
Electronic Cigarette Aerosol Is Cytotoxic and Increases ACE2 Expression on Human Airway Epithelial Cells: Implications for SARS-CoV-2 (COVID-19)

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Electronic Cigarette Aerosol Condensate Production
2.2. Cell Culture
2.3. Cytotoxicity and Viability Measurements
2.4. qPCR
2.5. Immunofluorescence
2.6. Imaging
2.7. Statistics
3. Results
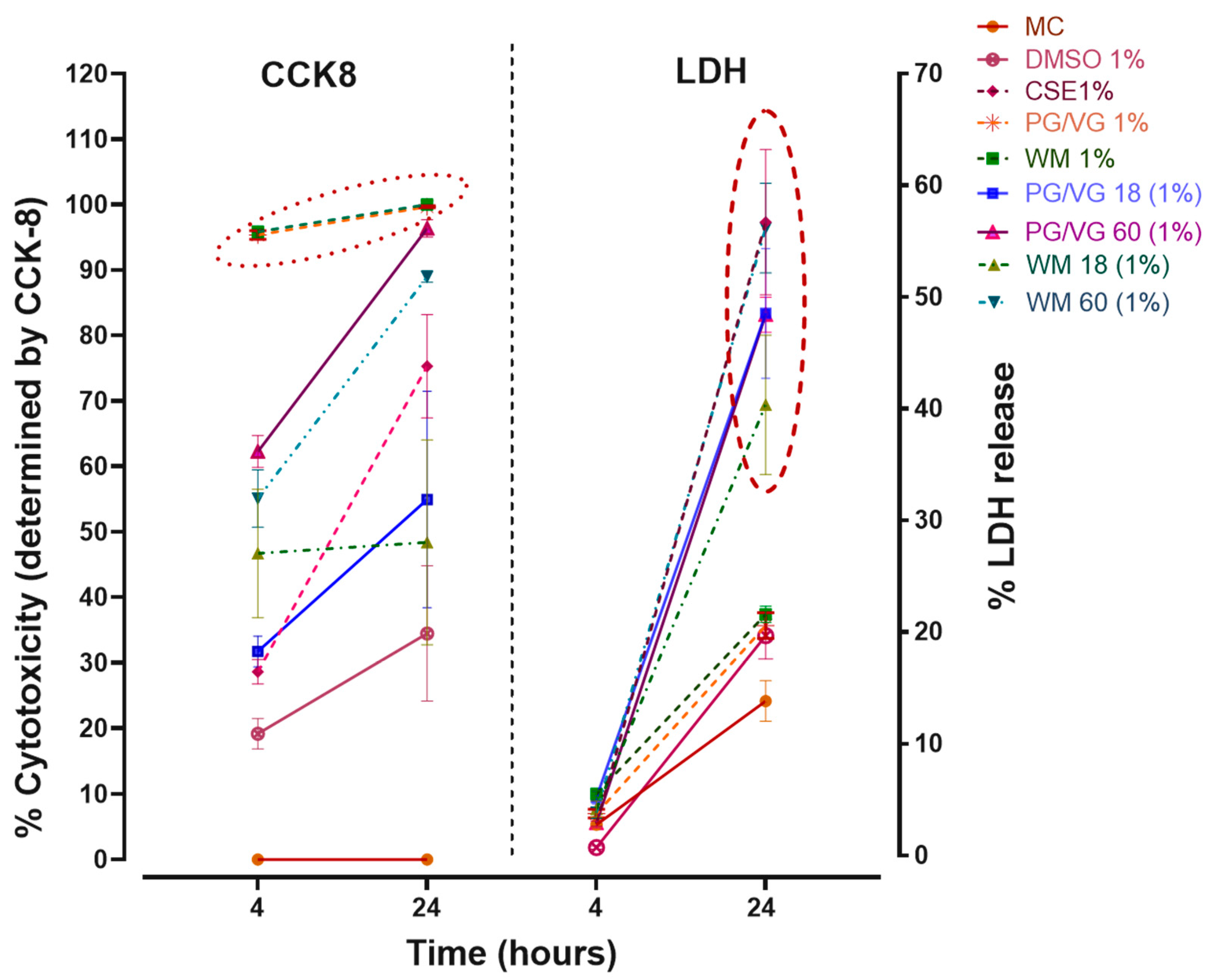
3.1. Exposure to Electronic Cigarette Aerosol Condensate and Cigarette Smoke Is Toxic to Bronchial Epithelial Cells (BEAS-2B)
3.2. Cellular Membrane Damage from e-Cigarette Aerosol Condensate and Cigarette Smoke Extract Are Distinct Following 24 h Exposure
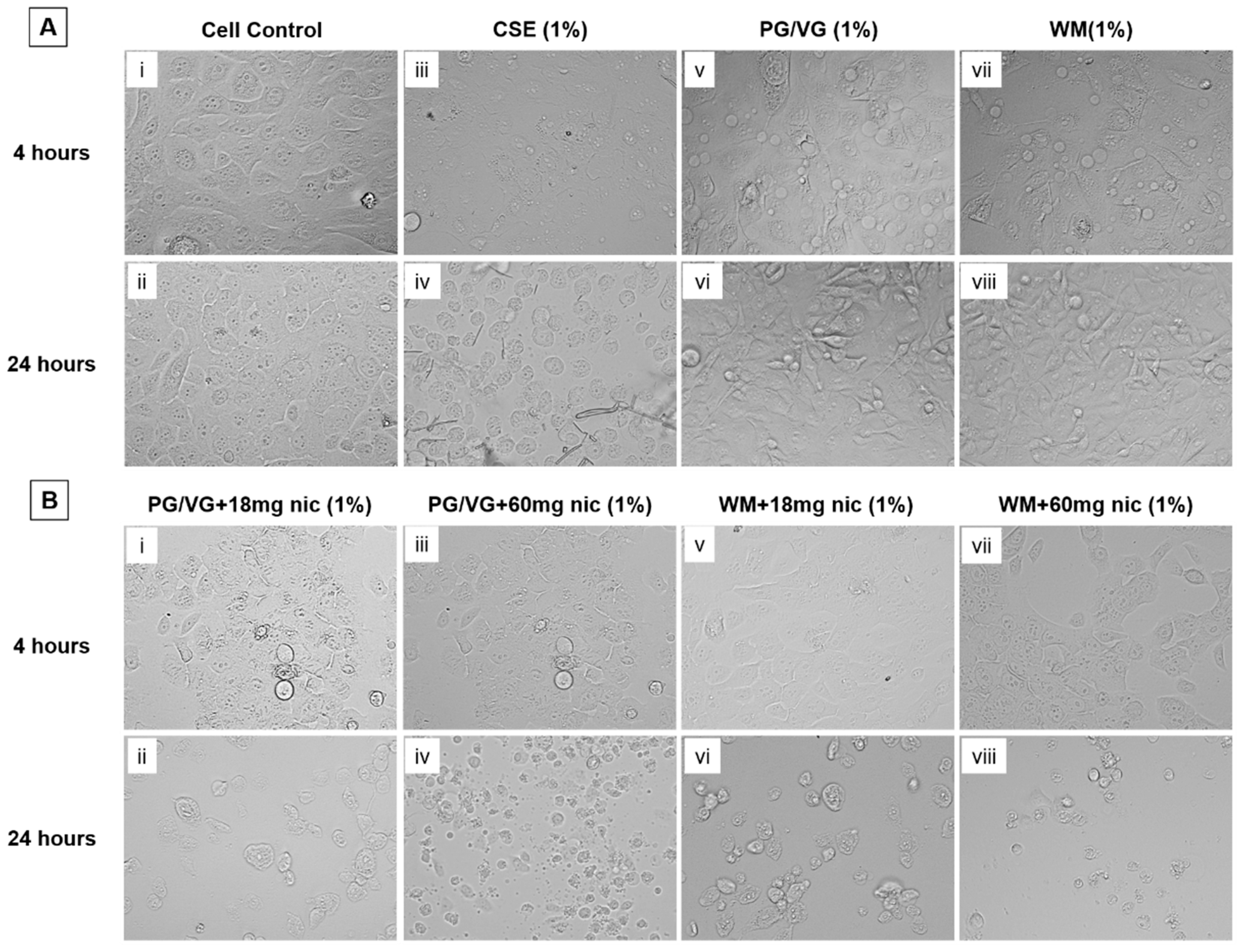
3.3. Notable Morphological Changes to BEAS-2B Cells Resulting from 24 h Exposure from e-Cigarette Aerosol Condensate and Cigarette Smoke Extract
3.4. Exposure to CSE Increases ACE2 Gene and Protein Expression in BEAS-2B Cells
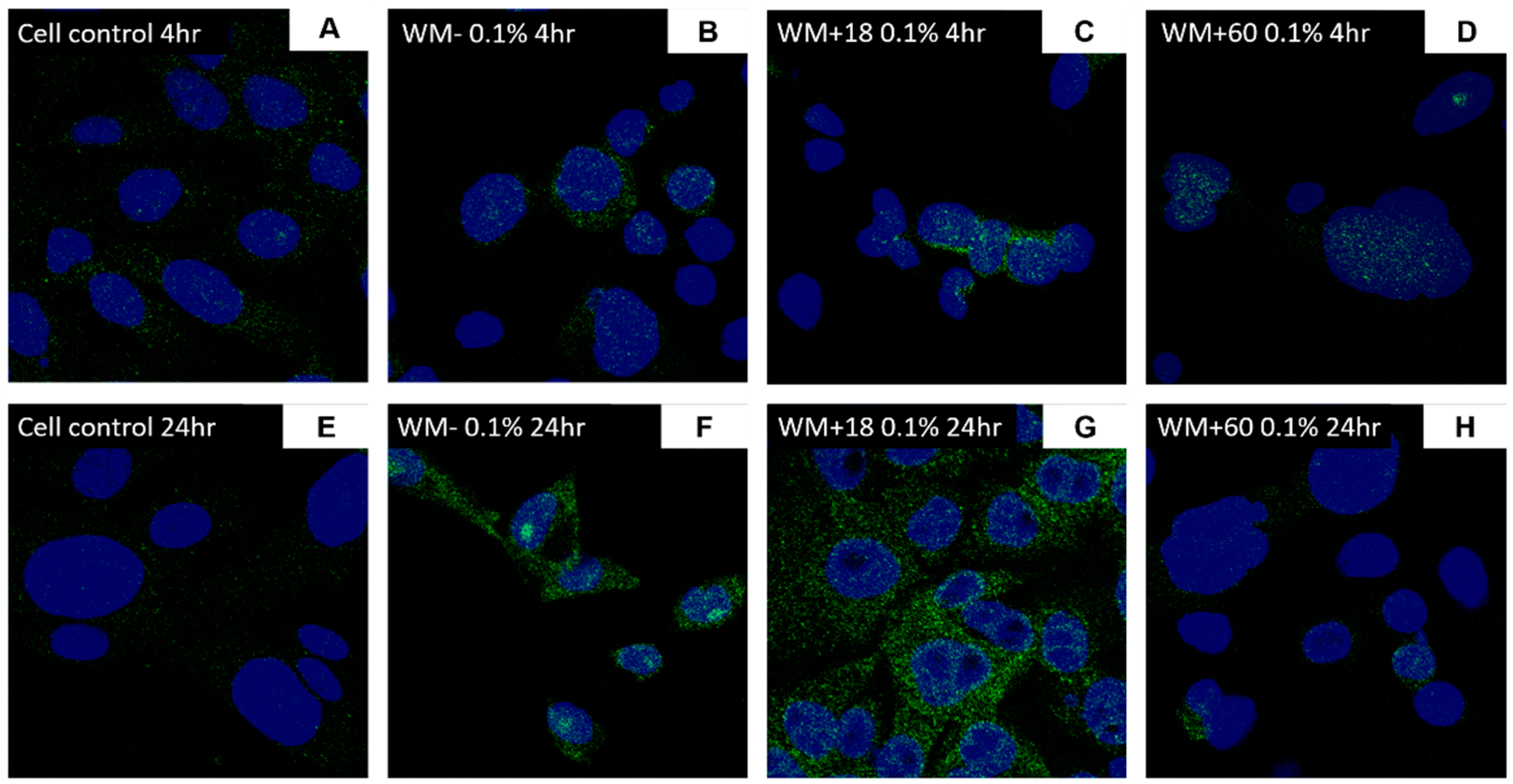
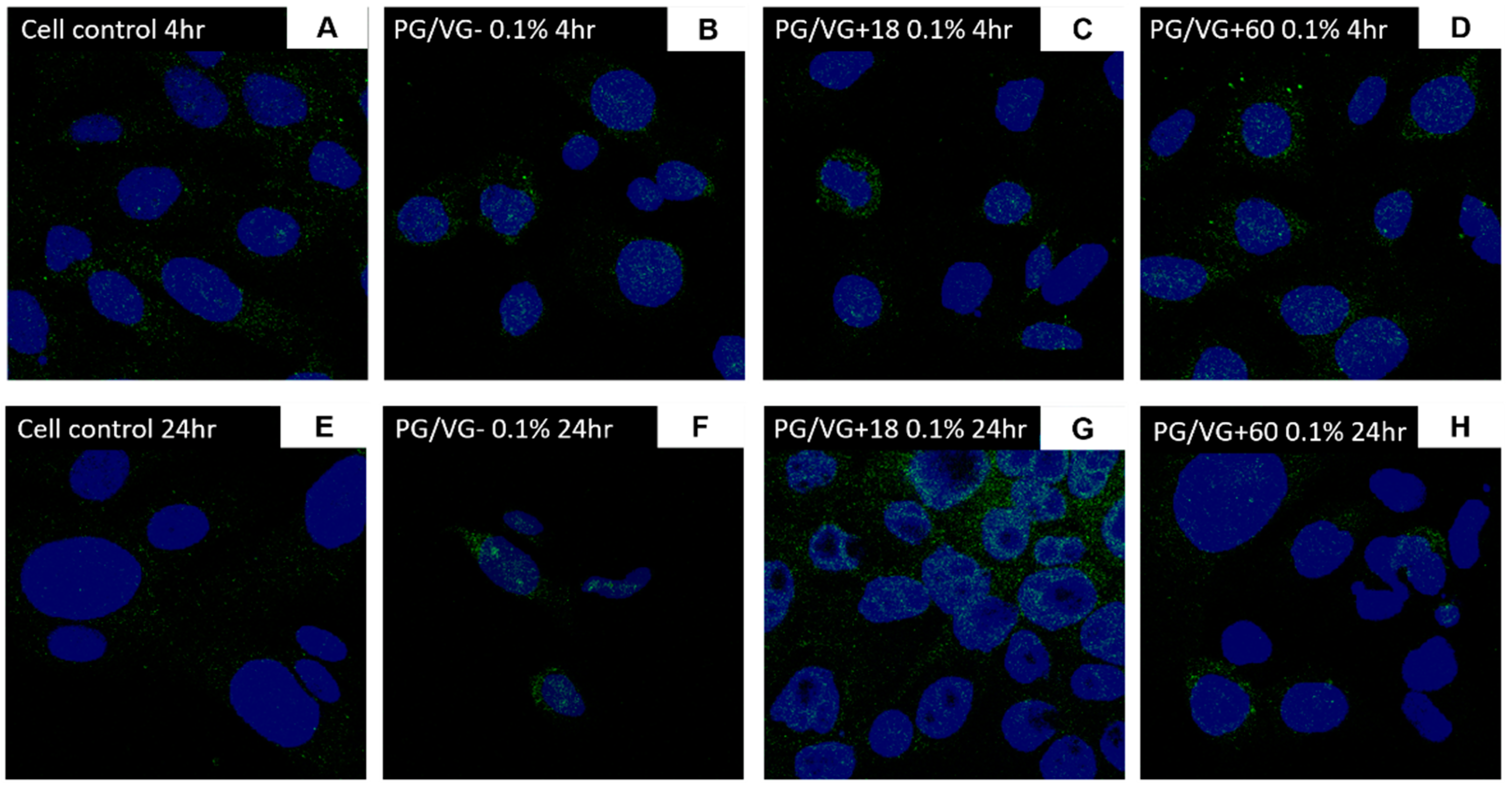
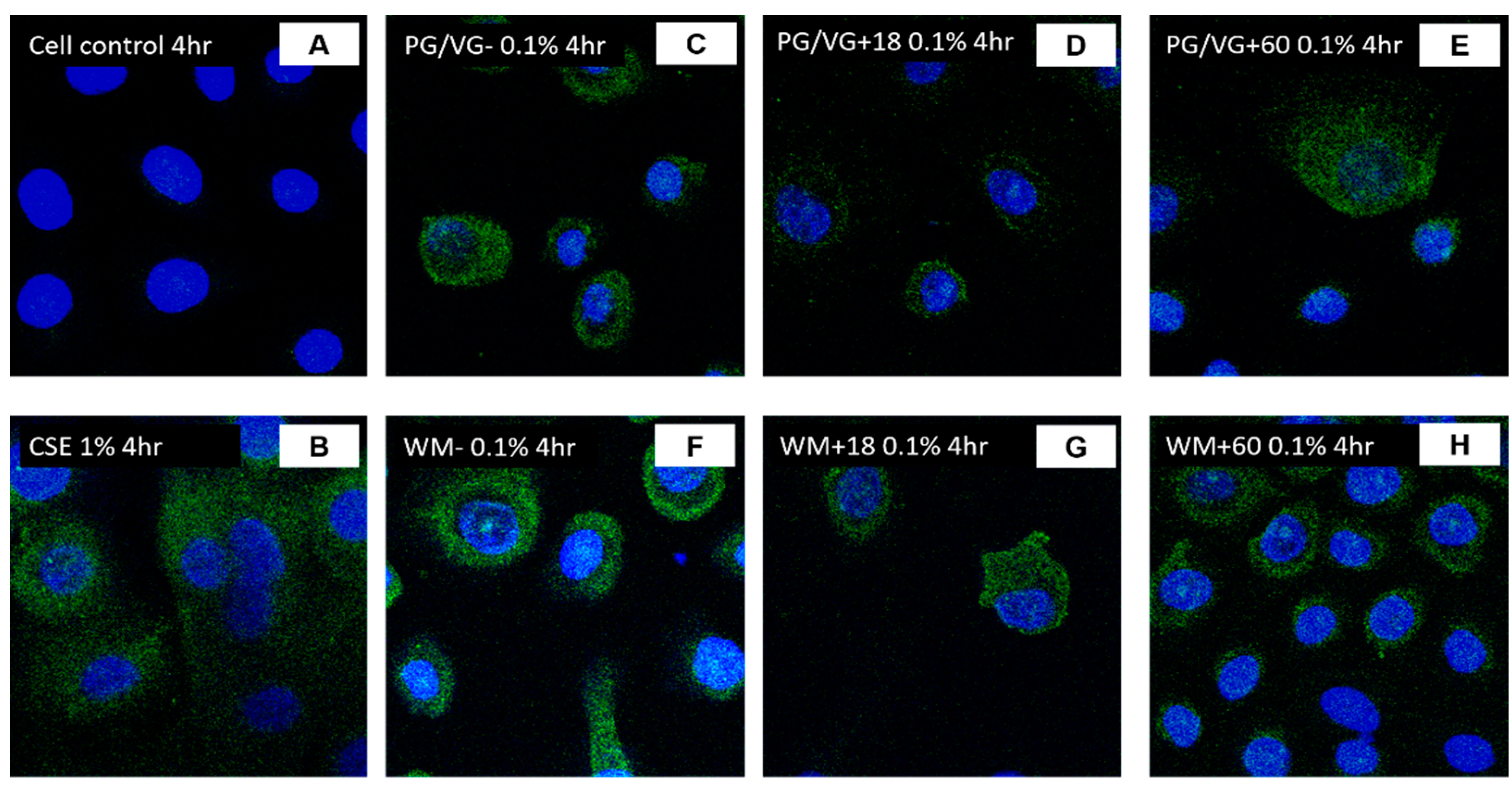
3.5. ACE2 Gene and Protein Expression Are Increased in BEAS-2B Cells Following Electronic Cigarette Aerosol Condensate Exposure
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Asrani, P.; Eapen, M.S.; Chia, C.; Haug, G.; Weber, H.C.; Hassan, I.; Sohal, S.S. Diagnostic approaches in COVID-19: Clinical updates. Expert Rev. Respir. Med. 2021, 15, 197–212. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Singh, A.; Ngasainao, M.R.; Shakeel, I.; Kumar, S.; Lal, S.; Singhal, A.; Sohal, S.; Singh, I.K.; Hassan, M. Potential diagnostics and therapeutic approaches in COVID-19. Clin. Chim. Acta 2020, 510, 488–497. [Google Scholar] [CrossRef]
- Asrani, P.; Hasan, G.M.; Sohal, S.S.; Hassan, I. Molecular Basis of Pathogenesis of Coronaviruses: A Comparative Genomics Approach to Planetary Health to Prevent Zoonotic Outbreaks in the 21st Century. OMICS A J. Integr. Biol. 2020, 24, 634–644. [Google Scholar] [CrossRef]
- World Health Organization. WHO Coronavirus Disease (COVID-19) Dashboard. 2020. Available online: https://covid19.who.int/ (accessed on 27 February 2021).
- National Institutes of Health, U.S. Department of Health and Human Services. Novel Coronavirus Structure Reveals Targets for Vaccines and Treatments. 2020. Available online: www.nih.gov/news-events/nih-research-matters/novel-coronavirus-structure-reveals-targets-vaccines-treatments (accessed on 20 November 2020).
- Asrani, P.; Hussain, A.; Nasreen, K.; AlAjmi, M.F.; Amir, S.; Sohal, S.S.; Hassan, M.I. Guidelines and Safety Considerations in the Laboratory Diagnosis of SARS-CoV-2 Infection: A Prerequisite Study for Health Professionals. Risk Manag. Healthc. Policy 2021, 14, 379–389. [Google Scholar] [CrossRef]
- Li, Q.; Guan, X.; Wu, P.; Wang, X.; Zhou, L.; Tong, Y.; Ren, R.; Leung, K.S.M.; Lau, E.H.Y.; Wong, J.Y.; et al. Early transmission dynamics in Wuhan, China, of Novel Coronavirus–infected pneumonia. N. Engl. J. Med. 2020, 382, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Sanche, S.; Lin, Y.T.; Xu, C.; Romero-Severson, E.; Hengartner, N.; Ke, R. High Contagiousness and Rapid Spread of Severe Acute Respiratory Syndrome Coronavirus 2. Emerg. Infect. Dis. 2020, 26, 1470–1477. [Google Scholar] [CrossRef]
- Tang, B.; Wang, X.; Li, Q.; Bragazzi, N.L.; Tang, S.; Xiao, Y.; Wu, J. Estimation of the Transmission Risk of the 2019-nCoV and Its Implication for Public Health Interventions. J. Clin. Med. 2020, 9, 462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Li, W.; Farzan, M.; Harrison, S.C. Structure of SARS Coronavirus Spike Receptor-Binding Domain Complexed with Receptor. Science 2005, 309, 1864–1868. [Google Scholar] [CrossRef]
- Coutard, B.; Valle, C.; De Lamballerie, X.; Canard, B.; Seidah, N.; Decroly, E. The spike glycoprotein of the new coronavirus 2019-nCoV contains a furin-like cleavage site absent in CoV of the same clade. Antivir. Res. 2020, 176, 104742. [Google Scholar] [CrossRef]
- Rice, G.I.; Thomas, D.A.; Grant, P.J.; Turner, A.J.; Hooper, N.M. Evaluation of angiotensin-converting enzyme (ACE), its homologue ACE2 and neprilysin in angiotensin peptide metabolism. Biochem. J. 2004, 383, 45–51. [Google Scholar] [CrossRef]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.-L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patanavanich, R.; Glantz, A.S. Smoking Is Associated With COVID-19 Progression: A Meta-analysis. Nicotine Tob. Res. 2020, 22, 1653–1656. [Google Scholar] [CrossRef] [PubMed]
- Polverino, F. Cigarette Smoking and COVID-19: A Complex Interaction. Am. J. Respir. Crit. Care Med. 2020, 202, 471–472. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Bayonas, A.; Jimenez-Fonseca, P.; Sánchez Arraez, Á.; Álvarez Manceñido, F.; Castañón, E. Does active smoking worsen Covid-19? Eur. J. Intern. Med. 2020, 77, 129–131. [Google Scholar] [CrossRef]
- Leung, J.M.; Sin, D.D. Smoking, ACE-2, and COVID-19: Ongoing Controversies. Eur. Respir. J. 2020, 56, 2001759. [Google Scholar] [CrossRef] [PubMed]
- Lukassen, S.; Chua, R.L.; Trefzer, T.; Kahn, N.C.; Schneider, M.A.; Muley, T.; Winter, H.; Meister, M.; Veith, C.; Boots, A.W.; et al. SARS -CoV-2 receptor ACE 2 and TMPRSS 2 are primarily expressed in bronchial transient secretory cells. EMBO J. 2020, 39, e105114. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.C.; Sausville, E.L.; Girish, V.; Yuan, M.L.; Vasudevan, A.; John, K.M.; Sheltzer, J.M. Cigarette Smoke Exposure and Inflammatory Signaling Increase the Expression of the SARS-CoV-2 Receptor ACE2 in the Respiratory Tract. Dev. Cell 2020, 53, 514–529.e513. [Google Scholar] [CrossRef]
- Zhang, H.; Rostami, M.R.; Leopold, P.L.; Mezey, J.G.; O’Beirne, S.L.; Strulovici-Barel, Y.; Crystal, R.G. Expression of the SARS-CoV-2 ACE2 Receptor in the Human Airway Epithelium. Am. J. Respir. Crit. Care Med. 2020, 202, 219–229. [Google Scholar] [CrossRef]
- Cai, G.; Bossé, Y.; Xiao, F.; Kheradmand, F.; Amos, C.I. Tobacco Smoking Increases the Lung Gene Expression of ACE2, the Receptor of SARS-CoV-2. Am. J. Respir. Crit. Care Med. 2020, 201, 1557–1559. [Google Scholar] [CrossRef]
- Brake, S.J.; Barnsley, K.; Lu, W.; McAlinden, K.D.; Eapen, M.S.; Sohal, S.S. Smoking Upregulates Angiotensin-Converting Enzyme-2 Receptor: A Potential Adhesion Site for Novel Coronavirus SARS-CoV-2 (Covid-19). J. Clin. Med. 2020, 9, 841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, J.M.; Yang, C.X.; Tam, A.; Shaipanich, T.; Hackett, T.-L.; Singhera, G.K.; Dorscheid, D.R.; Sin, D.D. ACE-2 expression in the small airway epithelia of smokers and COPD patients: Implications for COVID-19. Eur. Respir. J. 2020, 55, 2000688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, M.; Van Eeckhoutte, H.P.; Wijnant, S.R.A.; Janssens, W.; Joos, G.F.; Brusselle, G.G.; Bracke, K.R. Increased expression of ACE2, the SARS-CoV-2 entry receptor, in alveolar and bronchial epithelium of smokers and COPD subjects. Eur. Respir. J. 2020, 56, 2002378. [Google Scholar] [CrossRef] [PubMed]
- Eapen, M.S.; Lu, W.; Hackett, T.L.; Singhera, G.K.; Thompson, I.E.; McAlinden, K.D.; Hardikar, A.; Weber, H.C.; Haug, G.; Wark, P.A.B.; et al. Dysregulation of endocytic machinery and ACE2 in small airways of smokers and COPD patients can augment their susceptibility to SARS-CoV-2 (COVID-19) infections. Am. J. Physiol. Lung Cell. Mol. Physiol. 2020, 320, L158–L163. [Google Scholar] [CrossRef] [PubMed]
- Wark, P.A.B.; Pathinayake, P.S.; Kaiko, G.; Nichol, K.; Ali, A.; Chen, L.; Sutanto, E.N.; Garratt, L.W.; Sohal, S.S.; Lu, W.; et al. ACE2 expression is elevated in airway epithelial cells from older and male healthy individuals but reduced in asthma. Respirology 2021. [Google Scholar] [CrossRef]
- Russo, P.; Bonassi, S.; Giacconi, R.; Malavolta, M.; Tomino, C.; Maggi, F. COVID-19 and Smoking. Is Nicotine the Hidden Link? Eur. Respir. J. 2020, 55, 2001116. [Google Scholar] [CrossRef]
- Lee, A.C.; Chakladar, J.; Li, W.T.; Chen, C.; Chang, E.Y.; Wang-Rodriguez, J.; Ongkeko, W.M. Tobacco, but Not Nicotine and Flavor-Less Electronic Cigarettes, Induces ACE2 and Immune Dysregulation. Int. J. Mol. Sci. 2020, 21, 5513. [Google Scholar] [CrossRef] [PubMed]
- Gaiha, S.M.; Cheng, J.; Halpern-Felsher, B. Association Between Youth Smoking, Electronic Cigarette Use, and COVID-19. J. Adolesc. Health 2020, 67, 519–523. [Google Scholar] [CrossRef]
- McAlinden, K.D.; Eapen, M.S.; Lu, W.; Chia, C.; Haug, G.; Sohal, S.S. COVID-19 and vaping: Risk for increased susceptibility to SARS-CoV-2 infection? Eur. Respir. J. 2020, 56, 2001645. [Google Scholar] [CrossRef]
- McAlinden, K.D.; Eapen, M.S.; Lu, W.; Sharma, P.; Sohal, S.S. The rise of electronic nicotine delivery systems and the emergence of electronic-cigarette-driven disease. Am. J. Physiol. Cell. Mol. Physiol. 2020, 319, L585–L595. [Google Scholar] [CrossRef]
- Ji, M.; Zhang, Y.; Li, N.; Wang, C.; Xia, R.; Zhang, Z.; Wang, S.-L. Nicotine Component of Cigarette Smoke Extract (CSE) Decreases the Cytotoxicity of CSE in BEAS-2B Cells Stably Expressing Human Cytochrome P450 2A13. Int. J. Environ. Res. Public Health 2017, 14, 1221. [Google Scholar] [CrossRef] [Green Version]
- Haug, G.; Eapen, M.S.; Sohal, S.S. Renin-Angiotensin-Aldosterone System Inhibitors in Covid-19. N. Engl. J. Med. 2020, 382, 92. [Google Scholar] [CrossRef]
- Serpa, G.L.; Renton, N.D.; Lee, N.; Crane, M.J.; Jamieson, A.M. Electronic Nicotine Delivery System Aerosol-induced Cell Death and Dysfunction in Macrophages and Lung Epithelial Cells. Am. J. Respir. Cell Mol. Biol. 2020, 63, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Yu, V.; Rahimy, M.; Korrapati, A.; Xuan, Y.; Zou, A.E.; Krishnan, A.R.; Tsui, T.; Aguilera, J.A.; Advani, S.J.; Alexander, L.E.C.; et al. Electronic cigarettes induce DNA strand breaks and cell death independently of nicotine in cell lines. Oral Oncol. 2016, 52, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Kosmider, L.; Sobczak, A.; Fik, M.; Knysak, J.; Zaciera, M.; Kurek, J.; Goniewicz, M.L. Carbonyl Compounds in Electronic Cigarette Vapors: Effects of Nicotine Solvent and Battery Output Voltage. Nicotine Tob. Res. 2014, 16, 1319–1326. [Google Scholar] [CrossRef] [PubMed]
- Hartnett, K.P.; Kite-Powell, A.; Patel, M.T.; Haag, B.L.; Sheppard, M.J.; Dias, T.P.; King, B.A.; Melstrom, P.C.; Ritchey, M.D.; Stein, Z.; et al. Syndromic Surveillance for E-Cigarette, or Vaping, Product Use–Associated Lung Injury. N. Engl. J. Med. 2020, 382, 766–772. [Google Scholar] [CrossRef]
- Muthumalage, T.; Lucas, J.H.; Wang, Q.; Lamb, T.; McGraw, M.D.; Rahman, I. Pulmonary Toxicity and Inflammatory Response of E-Cigarette Vape Cartridges Containing Medium-Chain Triglycerides Oil and Vitamin E Acetate: Implications in the Pathogenesis of EVALI. Toxics 2020, 8, 46. [Google Scholar] [CrossRef]
- Wang, Q.; Sundar, I.K.; Li, D.; Lucas, J.H.; Muthumalage, T.; McDonough, S.R.; Rahman, I. E-cigarette-induced pulmonary inflammation and dysregulated repair are mediated by nAChR α7 receptor: Role of nAChR α7 in SARS-CoV-2 Covid-19 ACE2 receptor regulation. Respir. Res. 2020, 21, 154. [Google Scholar] [CrossRef]
- Ramamurthi, D.; Chau, C.; Jackler, R.K. Exploitation of the COVID-19 pandemic by e-cigarette marketers. Tob. Control. 2020. [Google Scholar] [CrossRef] [PubMed]
- Sohal, S.S.; Eapen, M.S.; Naidu, V.G.; Sharma, P. IQOS exposure impairs human airway cell homeostasis: Direct comparison with traditional cigarette and e-cigarette. ERJ Open Res. 2019, 5. [Google Scholar] [CrossRef] [Green Version]
- McAlinden, K.D.; Eapen, M.S.; Lu, W.; Sharma, P.; Sohal, S.S. The Ill Effects of IQOS on Airway Cells: Let’s Not Get Burned All Over Again. Am. J. Respir. Cell Mol. Biol. 2020, 63, 269–270. [Google Scholar] [CrossRef]
- Crotty Alexander, L.E.; Ware, L.B.; Calfee, C.S.; Callahan, S.J.; Eissenberg, T.; Farver, C.; Goniewicz, M.L.; Jaspers, I.; Kheradmand, F.; King, T.E.; et al. E-Cigarette or Vaping Product Use-associated Lung Injury: Developing a Research Agenda. An NIH Workshop Report. Am. J. Respir. Crit. Care Med. 2020, 202, 795–802. [Google Scholar] [CrossRef] [PubMed]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McAlinden, K.D.; Lu, W.; Ferdowsi, P.V.; Myers, S.; Markos, J.; Larby, J.; Chia, C.; Weber, H.C.; Haug, G.; Eapen, M.S.; et al. Electronic Cigarette Aerosol Is Cytotoxic and Increases ACE2 Expression on Human Airway Epithelial Cells: Implications for SARS-CoV-2 (COVID-19). J. Clin. Med. 2021, 10, 1028. https://doi.org/10.3390/jcm10051028
McAlinden KD, Lu W, Ferdowsi PV, Myers S, Markos J, Larby J, Chia C, Weber HC, Haug G, Eapen MS, et al. Electronic Cigarette Aerosol Is Cytotoxic and Increases ACE2 Expression on Human Airway Epithelial Cells: Implications for SARS-CoV-2 (COVID-19). Journal of Clinical Medicine. 2021; 10(5):1028. https://doi.org/10.3390/jcm10051028
Chicago/Turabian StyleMcAlinden, Kielan Darcy, Wenying Lu, Parisa Vahidi Ferdowsi, Stephen Myers, James Markos, Josie Larby, Collin Chia, Heinrich C. Weber, Greg Haug, Mathew Suji Eapen, and et al. 2021. "Electronic Cigarette Aerosol Is Cytotoxic and Increases ACE2 Expression on Human Airway Epithelial Cells: Implications for SARS-CoV-2 (COVID-19)" Journal of Clinical Medicine 10, no. 5: 1028. https://doi.org/10.3390/jcm10051028
APA StyleMcAlinden, K. D., Lu, W., Ferdowsi, P. V., Myers, S., Markos, J., Larby, J., Chia, C., Weber, H. C., Haug, G., Eapen, M. S., & Sohal, S. S. (2021). Electronic Cigarette Aerosol Is Cytotoxic and Increases ACE2 Expression on Human Airway Epithelial Cells: Implications for SARS-CoV-2 (COVID-19). Journal of Clinical Medicine, 10(5), 1028. https://doi.org/10.3390/jcm10051028