
Mechanistic View on the Effects of SGLT2 Inhibitors on Lipid Metabolism in Diabetic Milieu


Abstract
:1. Introduction
2. Sodium-Glucose Cotransporter-2 Inhibitors
3. Lipids
4. Lipid Metabolism in Diabetes with Focus on Adverse Effects of Diabetes on Lipoproteins
5. SGLT2 Inhibitors and Lipid Metabolism
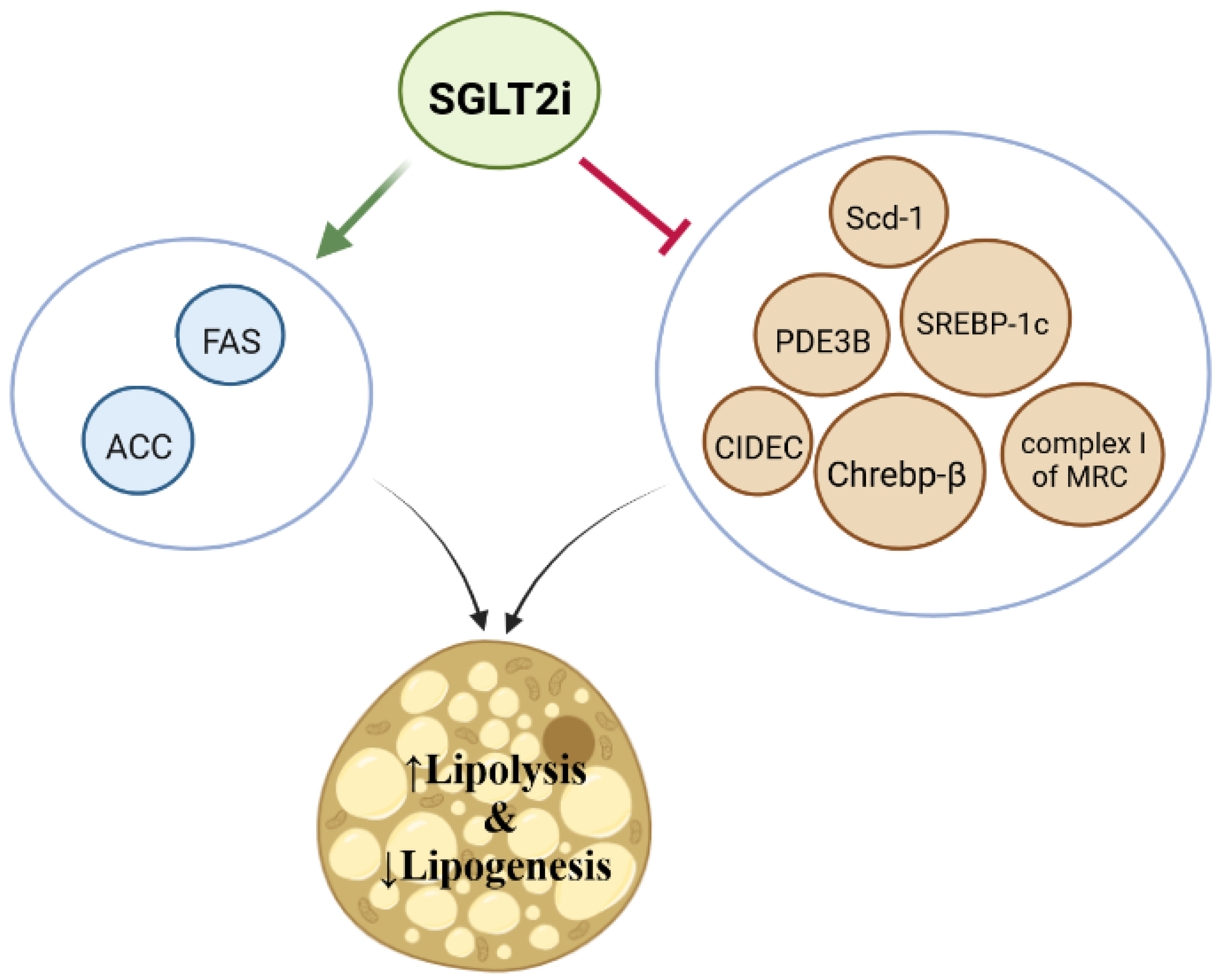
5.1. Biogenesis of Lipids
5.2. Cholesterol Homeostasis
5.3. Fatty Acid Uptake and Utilization (β-Oxidation)
5.4. Lipid Peroxidation
5.5. Lipid Absorption/Transport
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Divers, J.; Mayer-Davis, E.J.; Lawrence, J.M.; Isom, S.; Dabelea, D.; Dolan, L.; Imperatore, G.; Marcovina, S.; Pettitt, D.J.; Pihoker, C. Trends in incidence of type 1 and type 2 diabetes among youths—Selected counties and Indian reservations, United States, 2002–2015. Morb. Mortal. Wkly. Rep. 2020, 69, 161. [Google Scholar] [CrossRef] [PubMed]
- Forbes, J.M.; Cooper, M.E. Mechanisms of diabetic complications. Physiol. Rev. 2013, 93, 137–188. [Google Scholar] [CrossRef] [PubMed]
- Athyros, V.G.; Doumas, M.; Imprialos, K.P.; Stavropoulos, K.; Georgianou, E.; Katsimardou, A.; Karagiannis, A. Diabetes and lipid metabolism. Hormones 2018, 17, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Yaribeygi, H.; Bo, S.; Ruscica, M.; Sahebkar, A. Ceramides and diabetes mellitus: An update on the potential molecular relationships. Diabet. Med. 2020, 37, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Schoeler, M.; Caesar, R. Dietary lipids, gut microbiota and lipid metabolism. Rev. Endocr. Metab. Disord. 2019, 20, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Carpentier, A.C. Abnormal myocardial dietary fatty acid metabolism and diabetic cardiomyopathy. Can. J. Cardiol. 2018, 34, 605–614. [Google Scholar] [CrossRef]
- Iacobini, C.; Vitale, M.; Pesce, C.; Pugliese, G.; Menini, S. Diabetic complications and oxidative stress: A 20-year voyage back in time and back to the future. Antioxidants 2021, 10, 727. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Sathyapalan, T.; Maleki, M.; Jamialahmadi, T.; Sahebkar, A. Molecular mechanisms by which SGLT2 inhibitors can induce insulin sensitivity in diabetic milieu: A mechanistic review. Life Sci. 2020, 240, 117090. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Simental-Mendía, L.E.; Banach, M.; Bo, S.; Sahebkar, A. The major molecular mechanisms mediating the renoprotective effects of SGLT2 inhibitors: An update. Biomed. Pharmacother. 2019, 120, 109526. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Atkin, S.L.; Sahebkar, A. Mechanistic effects of SGLT2 inhibition on blood pressure in diabetes. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 1679–1683. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Katsiki, N.; Butler, A.E.; Sahebkar, A. Effects of antidiabetic drugs on NLRP3 inflammasome activity, with a focus on diabetic kidneys. Drug Discov. Today 2019, 24, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Yaribeygi, H.; Atkin, S.L.; Jamialahmadi, T.; Sahebkar, A. A review on the effects of new anti-diabetic drugs on platelet function. Endocr. Metab. Immune Disord. Drug Targets 2020, 20, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ma, X.; Ilyas, I.; Zheng, X.; Luo, S.; Little, P.J.; Kamato, D.; Sahebkar, A.; Wu, W.; Weng, J.; et al. Impact of sodium glucose cotransporter 2 (SGLT2) inhibitors on atherosclerosis: From pharmacology to pre-clinical and clinical therapeutics. Theranostics 2021, 11, 4502–4515. [Google Scholar] [CrossRef] [PubMed]
- Ranjbar, G.; Mikhailidis, D.P.; Sahebkar, A. Effects of newer antidiabetic drugs on nonalcoholic fatty liver and steatohepatitis: Think out of the box! Metab. Clin. Exp. 2019, 101, 154001. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Ashrafizadeh, M.; Henney, N.C.; Sathyapalan, T.; Jamialahmadi, T.; Sahebkar, A. Neuromodulatory effects of anti-diabetes medications: A mechanistic review. Pharmacol. Res. 2020, 152, 104611. [Google Scholar] [CrossRef]
- Szekeres, Z.; Toth, K.; Szabados, E. The Effects of SGLT2 Inhibitors on Lipid Metabolism. Metabolites 2021, 11, 87. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Atkin, S.L.; Butler, A.E.; Sahebkar, A. Sodium–glucose cotransporter inhibitors and oxidative stress: An update. J. Cell. Physiol. 2019, 234, 3231–3237. [Google Scholar] [CrossRef]
- Davidson, J.A.; Kuritzky, L. Sodium glucose co-transporter 2 inhibitors and their mechanism for improving glycemia in patients with type 2 diabetes. Postgrad. Med. 2014, 126, 33–48. [Google Scholar] [CrossRef]
- Gronda, E.; Lopaschuk, G.D.; Arduini, A.; Santoro, A.; Benincasa, G.; Palazzuoli, A.; Gabrielli, D.; Napoli, C. Mechanisms of action of SGLT2 inhibitors and their beneficial effects on the cardiorenal axis. Can. J. Physiol. Pharmacol. 2022, 100, 93–106. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Butler, A.E.; Atkin, S.L.; Katsiki, N.; Sahebkar, A. Sodium–glucose cotransporter 2 inhibitors and inflammation in chronic kidney disease: Possible molecular pathways. J. Cell. Physiol. 2019, 234, 223–230. [Google Scholar] [CrossRef]
- Takasu, T.; Yokono, M.; Tahara, A.; Takakura, S. In vitro pharmacological profile of ipragliflozin, a sodium glucose co-transporter 2 Inhibitor. Biol. Pharm. Bull. 2019, 42, 507–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, E.C. SGLT-2 inhibitors: A new mechanism for glycemic control. Clin. Diabetes 2014, 32, 4–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, E.C.; Henry, R.R. SGLT2 inhibition—A novel strategy for diabetes treatment. Nat. Rev. Drug Discov. 2010, 9, 551. [Google Scholar] [CrossRef]
- Clar, C.; Gill, J.A.; Waugh, N. Systematic review of SGLT2 receptor inhibitors in dual or triple therapy in type 2 diabetes. BMJ Open 2012, 2, e001007. [Google Scholar] [CrossRef]
- Kern, M.; Klöting, N.; Mark, M.; Mayoux, E.; Klein, T.; Blüher, M. The SGLT2 inhibitor empagliflozin improves insulin sensitivity in db/db mice both as monotherapy and in combination with linagliptin. Metab. Clin. Exp. 2016, 65, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Hagan, D.L.; Taylor, J.R.; Xin, L.; Meng, W.; Biller, S.A.; Wetterau, J.R.; Washburn, W.N.; Whaley, J.M. Dapagliflozin, a selective SGLT2 inhibitor, improves glucose homeostasis in normal and diabetic rats. Diabetes 2008, 57, 1723–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilding, J.; Woo, V.; Rohwedder, K.; Sugg, J.; Parikh, S. Dapagliflozin in patients with type 2 diabetes receiving high doses of insulin: Efficacy and safety over 2 years. Diabetes Obes. Metab. 2014, 16, 124–136. [Google Scholar] [CrossRef]
- Ferrannini, E.; Muscelli, E.; Frascerra, S.; Baldi, S.; Mari, A.; Heise, T.; Broedl, U.C.; Woerle, H.-J. Metabolic response to sodium-glucose cotransporter 2 inhibition in type 2 diabetic patients. J. Clin. Investig. 2014, 124, 499–508. [Google Scholar] [CrossRef] [Green Version]
- Reddy, R.M.; Inzucchi, S.E. SGLT2 inhibitors in the management of type 2 diabetes. Endocrine 2016, 53, 364–372. [Google Scholar] [CrossRef]
- Hædersdal, S.; Lund, A.; Nielsen-Hannerup, E.; Maagensen, H.; van Hall, G.; Holst, J.J.; Knop, F.K.; Vilsbøll, T. The role of glucagon in the acute therapeutic effects of SGLT2 inhibition. Diabetes 2020, 69, 2619–2629. [Google Scholar] [CrossRef]
- Shyr, Z.A.; Yan, Z.; Ustione, A.; Egan, E.M.; Remedi, M.S. SGLT2 inhibitors therapy protects glucotoxicity-induced β-cell failure in a mouse model of human KATP-induced diabetes through mitigation of oxidative and ER stress. PLoS ONE 2022, 17, e0258054. [Google Scholar] [CrossRef] [PubMed]
- Thiam, A.R.; Farese, R.V., Jr.; Walther, T.C. The biophysics and cell biology of lipid droplets. Nat. Rev. Mol. Cell Biol. 2013, 14, 775–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandel, N.S. Lipid metabolism. Cold Spring Harb. Perspect. Biol. 2021, 13, a040576. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.E.; Hall, M.E. Guyton and Hall Textbook of Medical Physiology e-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Yaribeygi, H.; Maleki, M.; Butler, A.E.; Jamialahmadi, T.; Sahebkar, A. The Impact of Incretin-Based Medications on Lipid Metabolism. J. Diabetes Res. 2021, 2021, 1815178. [Google Scholar] [CrossRef]
- Aguilar-Ballester, M.; Herrero-Cervera, A.; Vinué, Á.; Martínez-Hervás, S.; González-Navarro, H. Impact of cholesterol metabolism in immune cell function and atherosclerosis. Nutrients 2020, 12, 2021. [Google Scholar] [CrossRef]
- White, U.; Ravussin, E. Dynamics of adipose tissue turnover in human metabolic health and disease. Diabetologia 2019, 62, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Yaribeygi, H.; Simental-Mendía, L.E.; Barreto, G.E.; Sahebkar, A. Metabolic effects of antidiabetic drugs on adipocytes and adipokine expression. J. Cell. Physiol. 2019, 234, 16987–16997. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Farrokhi, F.R.; Butler, A.E.; Sahebkar, A. Insulin resistance: Review of the underlying molecular mechanisms. J. Cell. Physiol. 2019, 234, 8152–8161. [Google Scholar] [CrossRef]
- Ponziani, F.R.; Pecere, S.; Gasbarrini, A.; Ojetti, V. Physiology and pathophysiology of liver lipid metabolism. Expert Rev. Gastroenterol. Hepatol. 2015, 9, 1055–1067. [Google Scholar] [CrossRef]
- Ko, C.-W.; Qu, J.; Black, D.D.; Tso, P. Regulation of intestinal lipid metabolism: Current concepts and relevance to disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 169–183. [Google Scholar] [CrossRef]
- Cisa-Wieczorek, S.; Hernández-Alvarez, M.I. Deregulation of Lipid Homeostasis: A Fa (c) t in the Development of Metabolic Diseases. Cells 2020, 9, 2605. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.-H.; Duann, P. Dyslipidemia in kidney disorders: Perspectives on mitochondria homeostasis and therapeutic opportunities. Front. Physiol. 2020, 11, 1050. [Google Scholar] [CrossRef] [PubMed]
- Nishi, H.; Higashihara, T.; Inagi, R. Lipotoxicity in kidney, heart, and skeletal muscle dysfunction. Nutrients 2019, 11, 1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, J.; Zhang, C.-J.; Zhu, N.; Du, K.; Yin, Y.-F.; Tan, X.; Liao, D.-F.; Qin, L. Lipid metabolism and carcinogenesis, cancer development. Am. J. Cancer Res. 2018, 8, 778. [Google Scholar] [PubMed]
- Chen, L.; Chen, X.-W.; Huang, X.; Song, B.-L.; Wang, Y.; Wang, Y. Regulation of glucose and lipid metabolism in health and disease. Sci. China Life Sci. 2019, 62, 1420–1458. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Cuevas, J.; Santos, A.; Armendariz-Borunda, J. Pathophysiological molecular mechanisms of obesity: A link between MAFLD and NASH with cardiovascular diseases. Int. J. Mol. Sci. 2021, 22, 11629. [Google Scholar] [CrossRef]
- Gutiérrez-Cuevas, J.; Sandoval-Rodriguez, A.; Meza-Rios, A.; Monroy-Ramírez, H.C.; Galicia-Moreno, M.; García-Bañuelos, J.; Santos, A.; Armendariz-Borunda, J. Molecular mechanisms of obesity-linked cardiac dysfunction: An up-date on current knowledge. Cells 2021, 10, 629. [Google Scholar] [CrossRef]
- Séguro, F.; Rabès, J.P.; Taraszkiewicz, D.; Ruidavets, J.B.; Bongard, V.; Ferrières, J. Genetic diagnosis of familial hypercholesterolemia is associated with a premature and high coronary heart disease risk. Clin. Cardiol. 2018, 41, 385–391. [Google Scholar] [CrossRef]
- Penno, G.; Solini, A.; Zoppini, G.; Fondelli, C.; Trevisan, R.; Vedovato, M.; Gruden, G.; Lamacchia, O.; Pontiroli, A.E.; Arosio, M. Hypertriglyceridemia is independently associated with renal, but not retinal complications in subjects with type 2 diabetes: A cross-sectional analysis of the Renal Insufficiency And Cardiovascular Events (RIACE) Italian Multicenter Study. PLoS ONE 2015, 10, e0125512. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, C.; Shi, K.; Yin, X. Relationship between dyslipidemia and diabetic retinopathy: A systematic review and meta-analysis. Medicine 2018, 97, e12283. [Google Scholar] [CrossRef]
- Jeng, C.-J.; Hsieh, Y.-T.; Yang, C.-M.; Yang, C.-H.; Lin, C.-L.; Wang, I.-J. Diabetic retinopathy in patients with dyslipidemia: Development and progression. Ophthalmol. Retin. 2018, 2, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Haile, K.; Timerga, A. Dyslipidemia and its associated risk factors among adult type-2 diabetic patients at Jimma University Medical Center, Jimma, Southwest Ethiopia. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 4589. [Google Scholar] [CrossRef] [PubMed]
- Savelieff, M.G.; Callaghan, B.C.; Feldman, E.L. The emerging role of dyslipidemia in diabetic microvascular complications. Curr. Opin. Endocrinol. Diabetes Obes. 2020, 27, 115–123. [Google Scholar] [CrossRef]
- Garg, R.; Aggarwal, S.; Kumar, R.; Sharma, G. Association of atherosclerosis with dyslipidemia and co-morbid conditions: A descriptive study. J. Nat. Sci. Biol. Med. 2015, 6, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savransky, V.; Jun, J.; Li, J.; Nanayakkara, A.; Fonti, S.; Moser, A.B.; Steele, K.E.; Schweitzer, M.A.; Patil, S.P.; Bhanot, S. Dyslipidemia and atherosclerosis induced by chronic intermittent hypoxia are attenuated by deficiency of stearoyl coenzyme a desaturase. Circ. Res. 2008, 103, 1173–1180. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.-L.; Shi, Y.-H.; Hao, G.; Li, W.; Le, G.-W. Increasing oxidative stress with progressive hyperlipidemia in human: Relation between malondialdehyde and atherogenic index. J. Clin. Biochem. Nutr. 2008, 43, 154–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Feng, D.; Wang, Q.; Abdulla, A.; Xie, X.-J.; Zhou, J.; Sun, Y.; Yang, E.S.; Liu, L.-P.; Vaitheesvaran, B. Regulation of lipogenesis by cyclin-dependent kinase 8–mediated control of SREBP-1. J. Clin. Investig. 2012, 122, 2417–2427. [Google Scholar] [CrossRef]
- Ducheix, S.; Lobaccaro, J.; Martin, P.; Guillou, H. Liver X Receptor: An oxysterol sensor and a major player in the control of lipogenesis. Chem. Phys. Lipids 2011, 164, 500–514. [Google Scholar] [CrossRef]
- Wang, Y.; Viscarra, J.; Kim, S.-J.; Sul, H.S. Transcriptional regulation of hepatic lipogenesis. Nat. Rev. Mol. Cell Biol. 2015, 16, 678–689. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Xiaoli, A.M.; Yang, F. Regulation and metabolic significance of de novo lipogenesis in adipose tissues. Nutrients 2018, 10, 1383. [Google Scholar] [CrossRef]
- Tracey, T.J.; Steyn, F.J.; Wolvetang, E.J.; Ngo, S.T. Neuronal lipid metabolism: Multiple pathways driving functional outcomes in health and disease. Front. Mol. Neurosci. 2018, 11, 10. [Google Scholar] [CrossRef] [Green Version]
- Ahmadian, M.; Wang, Y.; Sul, H.S. Lipolysis in adipocytes. Int. J. Biochem. Cell Biol. 2010, 42, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Lin, B.; Zheng, X.; Chen, Z.; Cao, H.; Xu, H.; Liang, H.; Weng, J. GLP-1 receptor agonist promotes brown remodelling in mouse white adipose tissue through SIRT1. Diabetologia 2016, 59, 1059–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, F.W.; Griffin, J.L. De novo lipogenesis in the liver in health and disease: More than just a shunting yard for glucose. Biol. Rev. 2016, 91, 452–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, R.E.; Ahmadian, M.; Jaworski, K.; Sarkadi-Nagy, E.; Sul, H.S. Regulation of lipolysis in adipocytes. Annu. Rev. Nutr. 2007, 27, 79–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jojima, T.; Tomotsune, T.; Iijima, T.; Akimoto, K.; Suzuki, K.; Aso, Y. Empagliflozin (an SGLT2 inhibitor), alone or in combination with linagliptin (a DPP-4 inhibitor), prevents steatohepatitis in a novel mouse model of non-alcoholic steatohepatitis and diabetes. Diabetol. Metab. Syndr. 2016, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lauritsen, K.M.; Voigt, J.H.; Pedersen, S.B.; Hansen, T.K.; Møller, N.; Jessen, N.; Gormsen, L.C.; Søndergaard, E. Effects of SGLT2 inhibition on lipid transport in adipose tissue in type 2 diabetes. Endocr. Connect. 2022, 11, e210558. [Google Scholar] [CrossRef]
- Osataphan, S.; Macchi, C.; Singhal, G.; Chimene-Weiss, J.; Sales, V.; Kozuka, C.; Dreyfuss, J.M.; Pan, H.; Tangcharoenpaisan, Y.; Morningstar, J. SGLT2 inhibition reprograms systemic metabolism via FGF21-dependent and-independent mechanisms. JCI Insight 2019, 4, e123130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, E.A.; Ford, R.J.; Lu, J.H.; Lu, R.; Lundenberg, L.; Desjardins, E.M.; Green, A.E.; Lally, J.S.; Schertzer, J.D.; Steinberg, G.R. The SGLT2 inhibitor canagliflozin suppresses lipid synthesis and interleukin-1 beta in ApoE deficient mice. Biochem. J. 2020, 477, 2347–2361. [Google Scholar] [CrossRef] [PubMed]
- Hawley, S.A.; Ford, R.J.; Smith, B.K.; Gowans, G.J.; Mancini, S.J.; Pitt, R.D.; Day, E.A.; Salt, I.P.; Steinberg, G.R.; Hardie, D.G. The Na+/glucose cotransporter inhibitor canagliflozin activates AMPK by inhibiting mitochondrial function and increasing cellular AMP levels. Diabetes 2016, 65, 2784–2794. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Q. Cholesterol metabolism and homeostasis in the brain. Protein Cell 2015, 6, 254–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groenen, A.G.; Halmos, B.; Tall, A.R.; Westerterp, M. Cholesterol efflux pathways, inflammation, and atherosclerosis. Crit. Rev. Biochem. Mol. Biol. 2021, 56, 426–439. [Google Scholar] [CrossRef]
- Basu, D.; Huggins, L.-A.; Scerbo, D.; Obunike, J.; Mullick, A.E.; Rothenberg, P.L.; Di Prospero, N.A.; Eckel, R.H.; Goldberg, I.J. Mechanism of increased LDL (low-density lipoprotein) and decreased triglycerides with SGLT2 (sodium-glucose cotransporter 2) inhibition. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 2207–2216. [Google Scholar] [CrossRef] [PubMed]
- Fadini, G.P.; Bonora, B.M.; Zatti, G.; Vitturi, N.; Iori, E.; Marescotti, M.C.; Albiero, M.; Avogaro, A. Effects of the SGLT2 inhibitor dapagliflozin on HDL cholesterol, particle size, and cholesterol efflux capacity in patients with type 2 diabetes: A randomized placebo-controlled trial. Cardiovasc. Diabetol. 2017, 16, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gürkan, E. Effects of dapagliflozin on serum low-density lipoprotein cholesterol and triglyceride levels. Eur. J. Ther. 2020, 26, 76–80. [Google Scholar]
- Ejiri, K.; Miyoshi, T.; Kihara, H.; Hata, Y.; Nagano, T.; Takaishi, A.; Toda, H.; Namba, S.; Nakamura, Y.; Akagi, S. Effects of luseogliflozin and voglibose on high-risk lipid profiles and inflammatory markers in diabetes patients with heart failure. Sci. Rep. 2022, 12, 15449. [Google Scholar] [CrossRef]
- Sanchez-Garcia, A.; Simental-Mendia, M.; Millan-Alanis, J.M.; Simental-Mendia, L.E. Effect of sodium-glucose co-transporter 2 inhibitors on lipid profile: A systematic review and meta-analysis of 48 randomized controlled trials. Pharmacol. Res. 2020, 160, 105068. [Google Scholar] [CrossRef]
- Kamijo, Y.; Ishii, H.; Yamamoto, T.; Kobayashi, K.; Asano, H.; Miake, S.; Kanda, E.; Urata, H.; Yoshida, M. Potential impact on lipoprotein subfractions in type 2 diabetes. Clin. Med. Insights Endocrinol. Diabetes 2019, 12, 1179551419866811. [Google Scholar] [CrossRef] [Green Version]
- Shi, F.H.; Li, H.; Shen, L.; Fu, J.J.; Ma, J.; Gu, Z.C.; Lin, H.W. High-dose sodium-glucose co-transporter-2 inhibitors are superior in type 2 diabetes: A meta-analysis of randomized clinical trials. Diabetes Obes. Metab. 2021, 23, 2125–2136. [Google Scholar] [CrossRef]
- Cha, S.-A.; Park, Y.-M.; Yun, J.-S.; Lim, T.-S.; Song, K.-H.; Yoo, K.-D.; Ahn, Y.-B.; Ko, S.-H. A comparison of effects of DPP-4 inhibitor and SGLT2 inhibitor on lipid profile in patients with type 2 diabetes. Lipids Health Dis. 2017, 16, 58. [Google Scholar] [CrossRef] [Green Version]
- Bosch, A.; Ott, C.; Jung, S.; Striepe, K.; Karg, M.V.; Kannenkeril, D.; Dienemann, T.; Schmieder, R.E. How does empagliflozin improve arterial stiffness in patients with type 2 diabetes mellitus? Sub analysis of a clinical trial. Cardiovasc. Diabetol. 2019, 18, 44. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Temkin, S.M.; Hawkridge, A.M.; Guo, C.; Wang, W.; Wang, X.-Y.; Fang, X. Fatty acid oxidation: An emerging facet of metabolic transformation in cancer. Cancer Lett. 2018, 435, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Al Batran, R.; Almutairi, M.; Ussher, J.R. Glucagon-like peptide-1 receptor mediated control of cardiac energy metabolism. Peptides 2018, 100, 94–100. [Google Scholar] [CrossRef]
- Wu, S.; Näär, A.M. SREBP1-dependent de novo fatty acid synthesis gene expression is elevated in malignant melanoma and represents a cellular survival trait. Sci. Rep. 2019, 9, 10369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallenius, K.; Kroon, T.; Hagstedt, T.; Löfgren, L.; Sörhede-Winzell, M.; Boucher, J.; Lindén, D.; Oakes, N.D. The SGLT2 inhibitor dapagliflozin promotes systemic FFA mobilization, enhances hepatic β-oxidation, and induces ketosis. J. Lipid Res. 2022, 63, 100176. [Google Scholar] [CrossRef]
- Lauritsen, K.M.; Nielsen, B.R.; Tolbod, L.P.; Johannsen, M.; Hansen, J.; Hansen, T.K.; Wiggers, H.; Møller, N.; Gormsen, L.C.; Søndergaard, E. SGLT2 inhibition does not affect myocardial fatty acid oxidation or uptake, but reduces myocardial glucose uptake and blood flow in individuals with type 2 diabetes: A randomized double-blind, placebo-controlled crossover trial. Diabetes 2021, 70, 800–808. [Google Scholar] [CrossRef] [PubMed]
- Herring, R.A.; Shojaee-Moradie, F.; Stevenage, M.; Parsons, I.; Jackson, N.; Mendis, J.; Middleton, B.; Umpleby, A.M.; Fielding, B.A.; Davies, M. The SGLT2 Inhibitor Dapagliflozin Increases the Oxidation of Ingested Fatty Acids to Ketones in Type 2 Diabetes. Diabetes Care 2022, 45, 1408–1415. [Google Scholar] [CrossRef] [PubMed]
- Kroon, T.; Hagstedt, T.; Kjellstedt, A.; Lindblom, A.; Löfgren, L.; Boucher, J.; Sörhede Winzell, M.; Linden, D.; Wallenius, K. 1881-P: The SGLT2 Inhibitor Dapagliflozin Improves Glucose Control and Enhances Liver Fatty Acid Oxidation and Ketone Body Formation in Insulin Resistant Obese Zucker Rats. Diabetes 2019, 68, 1881. [Google Scholar] [CrossRef]
- Flores, I.; Shannon, C.; Fourcaudot, M.; Bakewell, T.; Norton, L. 165-LB: The SGLT2 Inhibitor Canagliflozin Prevents Increased Lipid Oxidation in the Heart following High-Fat Feeding. Diabetes 2019, 68, 165-LB. [Google Scholar] [CrossRef]
- Bartlett, K.; Eaton, S. Mitochondrial β-oxidation. Eur. J. Biochem. 2004, 271, 462–469. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxidative Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Mas-Bargues, C.; Escrivá, C.; Dromant, M.; Borrás, C.; Viña, J. Lipid peroxidation as measured by chromatographic determination of malondialdehyde. Human plasma reference values in health and disease. Arch. Biochem. Biophys. 2021, 709, 108941. [Google Scholar] [CrossRef]
- Ursini, F.; Maiorino, M. Lipid peroxidation and ferroptosis: The role of GSH and GPx4. Free. Radic. Biol. Med. 2020, 152, 175–185. [Google Scholar] [CrossRef]
- Marnett, L.J. Lipid peroxidation—DNA damage by malondialdehyde. Mutat. Res. Fundam. Mol. Mech. Mutagen. 1999, 424, 83–95. [Google Scholar] [CrossRef]
- Basu, S.; De, D.; Khanna, H.D.; Kumar, A. Lipid peroxidation, DNA damage and total antioxidant status in neonatal hyperbilirubinemia. J. Perinatol. 2014, 34, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.-J.; Liu, B.-H.; Wan, S.-J.; Cheng, Y.; Zhou, S.-M.; Sun, Y.; Yao, X.-M.; Hua, Q.; Meng, X.-J.; Cheng, J.-H. A SGLT2 Inhibitor dapagliflozin alleviates diabetic cardiomyopathy by suppressing high glucose-induced oxidative stress in vivo and in vitro. Front. Pharmacol. 2021, 12, 1756. [Google Scholar] [CrossRef]
- Kimura, Y.; Kuno, A.; Tanno, M.; Sato, T.; Ohno, K.; Shibata, S. Canagliflozin, an SGLT2 inhibitor, normalizes renal susceptibility to type 1 cardiorenal syndrome through reduction of renal oxidative stress in diabetic rats. J. Diabetes Investig. 2019, 10, 933–946. [Google Scholar] [CrossRef]
- Oshima, H.; Miki, T.; Kuno, A.; Mizuno, M.; Sato, T.; Tanno, M.; Yano, T.; Nakata, K.; Kimura, Y.; Abe, K. Empagliflozin, an SGLT2 inhibitor, reduced the mortality rate after acute myocardial infarction with modification of cardiac metabolomes and antioxidants in diabetic rats. J. Pharmacol. Exp. Ther. 2019, 368, 524–534. [Google Scholar] [CrossRef] [Green Version]
- Maurea, N.; Quagliariello, V.; Bonelli, A.; Caronna, A.; Grimaldi, I.; Lombari, C.; Conforti, G.; Botti, G. 1969P The SGLT2 inhibitor dapagliflozin enhanced anticancer activities and exerts cardioprotective effects against doxorubicin and trastuzumab toxicity through TLR4, MyD88, NF-kB signaling and NLRP3 inflammasome pathway. Ann. Oncol. 2020, 31, S1106. [Google Scholar] [CrossRef]
- Lichtenstein, A.H.; Jones, P.J. Lipids: Absorption and transport. Present Knowl. Nutr. 2012, 1, 111–117. [Google Scholar]
- Feingold, K.R.; Grunfeld, C. Introduction to Lipids and Lipoproteins; MDText.com, Inc.: South Dartmouth, MA, USA, 2015. [Google Scholar]
- Arvind, A.; Osganian, S.A.; Cohen, D.E.; Corey, K.E. Lipid and Lipoprotein Metabolism in Liver Disease; MDText.com, Inc.: South Dartmouth, MA, USA, 2019. [Google Scholar]
- Li, W.; Xu, X.; Dong, D.; Lei, T.; Ou, H. Up-regulation of thioredoxin system by puerarin inhibits lipid uptake in macrophages. Free Radic. Biol. Med. 2021, 162, 542–554. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Location (Kidney) | Involvement in Glucose Reabsorption | Affinity for Glucose | Capacity | Na+/Glucose Transport Ratio | Main Inhibitors |
|---|---|---|---|---|---|---|
| SGLT2 | S1 and S2 segment of proximal tubules | ~90% | Low | High | 1:1 | Dapagliflozin, Canagliflozin, Luseogliflozin, Sotagliflozin |
| Lipid Metabolism | Used SGLT2 Inhibitors | Effects of SGLT2 Inhibition | Refs. | Clinical Evidence |
|---|---|---|---|---|
| Lipogenesis and Lipolysis | Empagliflozin, Canagliflozin | Induces lipolysis and inhibits lipogenesis | [67,69] | [68] |
| Lipid Peroxidation | Empagliflozin, Dapagliflozin, Canagliflozin | Reduces lipid peroxidation and oxidative damages | [97,98,99,100] | [77] |
| Fatty Acid β-Oxidation | Dapagliflozin | Induces and promotes FFAs oxidation/utilization | [86] | [87,88] |
| Cholesterol Homeostasis | Canagliflozin | Increases cholesterol level | [74] | [81] |
| Canagliflozin | Decreases circulating cholesterol levels | [69] | [76] | |
| Luseogliflozin, Dapagliflozin, Empagliflozin | No significant effects | - | [75,77,82] | |
| Lipid Absorption/Transport | Dapagliflozin | Mobilizes ingested/stored lipids and diverts them from adipose tissues | [86] | [68] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yaribeygi, H.; Maleki, M.; Reiner, Ž.; Jamialahmadi, T.; Sahebkar, A. Mechanistic View on the Effects of SGLT2 Inhibitors on Lipid Metabolism in Diabetic Milieu. J. Clin. Med. 2022, 11, 6544. https://doi.org/10.3390/jcm11216544
Yaribeygi H, Maleki M, Reiner Ž, Jamialahmadi T, Sahebkar A. Mechanistic View on the Effects of SGLT2 Inhibitors on Lipid Metabolism in Diabetic Milieu. Journal of Clinical Medicine. 2022; 11(21):6544. https://doi.org/10.3390/jcm11216544
Chicago/Turabian StyleYaribeygi, Habib, Mina Maleki, Željko Reiner, Tannaz Jamialahmadi, and Amirhossein Sahebkar. 2022. "Mechanistic View on the Effects of SGLT2 Inhibitors on Lipid Metabolism in Diabetic Milieu" Journal of Clinical Medicine 11, no. 21: 6544. https://doi.org/10.3390/jcm11216544
APA StyleYaribeygi, H., Maleki, M., Reiner, Ž., Jamialahmadi, T., & Sahebkar, A. (2022). Mechanistic View on the Effects of SGLT2 Inhibitors on Lipid Metabolism in Diabetic Milieu. Journal of Clinical Medicine, 11(21), 6544. https://doi.org/10.3390/jcm11216544