PRDM16 Gene Polymorphism Is Associated with Obesity and Blood Lipids Profiles in Saudi Population

Abstract
:1. Introduction
2. Methods and Materials
2.1. Participant Recruitment
2.2. Sample Collection
2.3. DNA Extraction
2.4. SNP Genotyping
2.5. Blood Lipid Profile
2.6. Statistical Analysis
3. Results
3.1. Characteristics of the Study Participants
3.2. Genotype Frequencies and Distribution
3.3. Association with Risk of Obesity
3.4. Association with BMI and Blood Lipids Profile
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Levian, C.; Ruiz, E.; Yang, X. The pathogenesis of obesity from a genomic and systems biology perspective. Yale J. Biol. Med. 2014, 87, 113–126. [Google Scholar] [PubMed]
- Memish, Z.A.; El Bcheraoui, C.; Tuffaha, M.; Robinson, M.; Daoud, F.; Jaber, S.; Mikhitarian, S.; Al Saeedi, M.; AlMazroa, M.A.; Mokdad, A.H.; et al. Obesity and Associated Factors—Kingdom of Saudi Arabia, 2013. Prev. Chronic Dis. 2014, 11, E174. [Google Scholar] [CrossRef] [PubMed]
- Ricquier, D. Respiration uncoupling and metabolism in the control of energy expenditure. Proc. Nutr. Soc. 2005, 64, 47–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasparetti, A.L.; de Souza, C.T.; Pereira-da-Silva, M.; Oliveira, R.L.G.S.; Saad, M.J.A.; Carneiro, E.M.; Velloso, L.A. Cold exposure induces tissue-specific modulation of the insulin-signalling pathway in Rattus norvegicus. J. Physiol. 2003, 552 Pt 1, 149–162. [Google Scholar] [CrossRef]
- Labbé, S.M.; Caron, A.; Bakan, I.; Laplante, M.; Carpentier, A.C.; Lecomte, R.; Richard, D. In vivo measurement of energy substrate contribution to cold-induced brown adipose tissue thermogenesis. FASEB J. 2015, 29, 2046–2058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diehl, A.M.; Hoek, J.B. Mitochondrial uncoupling: Role of uncoupling protein anion carriers and relationship to thermogenesis and weight control “The benefits of losing control”. J. Bioenerget. Biomembr. 1999, 31, 493–506. [Google Scholar] [CrossRef]
- Tiraby, C.; Langin, D. Conversion from white to brown adipocytes: A strategy for the control of fat mass? Trends Endocrinol. Metab. 2003, 14, 439–441. [Google Scholar] [CrossRef] [PubMed]
- Tiraby, C.; Tavernier, G.; Lefort, C.; Larrouy, D.; Bouillaud, F.; Ricquier, D.; Langin, D. Acquirement of brown fat cell features by human white adipocytes. J. Biol. Chem. 2003, 278, 33370–33376. [Google Scholar] [CrossRef] [PubMed]
- Carey, A.L.; Formosa, M.F.; Van Every, B.; Bertovic, D.; Eikelis, N.; Lambert, G.W.; Kalff, V.; Duffy, S.J.; Cherk, M.H.; Kingwell, B.A. Ephedrine activates brown adipose tissue in lean but not obese humans. Diabetologia 2013, 56, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Van Marken Lichtenbelt, W.D.; Vanhommerig, J.W.; Smulders, N.M.; Drossaerts, J.M.; Kemerink, G.J.; Bouvy, N.D.; Schrauwen, F.; Teule, G.J.J. Cold-activated brown adipose tissue in healthy men. N. Engl. J. Med. 2009, 360, 1500–1508. [Google Scholar] [CrossRef] [PubMed]
- Christian, M.; Parker, M.G. The engineering of brown fat. J. Mol. Cell Biol. 2010, 2, 23–25. [Google Scholar] [CrossRef] [PubMed]
- Seale, P.; Conroe, H.M.; Estall, J.; Kajimura, S.; Frontini, A.; Ishibashi, J.; Cohen, P.; Cinti, S.; Spiegelman, B.M. Prdm16 determines the thermogenic program of subcutaneous white adipose tissue in mice. J. Clin. Investig. 2011, 121, 96–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seale, P.; Kajimura, S.; Yang, W.; Chin, S.; Rohas, L.M.; Uldry, M.; Tavernier, G.; Langin, D.; Spiegelman, B.M. Transcriptional control of brown fat determination by PRDM16. Cell Metab. 2007, 6, 38–54. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Dai, Y.; Bian, L.; Wang, W.; Muramatsu, M.; Hua, Q. Association of the cell death-inducing DNA fragmentation factor alpha-like effector A (CIDEA) gene V115F (G/T) polymorphism with phenotypes of metabolic syndrome in a Chinese population. Diabetes Res. Clin. Pract. 2011, 91, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Kraynik, S.M.; Miyaoka, R.S.; Beavo, J.A. PDE3 and PDE4 Isozyme-Selective Inhibitors Are both required for synergistic activation of brown adipose tissue. Mol. Pharmacol. 2013, 83, 1155–1165. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; He, J.; Ke, Y.; Zhang, H.; Wang, C.; Hu, W.; Gu, J.M.; Fu, W.Z.; Hu, Y.Q.; Li, M.; et al. Association of single nucleotide polymorphism Rs2236518 in PRDM16 gene with BMI in Chinese males. Acta Pharmacol. Sin. 2013, 34, 710–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urano, T.; Shiraki, M.; Sasaki, N.; Ouchi, Y.; Inoue, S. Large-scale analysis reveals a functional single-nucleotide polymorphism in the 5’-flanking region of PRDM16 gene associated with lean body mass. Aging Cell 2014, 13, 739–743. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Abumrad, N.A. Cellular fatty acid uptake: A pathway under construction. Trends Endocrinol. Metab. 2009, 20, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Ran, C.; Graae, L.; Magnusson, P.K.E.; Pedersen, N.L.; Olson, L.; Belin, A.C. A replication study of GWAS findings in migraine identifies association in a Swedish case-control sample. BMC Med. Genet. 2014, 15, 38. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chow, S.-C. Sample size calculation for comparing proportions. Wiley Encycl. Clin. Trials 2007, 1–11. [Google Scholar]
- Visscher, P.M.; Hemani, G.; Vinkhuyzen, A.A.E.; Chen, G.B.; Lee, S.H.; Wray, N.R.; Goddard, M.E.; Yang, J. Statistical power to detect genetic (co)variance of complex traits using SNP data in unrelated samples. PLoS Genet. 2014, 10, e1004269. [Google Scholar] [CrossRef] [PubMed]
- Kajimura, S.; Saito, M. A new era in brown adipose tissue biology: Molecular control of brown fat development and energy homeostasis. Annu. Rev. Physiol. 2014, 76, 225–249. [Google Scholar] [CrossRef] [PubMed]
- Geerling, J.J.; Boon, M.R.; van der Zon, G.C.; van den Berg, S.A.A.; van den Hoek, A.M.; Lombès, M.; Princen, H.M.; Havekes, L.M.; Rensen, P.C.; Guigas, B. Metformin lowers plasma triglycerides by promoting VLDL-triglyceride clearance by brown adipose tissue in mice. Diabetes 2014, 63, 880–891. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Yang, X.; Lim, S.; Cao, Z.; Honek, J.; Lu, H.; Zhang, C.; Seki, T.; Hosaka, K.; Wahlberg, E.; et al. Cold exposure promotes atherosclerotic plaque growth and instability via UCP1-dependent lipolysis. Cell Metab. 2013, 18, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Lubkowska, A.; Banfi, G.; Dołegowska, B.; d’Eril, G.V.M.; Łuczak, J.; Barassi, A. Changes in lipid profile in response to three different protocols of whole-body cryostimulation treatments. Cryobiology 2010, 61, 22–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Total (173) | Obese (89) | Non-obese (84) | p Value | |
|---|---|---|---|---|
| Age (years) | 32 (±10) | 35.6 (±10) | 32.4 (±9) | 0.3 |
| Male % | 20.5% | 20% | 21% | 1.0 |
| BMI (kg/m2) | 29.5 (±7) | 35.5 (±5.5) | 25.3 (±3) | <0.0001 |
| Cholesterol (mmol/L) | 4.5 (±0.9) | 4.4 (±0.9) | 4.5 (±1.1) | 0.5 |
| HDL (mmol/L) | 1.2 (±0.3) | 1.1 (±0.28) | 1.3 (±0.4) | 0.0009 |
| Triglyceride (mmol/L) | 1.2 (±0.7) | 1.28 (±0.6) | 1.1 (±0.9) | 0.1 |
| LDL (mmol/L) | 3.1 (±0.8) | 3.1 (±0.9) | 3.1 (±0.8) | 0.15 |
| Systolic BP (mmHg) | 88 (±14) | 122 (±15) | 115 (±12) | 0.003 |
| Diastolic BP (mmHg) | 71 (±12) | 71 (±9) | 71 (±13) | 1.0 |
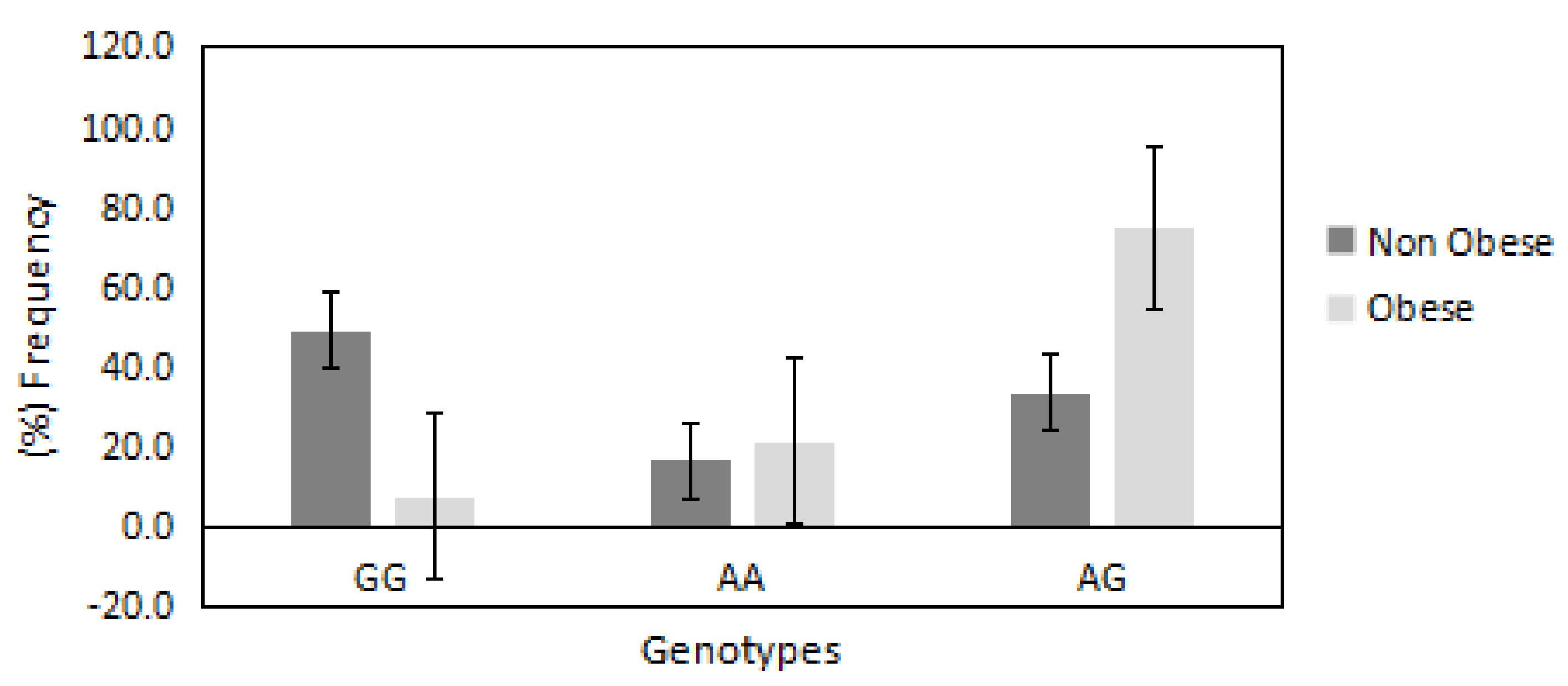
| PRDM16/rs2651899 | |||||
| Genotypes | (GG) | (AA) | (AG) | Allele (G) | Allele (A) |
| Obese (%) | 18 | 35 | 67 | 30 | 70 |
| Non-obese (%) | 82 | 65 | 24 | 66 | 34 |
| OR | 1 | 8.0 | 13.0 | 1 | 4.5 |
| 95% CI | Reference | 2.8308–22.7266 | 5.6623–33.8973 | Reference | 2.4980–8.2127 |
| p value | — | p = 0.0001 * | p < 0.0001 * | — | p < 0.0001 * |
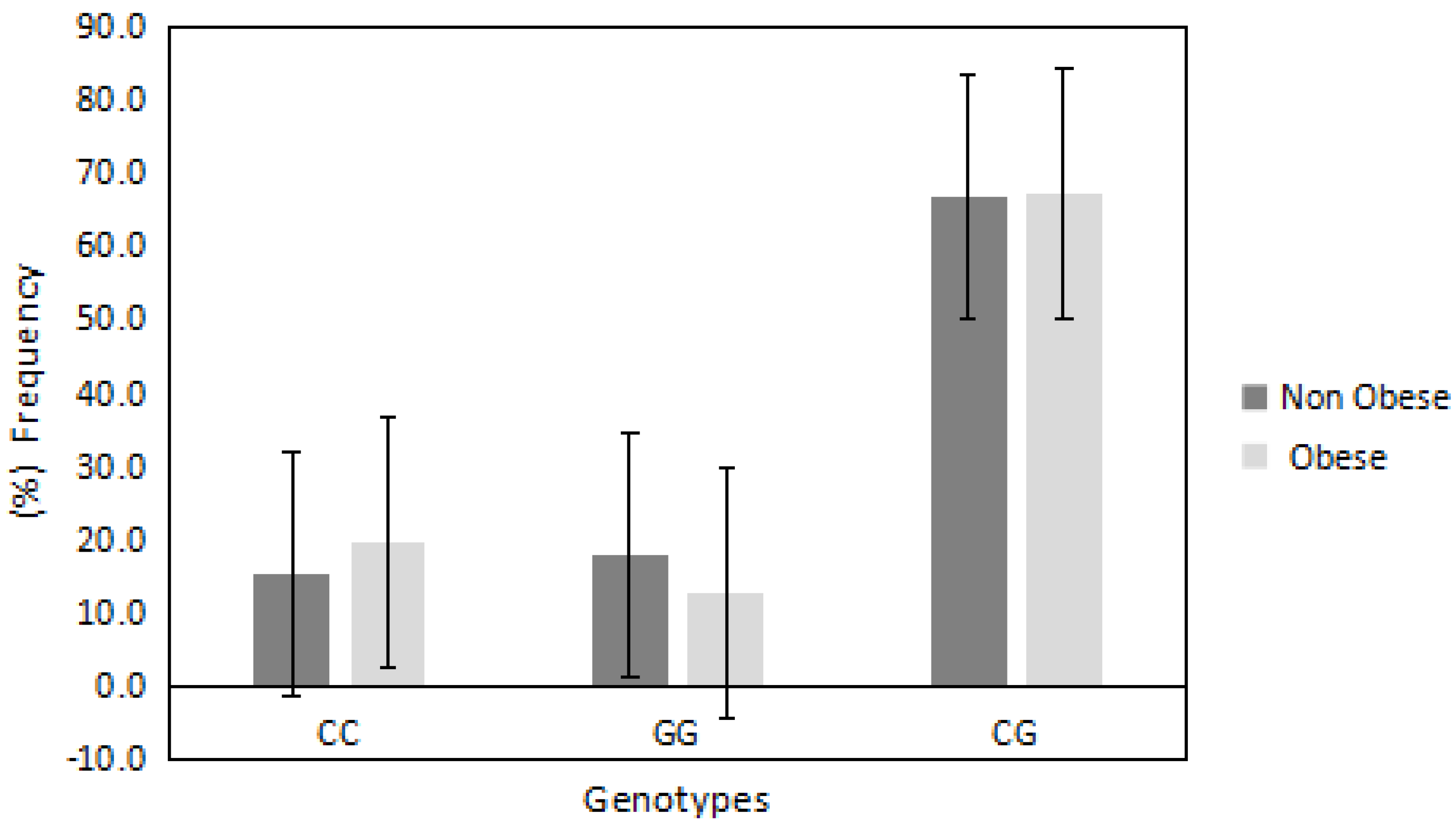
| PDE4D/rs295978 | |||||
| Genotypes | (CC) | (GG) | (CG) | Allele (C) | Allele (G) |
| Obese (%) | 62.5 | 48.2 | 56.7 | 51 | 49 |
| Non-obese (%) | 37.5 | 51.8 | 43.3 | 47 | 52 |
| OR | 1 | 0.5 | 0.7 | 1 | 1.5 |
| 95% CI | Reference | 0.1969–1.5767 | 0.3520–1.7490 | Reference | 0.6603–2.0083 |
| p value | — | p = 0.2 | p = 0.5 | — | 0.6 |
| PRDM16/rs2651899 | |||||||
| Genotypes | ANOVA | Tukey HSD | |||||
| GG MN (±SD) | AA MN (±SD) | AG MN (±SD) | F (p) | GG vs. AA Q (p) | GG vs. AG Q (p) | AA vs. AG Q (p) | |
| BMI (kg/m2) | 25 (±3) | 32 (±8) | 33 (±6) | 20 (<0.00001) * | 6.6 (0.001) * | 0.7 (0.8) | 8.9 (0.001) * |
| Cholesterol (mmol/L) | 4.6 (±0.7) | 3.8 (±0.8) | 4.6 (±0.9) | 5.8 (0.003) * | 3.9 (0.01) * | 4.6 (0.003) * | 0.1 (0.8) |
| HDL (mmol/L) | 1.3 (±0.3) | 1.0 (±0.3) | 1.1 (±0.3) | 4.9 (0.008) * | 4.2 (0.008) * | 1.7 (0.4) | 3.3 (0.05) * |
| LDL (mmol/L) | 3.0 (±0.6) | 2.5 (±0.8) | 3.2 (±0.4) | 3.8 (0.02) * | 2.8 (0.1) | 3.8 (0.01) * | 0.8 (0.8) |
| Triglycerides (mmol/L) | 0.9 (±0.3) | 1.0 (±0.3) | 1.4 (±0.4) | 5.8 (0.003) * | 1.0 (0.7) | 4.5 (0.004) * | 3.0 (0.08) |
| SBP (mmHg) | 119 (±10) | 117 (±9) | 120 (±11) | 0.5 (0.5) | 0.5 (0.8) | 0.2 (0.8) | 1.4 (0.5) |
| DBP (mmHg) | 70 (±10) | 71 (±9) | 69 (±10) | 0.2 (0.5) | 0.2 (0.8) | 0.7 (0.8) | 0.8 (0.8) |
| PDE4D/rs295978 | |||||||
| Genotypes | ANOVA | Tukey HSD | |||||
| CC MN (±SD) | GG MN (±SD) | CG MN (±SD) | F (p) | CC vs. GG Q (p) | CC vs. CG Q (p) | GG vs. CG Q (p) | |
| BMI (kg/m2) | 32 (±8) | 29 (±6) | 30 (±7) | 1.0 (0.3) | 2.1 (0.3) | 1.3 (0.6) | 1.3 (0.6) |
| Cholesterol (mmol/L) | 4.1 (±1) | 4.6 (±1.1) | 4.6 (±0.9) | 2.0 (0.1) | 2.1 (0.2) | 2.0 (0.1) | 0.1 (0.8) |
| HDL (mmol/L) | 1.0 (±0.3) | 1.2 (±0.3) | 1.0 (±0.2) | 3.3 (0.03) * | 3.2 (0.05) * | 0.2 (0.8) | 3.3 (0.05) * |
| LDL (mmol/L) | 2.8 (±0.6) | 3.1 (±0.8) | 3.1 (±0.4) | 0.9 (0.3) | 1.4 (0.5) | 1.9 (0.3) | 0.1 (0.8) |
| Triglycerides (mmol/L) | 1.2 (±0.5) | 1.1 (±0.3) | 1.2 (±0.4) | 0.6 (0.5) | 1.1 (0.6) | 1.2 (0.8) | 0.6 (0.4) |
| SBP (mmHg) | 120 (±11) | 112 (±9) | 119 (±10) | 0.5 (0.5) | 0.5 (0.8) | 0.2 (0.8) | 1.4 (0.5) |
| DBP (mmHg) | 72 (±8) | 69 (±6) | 70 (±9) | 0.2 (0.5) | 0.2 (0.8) | 0.7 (0.8) | 0.8 (0.8) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
AlAmrani, A.; AbdelKarim, M.; AlZoghaibi, M. PRDM16 Gene Polymorphism Is Associated with Obesity and Blood Lipids Profiles in Saudi Population. J. Clin. Med. 2018, 7, 141. https://doi.org/10.3390/jcm7060141
AlAmrani A, AbdelKarim M, AlZoghaibi M. PRDM16 Gene Polymorphism Is Associated with Obesity and Blood Lipids Profiles in Saudi Population. Journal of Clinical Medicine. 2018; 7(6):141. https://doi.org/10.3390/jcm7060141
Chicago/Turabian StyleAlAmrani, Aishah, Mouaadh AbdelKarim, and Mohammed AlZoghaibi. 2018. "PRDM16 Gene Polymorphism Is Associated with Obesity and Blood Lipids Profiles in Saudi Population" Journal of Clinical Medicine 7, no. 6: 141. https://doi.org/10.3390/jcm7060141
APA StyleAlAmrani, A., AbdelKarim, M., & AlZoghaibi, M. (2018). PRDM16 Gene Polymorphism Is Associated with Obesity and Blood Lipids Profiles in Saudi Population. Journal of Clinical Medicine, 7(6), 141. https://doi.org/10.3390/jcm7060141