokine Growth Factor Rev. 2016, 32, 97–104. [Google Scholar] [CrossRef]
- Maurelli, M.; Gisondi, P.; Danese, E.; Gelati, M.; Papagrigoraki, A.; del Giglio, M.; Lippi, G.; Girolomoni, G. Psoriasin (S100A7) is increased in the serum of patients with moderate-to-severe psoriasis. Br. J. Dermatol. 2020, 182, 1502–1503. [Google Scholar] [CrossRef] [PubMed]
- Awad, S.M.; Attallah, D.A.; Salama, R.H.; Mahran, A.M.; Abu El-Hamed, E. Serum levels of psoriasin (S100A7) and koebnerisin (S100A15) as potential markers of atherosclerosis in patients with psoriasis. Clin. Exp. Dermatol. 2018, 43, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Batycka-Baran, A.; Hattinger, E.; Zwicker, S.; Summer, B.; Zack Howard, O.M.; Thomas, P.; Szepietowski, J.C.; Ruzicka, T.; Prinz, J.C.; Wolf, R. Leukocyte-derived koebnerisin (S100A15) and psoriasin (S100A7) are systemic mediators of inflammation in psoriasis. J. Dermatol. Sci. 2015, 79, 214–221. [Google Scholar] [CrossRef]
- Takeda, A.; Higuchi, D.; Takahashi, T.; Ogo, M.; Baciu, P.; Goetinck, P.F.; Hibino, T. Overexpression of serpin squamous cell carcinoma antigens in psoriatic skin. J. Investig. Dermatol. 2002, 118, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, Y.; Yamaguchi, Y.; Komitsu, N.; Ohta, S.; Azuma, Y.; Izuhara, K.; Aihara, M. Elevation of serum squamous cell carcinoma antigen 2 in patients with psoriasis: Associations with disease severity and response to the treatment. Br. J. Dermatol. 2016, 174, 1327–1336. [Google Scholar] [CrossRef] [PubMed]
- Naher, L.; Kiyoshima, T.; Kobayashi, I.; Wada, H.; Nagata, K.; Fujiwara, H.; Ookuma, Y.F.; Ozeki, S.; Nakamura, S.; Sakai, H. STAT3 signal transduction through interleukin-22 in oral squamous cell carcinoma. Int. J. Oncol. 2012, 41, 1577–1586. [Google Scholar] [CrossRef] [Green Version]
- Mitsuishi, K.; Nakamura, T.; Sakata, Y.; Yuyama, N.; Arima, K.; Sugita, Y.; Suto, H.; Izuhara, K.; Ogawa, H. The squamous cell carcinoma antigens as relevant biomarkers of atopic dermatitis. Clin. Exp. Allergy 2005, 35, 1327–1333. [Google Scholar] [CrossRef]
- Okawa, T.; Yamaguchi, Y.; Kou, K.; Ono, J.; Azuma, Y.; Komitsu, N.; Inoue, Y.; Kohno, M.; Matsukura, S.; Kambara, T.; et al. Serum levels of squamous cell carcinoma antigens 1 and 2 reflect disease severity and clinical type of atopic dermatitis in adult patients. Allergol. Int. 2018, 67, 124–130. [Google Scholar] [CrossRef]
- Iizuka, H.; Takahashi, H.; Honma, M.; Ishida-Yamamoto, A. Unique keratinization process in psoriasis: Late differentiation markers are abolished because of the premature cell death. J. Dermatol. 2004, 31, 271–276. [Google Scholar] [CrossRef]
- Nakane, H.; Ishida-Yamamoto, A.; Takahashi, H.; Iizuka, H. Elafin, a secretory protein, is cross-linked into the cornified cell envelopes from the inside of psoriatic keratinocytes. J. Investig. Dermatol. 2002, 119, 50–55. [Google Scholar] [CrossRef] [Green Version]
- Nonomura, K.; Yamanishi, K.; Yasuno, H.; Nara, K.; Hirose, S. Up-regulation of elafin/SKALP gene expression in psoriatic epidermis. J. Investig. Dermatol. 1994, 103, 88–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sallenave, J.M. Secretory leukocyte protease inhibitor and elafin/trappin-2: Versatile mucosal antimicrobials and regulators of immunity. Am. J. Respir. Cell Mol. Biol. 2010, 42, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Elgharib, I.; Khashaba, S.A.; Elsaid, H.H.; Sharaf, M.M. Serum elafin as a potential inflammatory marker in psoriasis. Int. J. Dermatol. 2019, 58, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.R.; Newby, D.E.; Henriksen, P.A. Role of the endogenous elastase inhibitor, elafin, in cardiovascular injury: From epithelium to endothelium. Biochem. Pharmacol. 2012, 83, 695–704. [Google Scholar] [CrossRef]
- Shavadia, J.S.; Granger, C.B.; Alemayehu, W.; Westerhout, C.M.; Povsic, T.J.; Brener, S.J.; van Diepen, S.; Defilippi, C.; Armstrong, P.W. High-throughput targeted proteomics discovery approach and spontaneous reperfusion in ST-segment elevation myocardial infarction. Am. Heart J. 2020, 220, 137–144. [Google Scholar] [CrossRef]
- Wang, X.; Abraham, S.; McKenzie, J.A.G.; Jeffs, N.; Swire, M.; Tripathi, V.B.; Luhmann, U.F.O.; Lange, C.A.K.; Zhai, Z.; Arthur, H.M.; et al. LRG1 promotes angiogenesis by modulating endothelial TGF-β signalling. Nature 2013, 499, 306–311. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.J.; Hsieh, C.Y.; Shu, K.H.; Chen, I.Y.; Pan, S.Y.; Chuang, Y.F.; Chiu, Y.L.; Yang, W.S. Plasma Leucine-Rich α-2-Glycoprotein 1 Predicts Cardiovascular Disease Risk in End-Stage Renal Disease. Sci. Rep. 2020, 10, 5988. [Google Scholar] [CrossRef] [Green Version]
- Urushima, H.; Fujimoto, M.; Mishima, T.; Ohkawara, T.; Honda, H.; Lee, H.; Kawahata, H.; Serada, S.; Naka, T. Leucine-rich alpha 2 glycoprotein promotes Th17 differentiation and collagen-induced arthritis in mice through enhancement of TGF-β-Smad2 signaling in naïve helper T cells. Arthritis Res. Ther. 2017, 19, 137. [Google Scholar] [CrossRef]
- Nakajima, H.; Serada, S.; Fujimoto, M.; Naka, T.; Sano, S. Leucine-rich α-2 glycoprotein is an innovative biomarker for psoriasis. J. Dermatol. Sci. 2017, 86, 170–174. [Google Scholar] [CrossRef]
- Libreros, S.; Iragavarapu-Charyulu, V. YKL-40/CHI3L1 drives inflammation on the road of tumor progression. J. Leukoc. Biol. 2015, 98, 931–936. [Google Scholar] [CrossRef] [Green Version]
- Prakash, M.; Bodas, M.; Prakash, D.; Nawani, N.; Khetmalas, M.; Mandal, A.; Eriksson, C. Diverse pathological implications of YKL-40: Answers may lie in “outside-in” signaling. Cell. Signal. 2013, 25, 1567–1573. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Li, G.; Chang, D.; Su, X. YKL-40 as a novel biomarker in cardio-metabolic disorders and inflammatory diseases. Clin. Chim. Acta 2020, 511, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Imai, Y.; Tsuda, T.; Aochi, S.; Futatsugi-Yumikura, S.; Sakaguchi, Y.; Nakagawa, N.; Iwatsuki, K.; Yamanishi, K. YKL-40 (chitinase 3-like-1) as a biomarker for psoriasis vulgaris and pustular psoriasis. J. Dermatol. Sci. 2011, 64, 75–77. [Google Scholar] [CrossRef] [PubMed]
- Khashaba, S.A.; Attwa, E.; Said, N.; Ahmed, S.; Khattab, F. Serum YKL-40 and IL 17 in Psoriasis: Reliability as prognostic markers for disease severity and responsiveness to treatment. Dermatol. Ther. 2021, 34, e14606. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.F.; Attia, E.A.S.; Saad, A.A.; Sharara, M.; Fawzy, H.; El Nahrery, E.M.A. Serum YKL-40 in psoriasis with and without arthritis; Correlation with disease activity and high-resolution power Doppler ultrasonographic joint findings. J. Eur. Acad. Dermatol. Venereol. 2015, 29, 682–688. [Google Scholar] [CrossRef]
- Erfan, G.; Guzel, S.; Alpsoy, S.; Rifaioglu, E.N.; Kaya, S.; Kucukyalcın, V.; Topcu, B.; Kulac, M. Serum YKL-40: A potential biomarker for psoriasis or endothelial dysfunction in psoriasis? Mol. Cell. Biochem. 2015, 400, 207–212. [Google Scholar] [CrossRef]
- Hotamisligil, G.S.; Bernlohr, D.A. Metabolic functions of FABPs—Mechanisms and therapeutic implications. Nat. Rev. Endocrinol. 2015, 11, 592–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storch, J.; Thumser, A.E. Tissue-specific functions in the fatty acid-binding protein family. J. Biol. Chem. 2010, 285, 32679–32683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smathers, R.L.; Petersen, D.R. The human fatty acid-binding protein family: Evolutionary divergences and functions. Hum. Genom. 2011, 5, 170–191. [Google Scholar] [CrossRef]
- Watanabe, R.; Fujii, H.; Yamamoto, A.; Hashimoto, T.; Kameda, K.; Ito, M.; Ono, T. Immunohistochemical distribution of cutaneous fatty acid-binding protein in human skin. J. Dermatol. Sci. 1997, 16, 17–22. [Google Scholar] [CrossRef]
- Kuijpers, A.L.A.; Bergers, M.; Siegenthaler, G.; Zeeuwen, P.L.J.M.; Van De Kerkhof, P.C.M.; Schalkwijk, J. Skin-derived antileukoproteinase (SKALP) and epidermal fatty acid-binding protein (E-FABP): Two novel markers of the psoriatic phenotype that respond differentially to topical steroid. Acta Derm. Venereol. 1997, 77, 14–19. [Google Scholar] [PubMed]
- Madsen, P.; Rasmussen, H.H.; Leffers, H.; Honoré, B.; Celis, J.E. Molecular cloning and expression of a novel keratinocyte protein (psoriasis-associated fatty acid-binding protein [PA-FABP]) that is highly upregulated in psoriatic skin and that shares similarity to fatty acid-binding proteins. J. Investig. Dermatol. 1992, 99, 299–305. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, E.; Owada, Y.; Ikawa, S.; Adachi, Y.; Egawa, T.; Nemoto, K.; Suzuki, K.; Hishinuma, T.; Kawashima, H.; Kondo, H.; et al. Epidermal FABP (FABP5) regulates keratinocyte differentiation by 13(S)-HODE-mediated activation of the NF-B signaling pathway. J. Investig. Dermatol. 2011, 131, 604–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dallaglio, K.; Marconi, A.; Truzzi, F.; Lotti, R.; Palazzo, E.; Petrachi, T.; Saltari, A.; Coppini, M.; Pincelli, C. E-FABP induces differentiation in normal human keratinocytes and modulates the differentiation process in psoriatic keratinocytes in vitro. Exp. Dermatol. 2013, 22, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Myśliwiec, H.; Baran, A.; Harasim-Symbor, E.; Myśliwiec, P.; Milewska, A.J.; Chabowski, A.; Flisiak, I. Serum fatty acid profile in psoriasis and its comorbidity. Arch. Dermatol. Res. 2017, 309, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Baran, A.; Świderska, M.; Bacharewicz-Szczerbicka, J.; Myśliwiec, H.; Flisiak, I. Serum Fatty Acid-Binding Protein 4 is Increased in Patients with Psoriasis. Lipids 2017, 52, 51–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honma, M.; Shibuya, T.; Iinuma, S.; Ishida-Yamamoto, A. Serum fatty acid-binding protein 4 level is inversely correlated with serum thymus and activation-regulated chemokine level in psoriatic patients achieving clear skin by biologics. J. Dermatol. 2019, 46, e116–e117. [Google Scholar] [CrossRef]
- Shibuya, T.; Honma, M.; Iinuma, S.; Iwasaki, T.; Ishida-Yamamoto, A. Persistent pruritus in psoriatic patients during administration of biologics. J. Dermatol. 2018, 45, e223. [Google Scholar] [CrossRef]
- Baran, A.; Kiluk, P.; Maciaszek, M.; Świderska, M.; Flisiak, I. Liver fatty acid-binding protein might be a predictive marker of clinical response to systemic treatment in psoriasis. Arch. Dermatol. Res. 2019, 311, 389–397. [Google Scholar] [CrossRef] [Green Version]
- Sikora, M.; Stec, A.; Chrabaszcz, M.; Waskiel-Burnat, A.; Zaremba, M.; Olszewska, M.; Rudnicka, L. Intestinal Fatty Acid Binding Protein, a Biomarker of Intestinal Barrier, is Associated with Severity of Psoriasis. J. Clin. Med. 2019, 8, 1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Group | Biomarkers | Cellular Source | Findings |
|---|---|---|---|
| blood cell counts | NLR | - | increase especially in cases with arthritis |
| PLR | - | ||
| cytokines | IL-17A | Th17, Tc17, ILC3, etc. | relation with atherosclerosis, fatty liver, and insulin resistance |
| IL-17F | Th17, Tc17, ILC3, colon epithelial cells, etc. | much higher serum IL-17F levels than IL-17A levels | |
| IL-22 | Th17, Th22, Tc22, ILC3, etc. | vascular protective effect; relation with liver fibrosis | |
| IL-19 | monocytes, macrophages, keratinocytes, fibroblasts, etc. | vascular protective effect | |
| IL-36γ | epidermis | relatively specific to skin lesions | |
| chemokines | Fractalkine | APCs, ECs, and epidermis | close correlation with atherosclerosis |
| TARC | DCs, ECs, epidermis, and fibroblasts | a biomarker for AD; possible relation to deeper remission during anti-IL-17 therapy; correlation with severity of GPP | |
| adipokines | Resistin | macrophages, monocytes, and adipocytes | close correlation with atherosclerosis |
| Adiponectin | adipocytes | negatively correlated with atherosclerosis | |
| AMPs | β-defensin 2 | epidermis | relatively specific to skin lesion; |
| S100A7 | epidermis | correlation with atherosclerosis | |
| protease inhibitors | SCCA2 | epidermis | also increase in AD |
| Elafin | Epidermis and immune cells | correlation with CRP and ESR | |
| glycoproteins | LRG | hepatocytes, neutrophils, ECs, and macrophages | correlation with CRP and arthritis |
| YKL-40 | neutrophils, macrophages, fibroblasts, ECs, and smooth muscle cells | correlation with tumor progression, metabolic diseases, and arthritis | |
| FABPs | FABP-4 | adipocytes | increase in cardiovascular diseases; the expression in TRM infiltrating into psoriatic epidermis |
| i-FABP | intestine epithelial cells | correlation with disruption of intestine barrier |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Honma, M.; Nozaki, H. Molecular Pathogenesis of Psoriasis and Biomarkers Reflecting Disease Activity. J. Clin. Med. 2021, 10, 3199. https://doi.org/10.3390/jcm10153199
Honma M, Nozaki H. Molecular Pathogenesis of Psoriasis and Biomarkers Reflecting Disease Activity. Journal of Clinical Medicine. 2021; 10(15):3199. https://doi.org/10.3390/jcm10153199
Chicago/Turabian StyleHonma, Masaru, and Hiroyoshi Nozaki. 2021. "Molecular Pathogenesis of Psoriasis and Biomarkers Reflecting Disease Activity" Journal of Clinical Medicine 10, no. 15: 3199. https://doi.org/10.3390/jcm10153199
APA StyleHonma, M., & Nozaki, H. (2021). Molecular Pathogenesis of Psoriasis and Biomarkers Reflecting Disease Activity. Journal of Clinical Medicine, 10(15), 3199. https://doi.org/10.3390/jcm10153199