
Extracellular Vesicles Derived from Early and Late Stage Plasmodium falciparum-Infected Red Blood Cells Contain Invasion-Associated Proteins

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Parasite Culture
2.2. Extracellular Vesicle Isolation
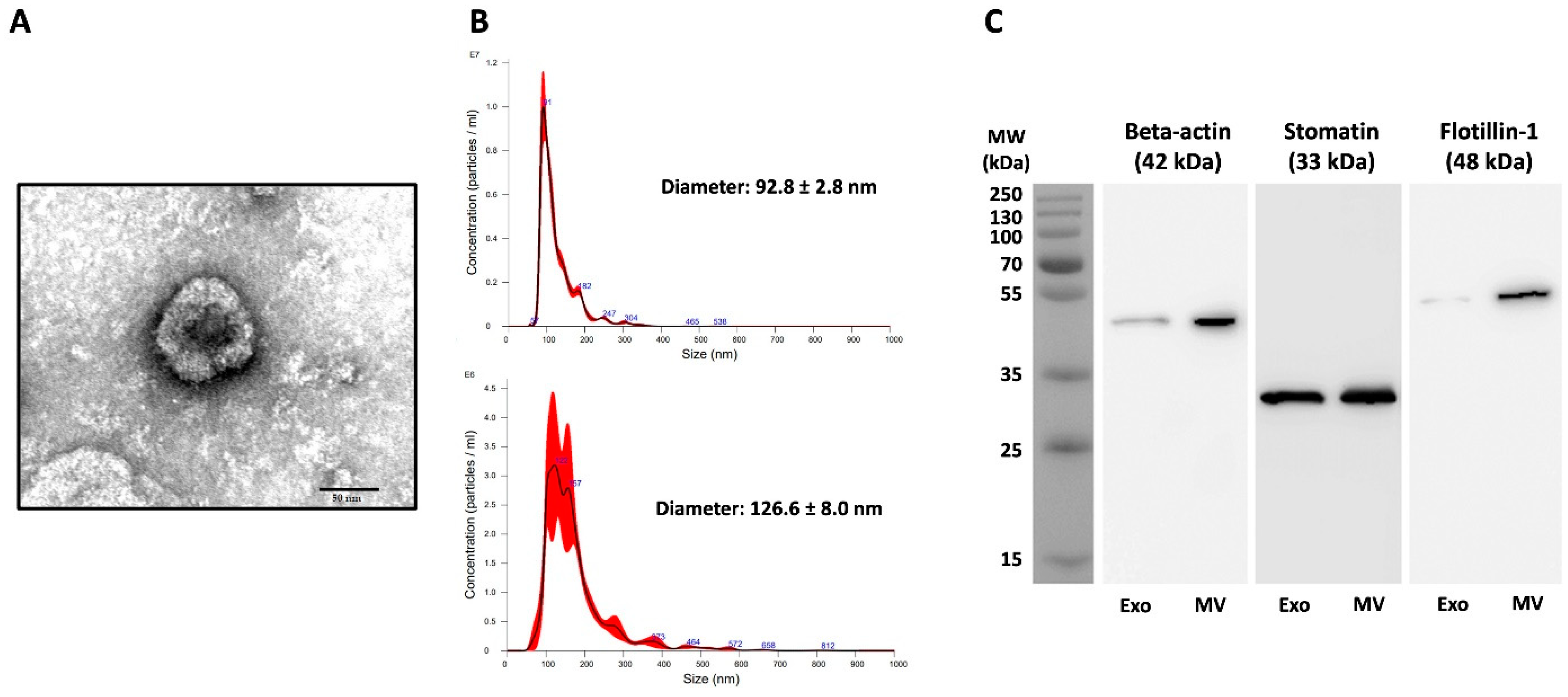
2.3. Transmission Electron Microscopy (TEM)
2.4. Nanoparticle Tracking Analysis (NTA)
2.5. Western Blotting Analysis
2.6. Exosomal Protein Preparation
2.7. Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS)
2.8. Bioinformatics and Data Analysis
2.9. Invasion and Growth Development Evaluation
2.10. Statistic Analysis
3. Results
3.1. Characterization of Pf-Infected RBC-Derived EVs
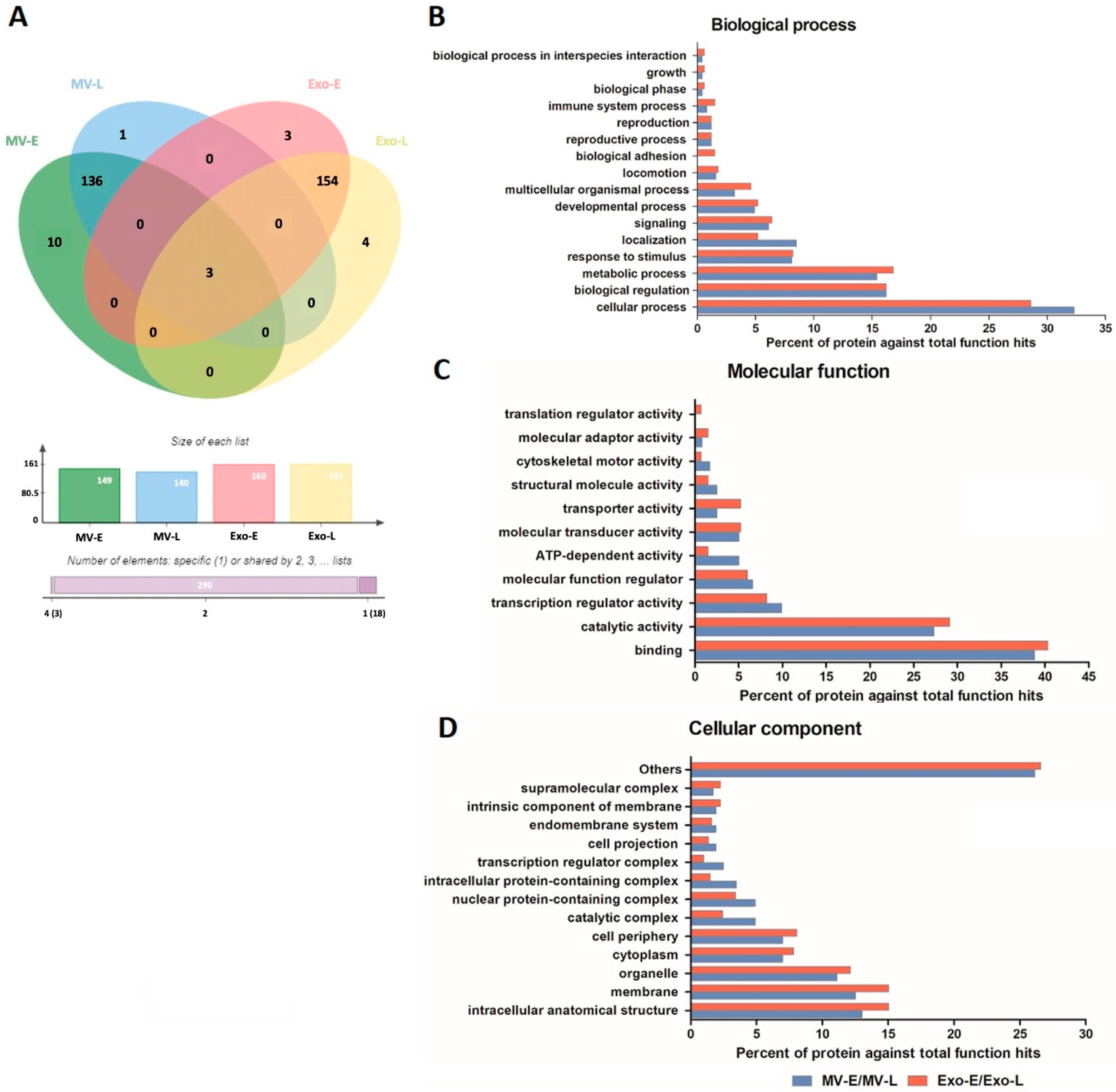
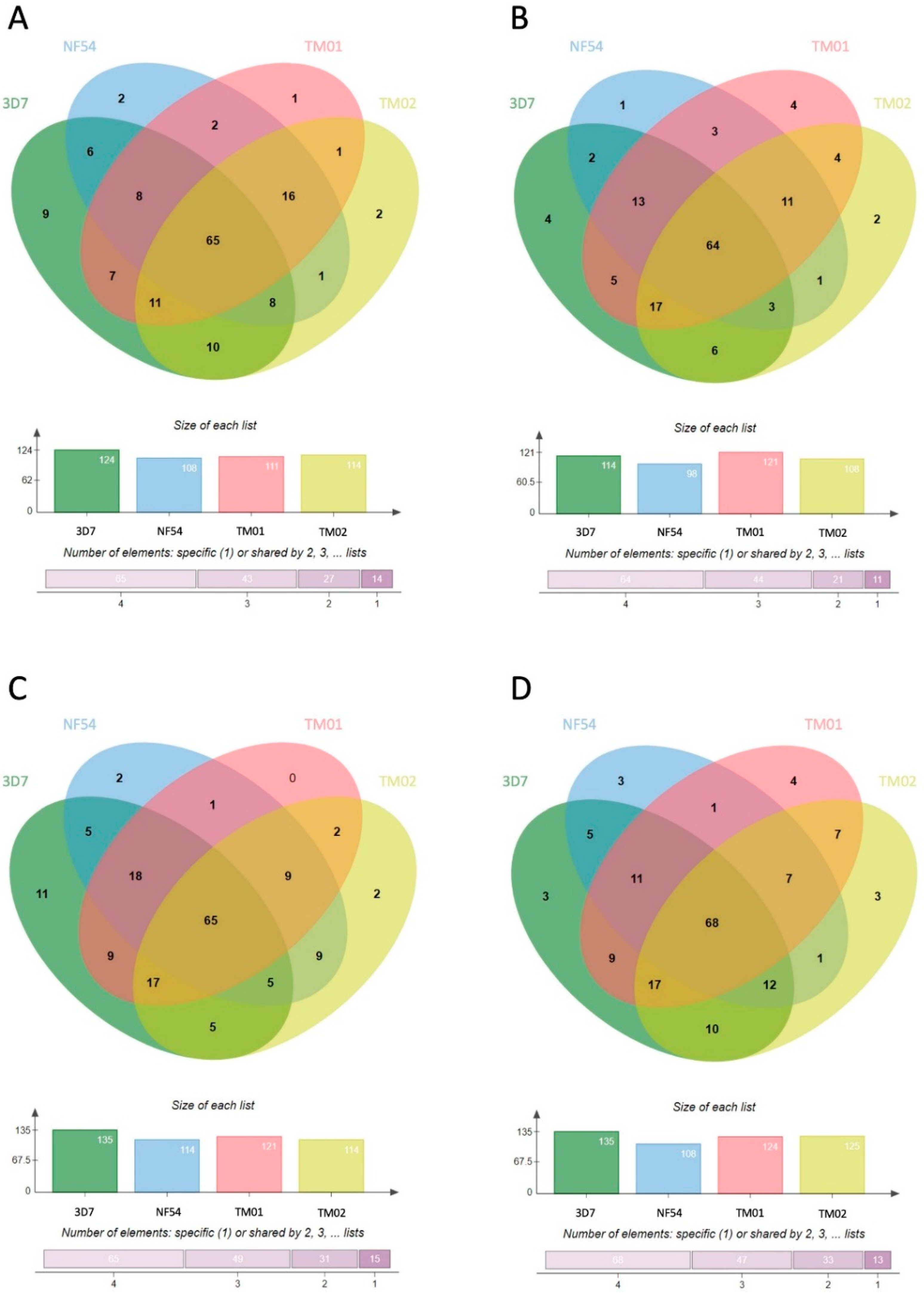
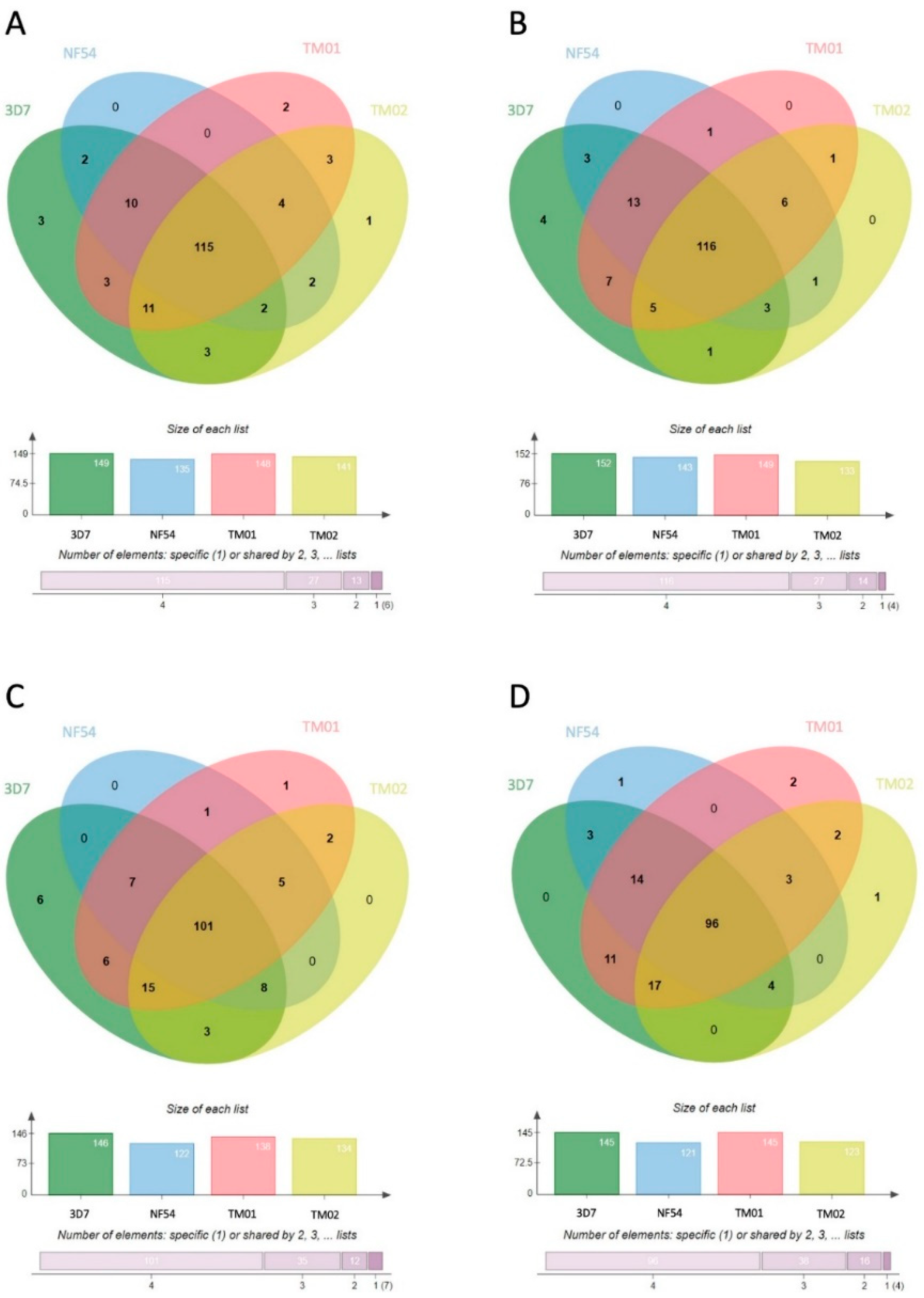
3.2. Characterization of the Human Proteome in P. falciparum-Infected RBC-Derived EVs
3.3. Characterization of the P. falciparum Proteome in Pf-EVs
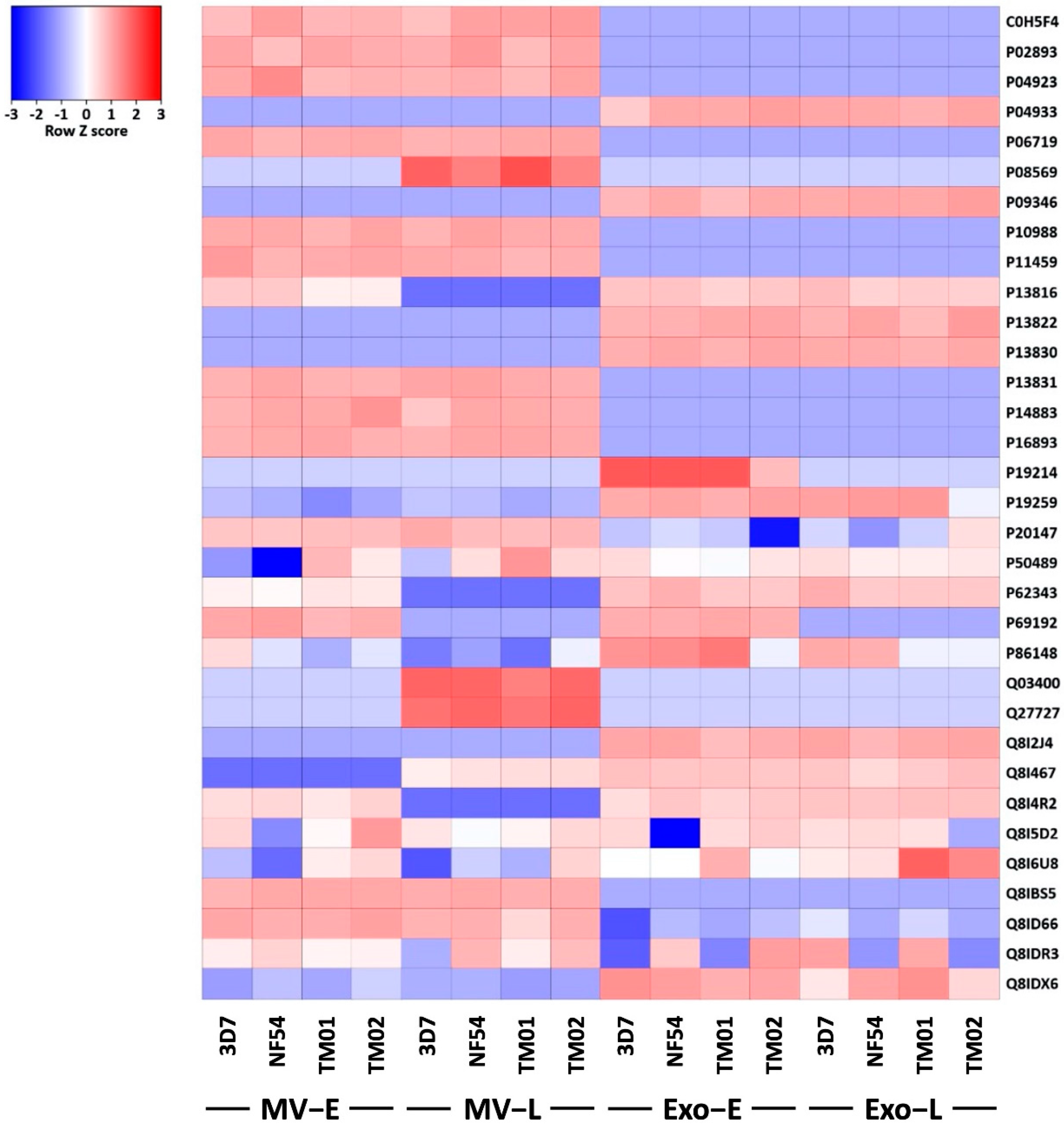
3.4. Pf-EVs Contain Parasite-Derived Proteins That Are Associated with Parasite Invasion
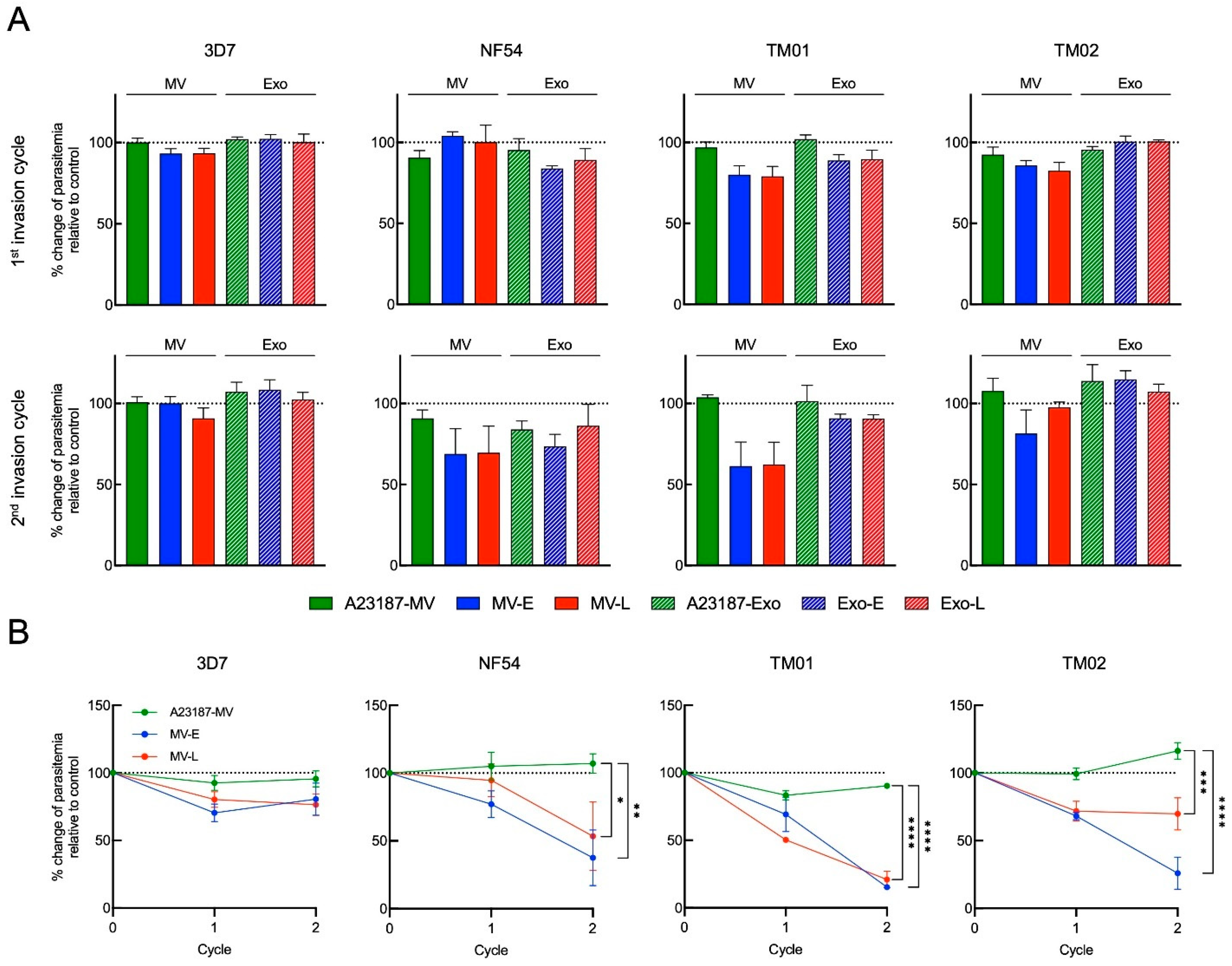
3.5. Pf-EVs in Invasion Efficiency and Growth Development
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. World Malaria Report 2021; WHO Press: Geneva, Switzerland, 2021. [Google Scholar]
- Volz, J.C.; Yap, A.; Sisquella, X.; Thompson, J.K.; Lim, N.T.; Whitehead, L.W.; Chen, L.; Lampe, M.; Tham, W.H.; Wilson, D.; et al. Essential role of the PfRh5/PfRipr/CyRPA complex during Plasmodium falciparum invasion of erythrocytes. Cell Host Microbe 2016, 20, 60–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, K.; Mayer, D.C.; Singh, S.; Reid, M.; Miller, L.H. Domain III of Plasmodium falciparum apical membrane antigen 1 binds to the erythrocyte membrane protein Kx. Proc. Natl. Acad. Sci. USA 2005, 102, 5552–5557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treeck, M.; Zacherl, S.; Herrmann, S.; Cabrera, A.; Kono, M.; Struck, N.S.; Engelberg, K.; Haase, S.; Frischknecht, F.; Miura, K.; et al. Functional analysis of the leading malaria vaccine candidate AMA-1 reveals an essential role for the cytoplasmic domain in the invasion process. PLoS Pathog. 2009, 5, e1000322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Chu, R.; Yin, Y.; Yu, H.; Xu, Q.; Yang, B.; Sun, Y.; Song, J.; Wang, Q.; Xu, J.; et al. Glycosylphosphatidylinositol-anchored micronemal antigen (GAMA) interacts with the band 3 receptor to promote erythrocyte invasion by malaria parasites. J. Biol. Chem. 2022, 298, 101765. [Google Scholar] [CrossRef] [PubMed]
- Schlott, A.C.; Knuepfer, E.; Green, J.L.; Hobson, P.; Borg, A.J.; Morales-Sanfrutos, J.; Perrin, A.J.; Maclachlan, C.; Collinson, L.M.; Snijders, A.P.; et al. Inhibition of protein N-myristoylation blocks Plasmodium falciparum intraerythrocytic development, egress and invasion. PLoS Biol. 2021, 19, e3001408. [Google Scholar] [CrossRef]
- Robert-Paganin, J.; Robblee, J.P.; Auguin, D.; Blake, T.C.A.; Bookwalter, C.S.; Krementsova, E.B.; Moussaoui, D.; Previs, M.J.; Jousset, G.; Baum, J.; et al. Plasmodium myosin A drives parasite invasion by an atypical force generating mechanism. Nat. Commun. 2019, 10, 3286. [Google Scholar] [CrossRef]
- Patarroyo, M.A.; Molina-Franky, J.; Gomez, M.; Arevalo-Pinzon, G.; Patarroyo, M.E. Hotspots in Plasmodium and RBC Receptor-Ligand Interactions: Key Pieces for Inhibiting Malarial Parasite Invasion. Int. J. Mol. Sci. 2020, 21, 4729. [Google Scholar] [CrossRef]
- Cowman, A.F.; Tonkin, C.J.; Tham, W.H.; Duraisingh, M.T. The Molecular basis of erythrocyte invasion by malaria parasites. Cell Host Microbe 2017, 22, 232–245. [Google Scholar] [CrossRef]
- Lopaticki, S.; Maier, A.G.; Thompson, J.; Wilson, D.W.; Tham, W.H.; Triglia, T.; Gout, A.; Speed, T.P.; Beeson, J.G.; Healer, J.; et al. Reticulocyte and erythrocyte binding-like proteins function cooperatively in invasion of human erythrocytes by malaria parasites. Infect. Immun. 2011, 79, 1107–1117. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.R.; Douglas, A.D.; Miura, K.; Illingworth, J.J.; Choudhary, P.; Murungi, L.M.; Furze, J.M.; Diouf, A.; Miotto, O.; Crosnier, C.; et al. Enhancing blockade of Plasmodium falciparum erythrocyte invasion: Assessing combinations of antibodies against PfRH5 and other merozoite antigens. PLoS Pathog. 2012, 8, e1002991. [Google Scholar] [CrossRef] [Green Version]
- Nantakomol, D.; Dondorp, A.M.; Krudsood, S.; Udomsangpetch, R.; Pattanapanyasat, K.; Combes, V.; Grau, G.E.; White, N.J.; Viriyavejakul, P.; Day, N.P.; et al. Circulating red cell-derived microparticles in human malaria. J. Infect. Dis. 2011, 203, 700–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slomka, A.; Urban, S.K.; Lukacs-Kornek, V.; Zekanowska, E.; Kornek, M. Large extracellular vesicles: Have we found the holy grail of inflammation? Front. Immunol. 2018, 9, 2723. [Google Scholar] [CrossRef]
- Hezel, M.E.V.; Nieuwland, R.; Bruggen, R.V.; Juffermans, N.P. The Ability of Extracellular vesicles to induce a pro-inflammatory host response. Int. J. Mol. Sci. 2017, 18, 1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robbins, P.D.; Morelli, A.E. Regulation of immune responses by extracellular vesicles. Nat. Rev. Immunol. 2014, 14, 195–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Xie, F.; Wang, L.; Zhang, L.; Zhang, S.; Fang, M.; Zhou, F. The function and clinical application of extracellular vesicles in innate immune regulation. Cell Mol. Immunol. 2020, 17, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Maas, S.L.N.; Breakefield, X.O.; Weaver, A.M. Extracellular vesicles: Unique intercellular delivery vehicles. Trends Cell Biol. 2017, 27, 172–188. [Google Scholar] [CrossRef] [Green Version]
- Julich, H.; Willms, A.; Lukacs-Kornek, V.; Kornek, M. Extracellular vesicle profiling and their use as potential disease specific biomarker. Front. Immunol. 2014, 5, 413. [Google Scholar] [CrossRef] [Green Version]
- El-Assaad, F.; Wheway, J.; Hunt, N.H.; Grau, G.E.; Combes, V. Production, fate and pathogenicity of plasma microparticles in murine cerebral malaria. PLoS Pathog. 2014, 10, e1003839. [Google Scholar] [CrossRef] [Green Version]
- Couper, K.N.; Barnes, T.; Hafalla, J.C.; Combes, V.; Ryffel, B.; Secher, T.; Grau, G.E.; Riley, E.M.; de Souza, J.B. Parasite-derived plasma microparticles contribute significantly to malaria infection-induced inflammation through potent macrophage stimulation. PLoS Pathog. 2010, 6, e1000744. [Google Scholar] [CrossRef] [Green Version]
- Combes, V.; Coltel, N.; Alibert, M.; van Eck, M.; Raymond, C.; Juhan-Vague, I.; Grau, G.E.; Chimini, G. ABCA1 gene deletion protects against cerebral malaria: Potential pathogenic role of microparticles in neuropathology. Am. J. Pathol. 2005, 166, 295–302. [Google Scholar] [CrossRef]
- Penet, M.F.; Abou-Hamdan, M.; Coltel, N.; Cornille, E.; Grau, G.E.; de Reggi, M.; Gharib, B. Protection against cerebral malaria by the low-molecular-weight thiol pantethine. Proc. Natl. Acad. Sci. USA 2008, 105, 1321–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, F.M.; Franklin, B.S.; Teixeira-Carvalho, A.; Filho, A.L.; de Paula, S.C.; Fontes, C.J.; Brito, C.F.; Carvalho, L.H. Augmented plasma microparticles during acute Plasmodium vivax infection. Malar. J. 2010, 9, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pankoui Mfonkeu, J.B.; Gouado, I.; Fotso Kuate, H.; Zambou, O.; Amvam Zollo, P.H.; Grau, G.E.; Combes, V. Elevated cell-specific microparticles are a biological marker for cerebral dysfunctions in human severe malaria. PLoS ONE 2010, 5, e13415. [Google Scholar] [CrossRef] [PubMed]
- Mantel, P.Y.; Hoang, A.N.; Goldowitz, I.; Potashnikova, D.; Hamza, B.; Vorobjev, I.; Ghiran, I.; Toner, M.; Irimia, D.; Ivanov, A.R.; et al. Malaria-infected erythrocyte-derived microvesicles mediate cellular communication within the parasite population and with the host immune system. Cell Host Microbe 2013, 13, 521–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sisquella, X.; Ofir-Birin, Y.; Pimentel, M.A.; Cheng, L.; Abou Karam, P.; Sampaio, N.G.; Penington, J.S.; Connolly, D.; Giladi, T.; Scicluna, B.J.; et al. Malaria parasite DNA-harbouring vesicles activate cytosolic immune sensors. Nat. Commun. 2017, 8, 1985. [Google Scholar] [CrossRef]
- Sampaio, N.G.; Emery, S.J.; Garnham, A.L.; Tan, Q.Y.; Sisquella, X.; Pimentel, M.A.; Jex, A.R.; Regev-Rudzki, N.; Schofield, L.; Eriksson, E.M. Extracellular vesicles from early stage Plasmodium falciparum-infected red blood cells contain PfEMP1 and induce transcriptional changes in human monocytes. Cell Microbiol. 2018, 20, e12822. [Google Scholar] [CrossRef]
- Mantel, P.Y.; Hjelmqvist, D.; Walch, M.; Kharoubi-Hess, S.; Nilsson, S.; Ravel, D.; Ribeiro, M.; Gruring, C.; Ma, S.; Padmanabhan, P.; et al. Infected erythrocyte-derived extracellular vesicles alter vascular function via regulatory Ago2-miRNA complexes in malaria. Nat. Commun. 2016, 7, 12727. [Google Scholar] [CrossRef] [Green Version]
- Regev-Rudzki, N.; Wilson, D.W.; Carvalho, T.G.; Sisquella, X.; Coleman, B.M.; Rug, M.; Bursac, D.; Angrisano, F.; Gee, M.; Hill, A.F.; et al. Cell-cell communication between malaria-infected red blood cells via exosome-like vesicles. Cell 2013, 153, 1120–1133. [Google Scholar] [CrossRef] [Green Version]
- Correa, R.; Coronado, L.; Caballero, Z.; Faral-Tello, P.; Robello, C.; Spadafora, C. Extracellular vesicles carrying lactate dehydrogenase induce suicide in increased population density of Plasmodium falciparum in vitro. Sci. Rep. 2019, 9, 5042. [Google Scholar] [CrossRef]
- Pena, R.T.; Blasco, L.; Ambroa, A.; Gonzalez-Pedrajo, B.; Fernandez-Garcia, L.; Lopez, M.; Bleriot, I.; Bou, G.; Garcia-Contreras, R.; Wood, T.K.; et al. Relationship between quorum sensing and secretion systems. Front. Microbiol. 2019, 10, 1100. [Google Scholar] [CrossRef] [Green Version]
- Briggs, E.M.; Rojas, F.; McCulloch, R.; Matthews, K.R.; Otto, T.D. Single-cell transcriptomic analysis of bloodstream Trypanosoma brucei reconstructs cell cycle progression and developmental quorum sensing. Nat. Commun. 2021, 12, 5268. [Google Scholar] [CrossRef] [PubMed]
- Tettey, M.D.; Rojas, F.; Matthews, K.R. Extracellular release of two peptidases dominates generation of the trypanosome quorum-sensing signal. Nat. Commun. 2022, 13, 3322. [Google Scholar] [CrossRef] [PubMed]
- Gualdron-Lopez, M.; Flannery, E.L.; Kangwanrangsan, N.; Chuenchob, V.; Fernandez-Orth, D.; Segui-Barber, J.; Royo, F.; Falcon-Perez, J.M.; Fernandez-Becerra, C.; Lacerda, M.V.G.; et al. Characterization of Plasmodium vivax proteins in plasma-derived exosomes from malaria-infected liver-chimeric humanized mice. Front. Microbiol. 2018, 9, 1271. [Google Scholar] [CrossRef]
- Aparici-Herraiz, I.; Gualdron-Lopez, M.; Castro-Cavadia, C.J.; Carmona-Fonseca, J.; Yasnot, M.F.; Fernandez-Becerra, C.; Del Portillo, H.A. Antigen discovery in circulating extracellular vesicles from Plasmodium vivax patients. Front. Cell Infect. Microbiol. 2021, 11, 811390. [Google Scholar] [CrossRef]
- Salzer, U.; Hinterdorfer, P.; Hunger, U.; Borken, C.; Prohaska, R. Ca(++)-dependent vesicle release from erythrocytes involves stomatin-specific lipid rafts, synexin (annexin VII), and sorcin. Blood J. Am. Soc. Hematol. 2002, 99, 2569–2577. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Howe, E.A.; Sinha, R.; Schlauch, D.; Quackenbush, J. RNA-Seq analysis in MeV. Bioinformatics 2011, 27, 3209–3210. [Google Scholar] [CrossRef] [Green Version]
- Bardou, P.; Mariette, J.; Escudie, F.; Djemiel, C.; Klopp, C. jvenn: An interactive Venn diagram viewer. BMC Bioinform. 2014, 15, 293. [Google Scholar] [CrossRef] [Green Version]
- Mi, H.; Muruganujan, A.; Ebert, D.; Huang, X.; Thomas, P.D. PANTHER version 14: More genomes, a new PANTHER GO-slim and improvements in enrichment analysis tools. Nucleic Acids Res. 2019, 47, D419–D426. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Santos, A.; von Mering, C.; Jensen, L.J.; Bork, P.; Kuhn, M. STITCH 5: Augmenting protein-chemical interaction networks with tissue and affinity data. Nucleic Acids Res. 2016, 44, D380–D384. [Google Scholar] [CrossRef]
- Fonseka, P.; Pathan, M.; Chitti, S.V.; Kang, T.; Mathivanan, S. FunRich enables enrichment analysis of OMICs datasets. J. Mol. Biol. 2021, 433, 166747. [Google Scholar] [CrossRef] [PubMed]
- Rivadeneira, E.M.; Wasserman, M.; Espinal, C.T. Separation and concentration of schizonts of Plasmodium falciparum by Percoll gradients. J. Protozool. 1983, 30, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Vimonpatranon, S.; Chotivanich, K.; Sukapirom, K.; Lertjuthaporn, S.; Khowawisetsut, L.; Pattanapanyasat, K. Enumeration of the invasion efficiency of Plasmodium falciparum in vitro in four different red blood cell populations using a three-color flow cytometry-based method. Cytom. Part A 2019, 95, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Bryk, A.H.; Wisniewski, J.R. Quantitative Analysis of Human Red Blood Cell Proteome. J. Proteome Res. 2017, 16, 2752–2761. [Google Scholar] [CrossRef] [Green Version]
- Ravenhill, B.J.; Kanjee, U.; Ahouidi, A.; Nobre, L.; Williamson, J.; Goldberg, J.M.; Antrobus, R.; Dieye, T.; Duraisingh, M.T.; Weekes, M.P. Quantitative comparative analysis of human erythrocyte surface proteins between individuals from two genetically distinct populations. Commun. Biol. 2019, 2, 350. [Google Scholar] [CrossRef]
- Keerthikumar, S.; Chisanga, D.; Ariyaratne, D.; Al Saffar, H.; Anand, S.; Zhao, K.; Samuel, M.; Pathan, M.; Jois, M.; Chilamkurti, N.; et al. ExoCarta: A Web-Based Compendium of Exosomal Cargo. J. Mol. Biol. 2016, 428, 688–692. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, M.; von Mering, C.; Campillos, M.; Jensen, L.J.; Bork, P. STITCH: Interaction networks of chemicals and proteins. Nucleic Acids Res. 2008, 36, D684–D688. [Google Scholar] [CrossRef]
- Dekel, E.; Yaffe, D.; Rosenhek-Goldian, I.; Ben-Nissan, G.; Ofir-Birin, Y.; Morandi, M.I.; Ziv, T.; Sisquella, X.; Pimentel, M.A.; Nebl, T.; et al. 20S proteasomes secreted by the malaria parasite promote its growth. Nat. Commun. 2021, 12, 1172. [Google Scholar] [CrossRef]
- Yoon, Y.J.; Kim, O.Y.; Gho, Y.S. Extracellular vesicles as emerging intercellular communicasomes. BMB Rep. 2014, 47, 531–539. [Google Scholar] [CrossRef]
- Opadokun, T.; Rohrbach, P. Extracellular vesicles in malaria: An agglomeration of two decades of research. Malar. J. 2021, 20, 442. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, N.G.; Cheng, L.; Eriksson, E.M. The role of extracellular vesicles in malaria biology and pathogenesis. Malar. J. 2017, 16, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antwi-Baffour, S.; Adjei, J.K.; Agyemang-Yeboah, F.; Annani-Akollor, M.; Kyeremeh, R.; Asare, G.A.; Gyan, B. Proteomic analysis of microparticles isolated from malaria positive blood samples. Proteome Sci. 2016, 15, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Jaular, L.; Nakayasu, E.S.; Ferrer, M.; Almeida, I.C.; Del Portillo, H.A. Exosomes from Plasmodium yoelii-infected reticulocytes protect mice from lethal infections. PLoS ONE 2011, 6, e26588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdi, A.; Yu, L.; Goulding, D.; Rono, M.K.; Bejon, P.; Choudhary, J.; Rayner, J. Proteomic analysis of extracellular vesicles from a Plasmodium falciparum Kenyan clinical isolate defines a core parasite secretome. Wellcome Open Res. 2017, 2, 50. [Google Scholar] [CrossRef] [Green Version]
- Ponnudurai, T.; Leeuwenberg, A.D.; Meuwissen, J.H. Chloroquine sensitivity of isolates of Plasmodium falciparum adapted to in vitro culture. Trop. Geogr. Med. 1981, 33, 50–54. [Google Scholar]
- Bryant, J.M.; Baumgarten, S.; Lorthiois, A.; Scheidig-Benatar, C.; Claes, A.; Scherf, A. De Novo Genome Assembly of a Plasmodium falciparum NF54 clone using single-molecule real-time sequencing. Genome Announc. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Delves, M.J.; Straschil, U.; Ruecker, A.; Miguel-Blanco, C.; Marques, S.; Dufour, A.C.; Baum, J.; Sinden, R.E. Routine in vitro culture of P. falciparum gametocytes to evaluate novel transmission-blocking interventions. Nat. Protoc. 2016, 11, 1668–1680. [Google Scholar] [CrossRef]
- Prudent, M.; Delobel, J.; Hubner, A.; Benay, C.; Lion, N.; Tissot, J.D. Proteomics of stored red blood cell membrane and storage-induced microvesicles reveals the association of flotillin-2 with band 3 complexes. Front. Physiol. 2018, 9, 421. [Google Scholar] [CrossRef]
- Chiangjong, W.; Netsirisawan, P.; Hongeng, S.; Chutipongtanate, S. Red blood cell extracellular vesicle-based drug delivery: Challenges and opportunities. Front. Med. 2021, 8, 761362. [Google Scholar] [CrossRef]
- Thangaraju, K.; Neerukonda, S.N.; Katneni, U.; Buehler, P.W. Extracellular vesicles from red blood cells and their evolving roles in health, coagulopathy and therapy. Int. J. Mol. Sci. 2020, 22, 153. [Google Scholar] [CrossRef] [PubMed]
- Tiberti, N.; Latham, S.L.; Bush, S.; Cohen, A.; Opoka, R.O.; John, C.C.; Juillard, A.; Grau, G.E.; Combes, V. Exploring experimental cerebral malaria pathogenesis through the characterisation of host-derived plasma microparticle protein content. Sci. Rep. 2016, 6, 37871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avalos-Padilla, Y.; Georgiev, V.N.; Lantero, E.; Pujals, S.; Verhoef, R.; Borgheti-Cardoso, L.N.; Albertazzi, L.; Dimova, R.; Fernandez-Busquets, X. The ESCRT-III machinery participates in the production of extracellular vesicles and protein export during Plasmodium falciparum infection. PLoS Pathog. 2021, 17, e1009455. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.A.; Silamut, K.; Chotivanich, K.; Pukrittayakamee, S.; White, N.J. Red cell selectivity in malaria: A study of multiple-infected erythrocytes. Trans. R. Soc. Trop. Med. Hyg. 1999, 93, 165–168. [Google Scholar] [CrossRef] [Green Version]
- Chotivanich, K.; Udomsangpetch, R.; Simpson, J.A.; Newton, P.; Pukrittayakamee, S.; Looareesuwan, S.; White, N.J. Parasite multiplication potential and the severity of Falciparum malaria. J. Infect. Dis. 2000, 181, 1206–1209. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Uniprot Accession | Entry Name | Protein Name |
|---|---|---|
| Proteins detected only in MV-E | ||
| O75140 | DEPD5_HUMAN | DEPDC5-DEP domain containing 5 |
| P50226 | ST1A2_HUMAN | SULT1A2-sulfotransferase family, cytosolic, 1A, phenol-preferring, member 2 |
| P56696 | KCNQ4_HUMAN | KCNQ4-potassium voltage-gated channel, KQT-like subfamily, member 4 |
| Q14C86 | GAPD1_HUMAN | GAPVD1-GTPase activating protein and VPS9 domains 1 |
| Q5U623 | MCAF2_HUMAN | ATF7IP2-activating transcription factor 7 interacting protein 2 |
| Q86YA3 | ZGRF1_HUMAN | C4orf21-chromosome 4 open reading frame 21 |
| Q8TD57 | DYH3_HUMAN | DNAH3-dynein, axonemal, heavy chain 3 |
| Q96QD9 | UIF_HUMAN | FYTTD1-forty-two-three domain containing 1 |
| Q9HC35 | EMAL4_HUMAN | EML4-echinoderm microtubule associated protein like 4 |
| Q9NP80 | PLPL8_HUMAN | PNPLA8-patatin-like phospholipase domain containing 8 |
| Proteins detected only in MV-L | ||
| Q14692 | BMS1_HUMAN | BMS1-BMS1 homolog, ribosome assembly protein |
| Proteins detected only in Exo-E | ||
| Q13546 | RIPK1_HUMAN | RIPK1-receptor (TNFRSF)-interacting serine-threonine kinase 1 |
| Q15645 | PCH2_HUMAN | TRIP13-thyroid hormone receptor interactor 13 |
| Q86YN1 | DOPP1_HUMAN | DOLPP1-dolichyl pyrophosphate phosphatase 1 |
| Proteins detected only in Exo-L | ||
| O60308 | CE104_HUMAN | CEP104-centrosomal protein 104 kDa |
| Q09328 | MGT5A_HUMAN | MGAT5-mannosyl (alpha-1,6-)-glycoprotein beta-1,6-N-acetyl-glucosaminyltransferase |
| Q8N1B4 | VPS52_HUMAN | VPS52-vacuolar protein sorting 52 homolog (S. cerevisiae) |
| Q9Y613 | FHOD1_HUMAN | FHOD1-formin homology 2 domain containing 1 |
| Uniprot Accession | Entry Name | Protein Name | Function |
|---|---|---|---|
| C0H5F4 | RBP2B_PLAF7 | Reticulocyte binding protein 2 homolog b | Involved in reticulocyte adhesion |
| P02893 | CSP_PLAFA | Circumsporozoite protein (CS) | Immunodominant surface antigen on the sporozoite |
| P04923 | CRA_PLAFA | Circumsporozoite protein-related antigen (CRA) | Located within parasitophorous vacuole and associated with membranous structures in RBC cytoplasm |
| P04933 | MSP1_PLAFW | Merozoite surface protein 1 (Merozoite surface antigens) (PMMSA) (p195) | Pathogenesis |
| P06719 | KNOB_PLAFN | Knob-associated histidine-rich protein (KAHRP) | Mimic human histidine-rich glycoproteins to anchor host thrombospondin or a parasite analog in a binding complex with the endothelial cell receptor |
| P08569 | MSP1_PLAFM | Merozoite surface protein 1 (Merozoite surface antigens) (PMMSA) (p190) | Pathogenesis |
| P09346 | KNOB_PLAFG | Knob-associated histidine-rich protein (KAHRP) (KP) | Mimic human histidine-rich glycoproteins to anchor host thrombospondin or a parasite analog in a binding complex with the endothelial cell receptor |
| P10988 | ACT1_PLAFO | Actin-1 (Actin I) (Pf-actin I) | Contribute to parasite gliding motility |
| P11459 | RHOA_PLAFA | Rhoptry antigen protein (Fragment) | Participate in the invasion of RBCs by merozoites |
| P13816 | GARP_PLAFF | Glutamic acid-rich protein | Enhance the adhesive properties of human RBC by engaging band 3 receptor |
| P13822 | SANT_PLAFP | S-antigen protein (Fragment) | Soluble heat-stable proteins |
| P13830 | RESA_PLAFF | Ring-infected erythrocyte surface antigen | Facilitate the invagination of the red cell membrane which is necessary for the formation of the parasitophorous vacuole. |
| P13831 | RESA_PLAFN | Ring-infected erythrocyte surface antigen (Fragment) | |
| P14883 | ACT2_PLAFO | Actin-2 (Actin II) (Pf-actin II) | Contribute to parasite gliding motility |
| P16893 | TRAP_PLAFA | Thrombospondin-related anonymous protein | Cell adhesion |
| P19214 | EBA1_PLAFC | Erythrocyte-binding antigen 175 (EBA-175) | Host cell surface receptor binding |
| P19259 | PF12_PLAFA | Merozoite surface protein PF12 | Cleaved from the surface during invasion |
| P20147 | HSP90_PLAFP | Heat shock 90 kDa protein homolog (Fragment) | Molecular chaperone |
| P50489 | AMA1_PLAFC | Apical membrane antigen 1 (Merozoite surface antigen) | Involved in parasite invasion of erythrocytes |
| P62343 | CDPK1_PLAFK | Calcium-dependent protein kinase 1 (EC 2.7.11.1) (PfCDPK1) (PfCPK) | Required for microneme secretion and thus merozoite egress from and invasion of host erythrocytes |
| P69192 | SERA_PLAFG | Serine-repeat antigen protein (111 kDa antigen) (p126) | It may function at the RBC membrane, perhaps as a component that influences the invasion process |
| P86148 | RBP1_PLAF7 | Reticulocyte-binding protein PFD0110w | Involved in reticulocyte adhesion |
| Q03400 | SANT_PLAF7 | S-antigen protein | Soluble heat-stable proteins |
| Q27727 | ENO_PLAFA | Enolase (EC 4.2.1.11) (2-phospho-D-glycerate hydrolyase) (2-phosphoglycerate dehydratase) | Catalytic activity |
| Q8I2J4 | PROF_PLAF7 | Profilin | Essential for the invasive blood stages of the parasite |
| Q8I467 | CADF1_PLAF7 | Cofilin/actin-depolymerizing factor homolog 1 | Essential for erythrocytic schizogony |
| Q8I4R2 | RBP3_PLAF7 | Reticulocyte-binding protein 3 | Involved in reticulocyte adhesion |
| Q8I5D2 | ABRA_PLAF7 | 101 kDa malaria antigen (Acidic basic repeat antigen) (p101) | Located at the merozoite surface, within the parasitophorous vacuole of P. falciparum |
| Q8I6U8 | GBP_PLAF7 | Glycophorin-binding protein (GBP-130) | Involved in erythrocyte invasion |
| Q8IBS5 | CDPK4_PLAF7 | Calcium-dependent protein kinase 4 (EC 2.7.11.1) | Host RBCs and hepatocytes infection cycles, sexual reproduction and mosquito transmission of the parasite |
| Q8ID66 | PF92_PLAF7 | Merozoite surface protein PF92 | Cys-rich surface protein; binds factor H andinvolved in complement evasion |
| Q8IDR3 | MYOA_PLAF7 | Myosin-A (PfM-A) | Actin-based motor molecules with ATPase activity which critical for P. falciparum RBC invasion |
| Q8IDX6 | RBP2A_PLAF7 | Reticulocyte-binding protein 2 homolog a | Involved in reticulocyte adhesion |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vimonpatranon, S.; Roytrakul, S.; Phaonakrop, N.; Lekmanee, K.; Atipimonpat, A.; Srimark, N.; Sukapirom, K.; Chotivanich, K.; Khowawisetsut, L.; Pattanapanyasat, K. Extracellular Vesicles Derived from Early and Late Stage Plasmodium falciparum-Infected Red Blood Cells Contain Invasion-Associated Proteins. J. Clin. Med. 2022, 11, 4250. https://doi.org/10.3390/jcm11144250
Vimonpatranon S, Roytrakul S, Phaonakrop N, Lekmanee K, Atipimonpat A, Srimark N, Sukapirom K, Chotivanich K, Khowawisetsut L, Pattanapanyasat K. Extracellular Vesicles Derived from Early and Late Stage Plasmodium falciparum-Infected Red Blood Cells Contain Invasion-Associated Proteins. Journal of Clinical Medicine. 2022; 11(14):4250. https://doi.org/10.3390/jcm11144250
Chicago/Turabian StyleVimonpatranon, Sinmanus, Sittiruk Roytrakul, Narumon Phaonakrop, Kittima Lekmanee, Anyapat Atipimonpat, Narinee Srimark, Kasama Sukapirom, Kesinee Chotivanich, Ladawan Khowawisetsut, and Kovit Pattanapanyasat. 2022. "Extracellular Vesicles Derived from Early and Late Stage Plasmodium falciparum-Infected Red Blood Cells Contain Invasion-Associated Proteins" Journal of Clinical Medicine 11, no. 14: 4250. https://doi.org/10.3390/jcm11144250
APA StyleVimonpatranon, S., Roytrakul, S., Phaonakrop, N., Lekmanee, K., Atipimonpat, A., Srimark, N., Sukapirom, K., Chotivanich, K., Khowawisetsut, L., & Pattanapanyasat, K. (2022). Extracellular Vesicles Derived from Early and Late Stage Plasmodium falciparum-Infected Red Blood Cells Contain Invasion-Associated Proteins. Journal of Clinical Medicine, 11(14), 4250. https://doi.org/10.3390/jcm11144250