Can the Fact That Myelin Proteins Are Old and Break down Explain the Origin of Multiple Sclerosis in Some People?

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Long-Lived Proteins and Autoimmunity
3. Long-Lived Proteins and Multiple Sclerosis
4. Myelin Basic Protein and Age
5. Myelin Basic Protein and MS
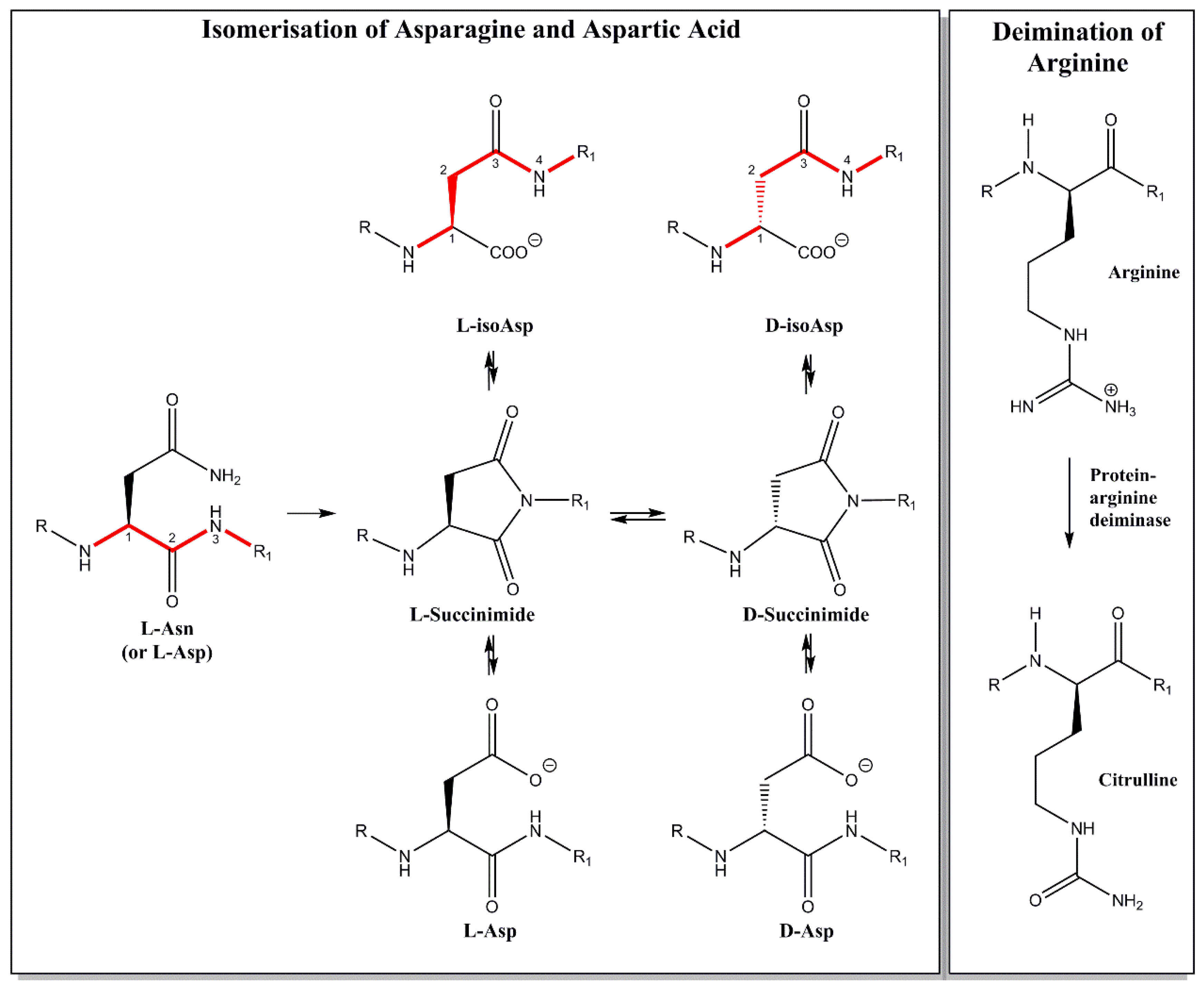
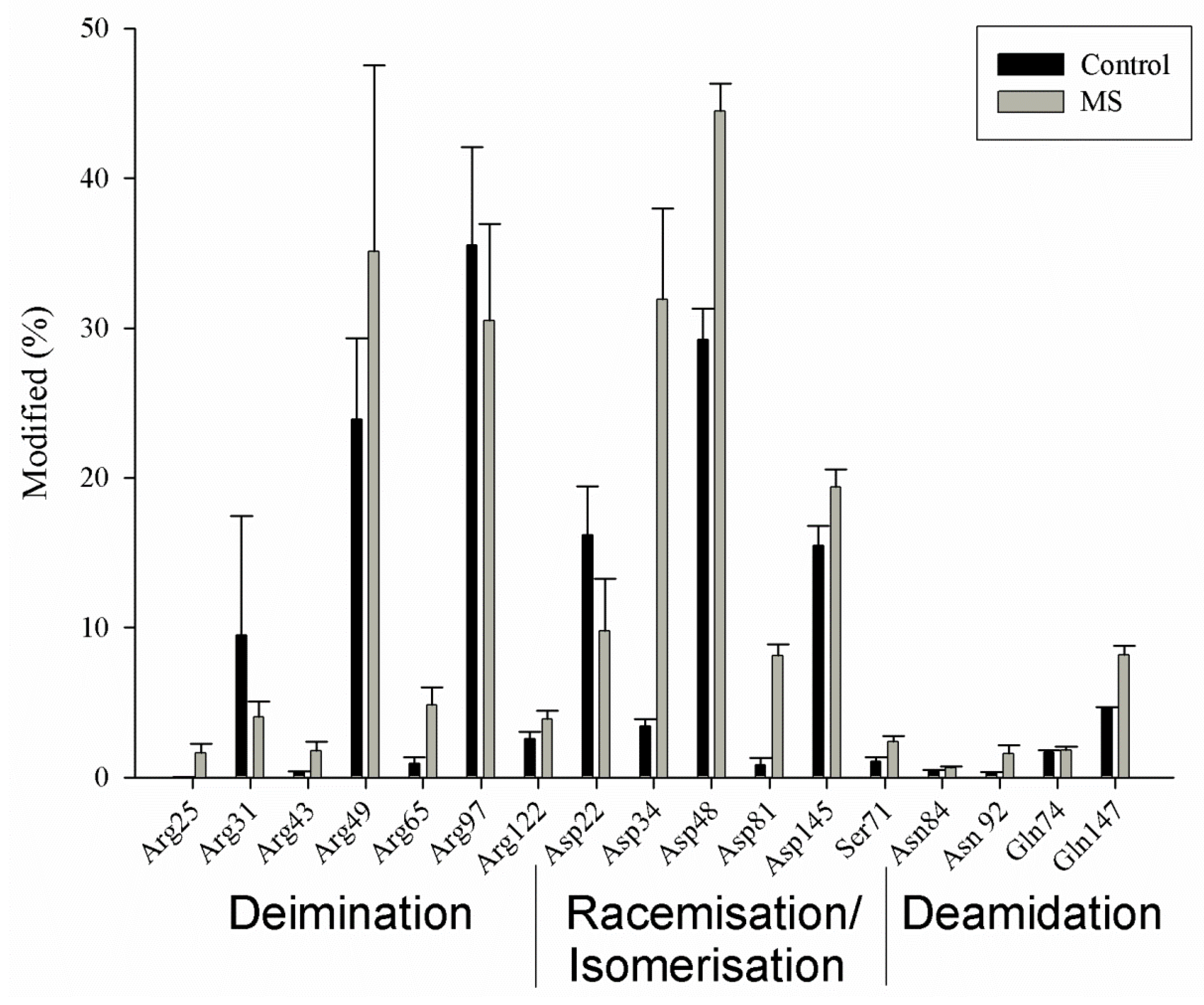
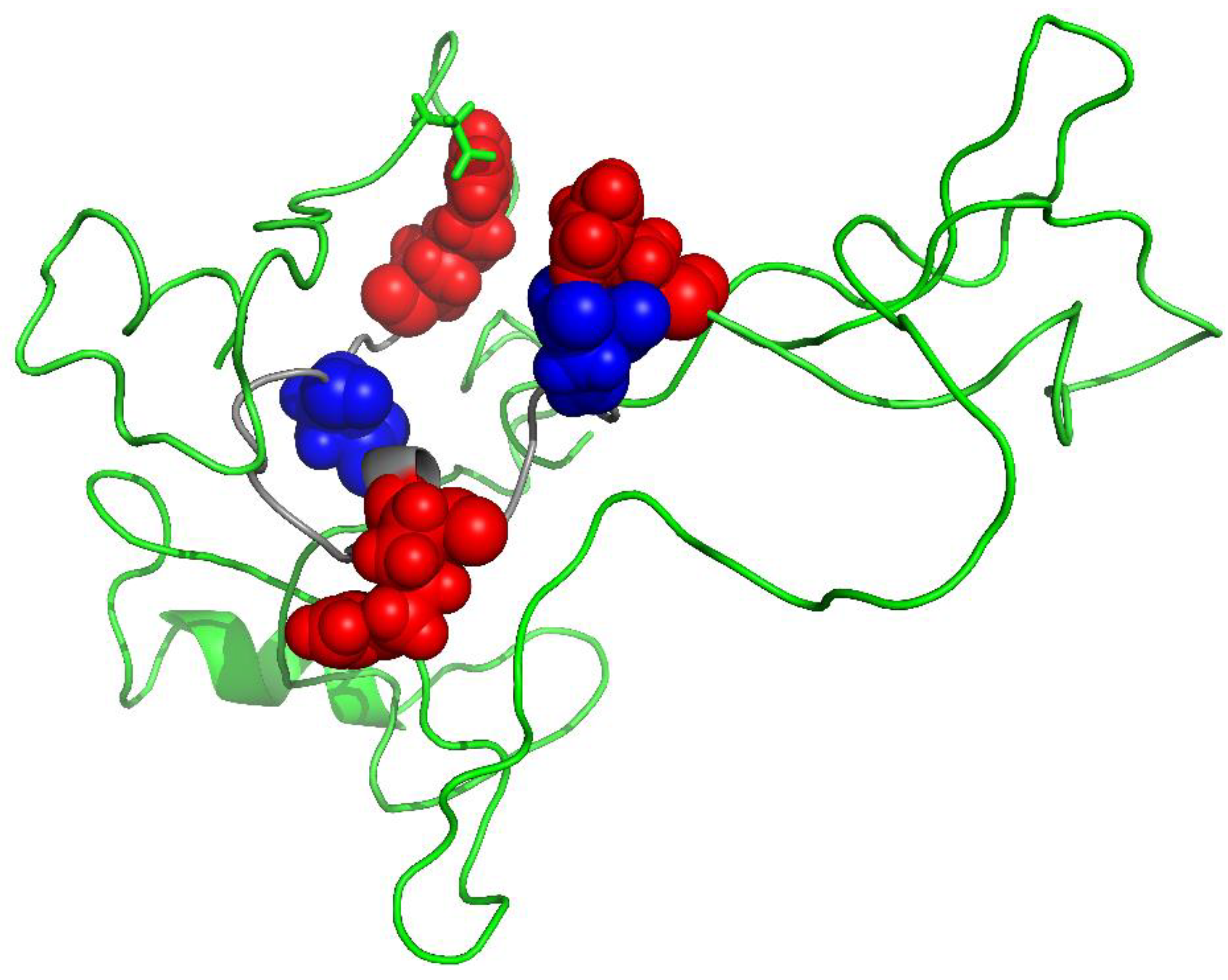
6. Myelin Basic Protein from MS Patients Is Different
7. Considerations If MS Results from Age-Related Changes to MBP
8. Gender, MS, and Age-Related Changes to MBP
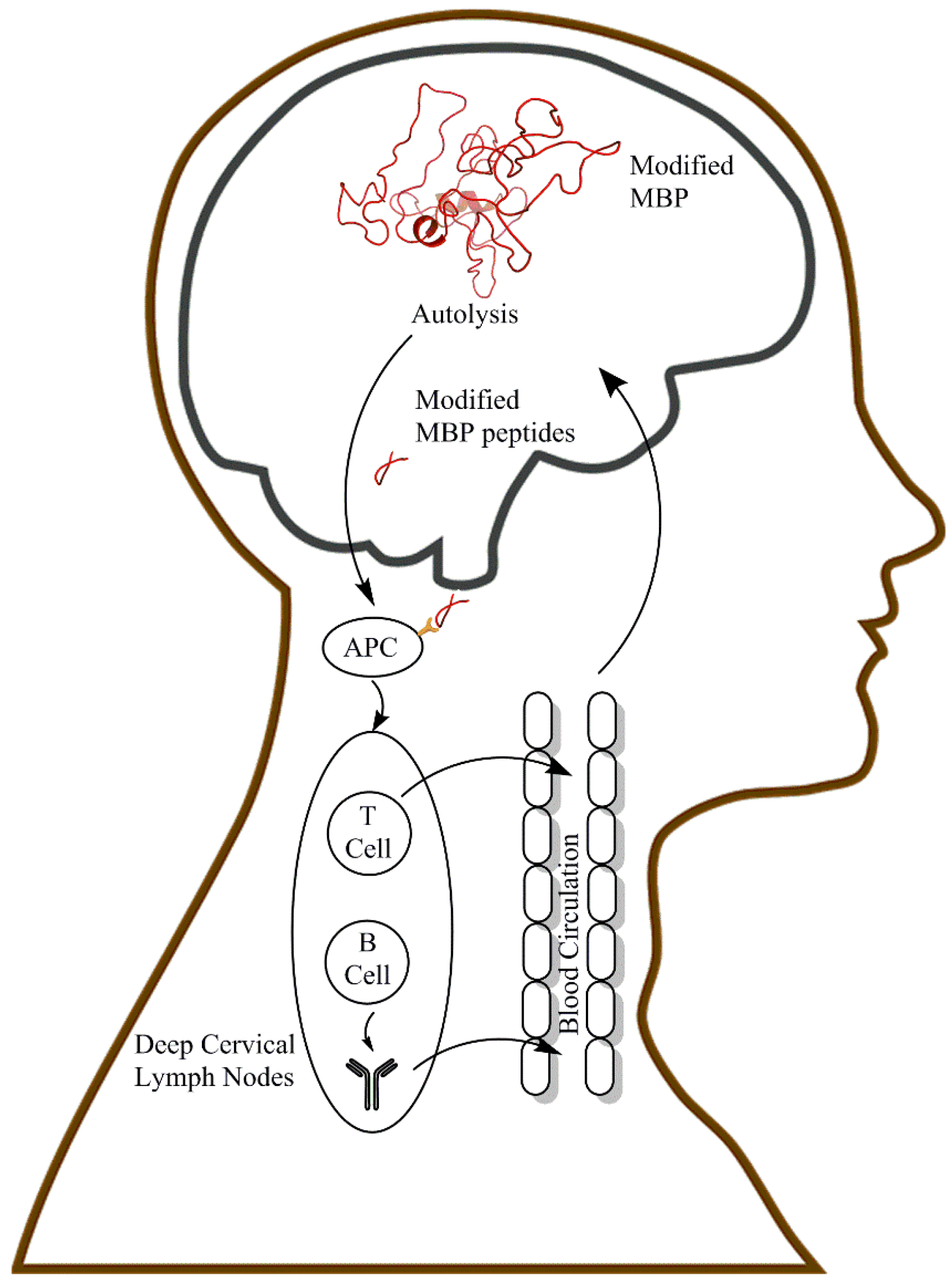
9. Linkage between the CNS and the Periphery in MS
10. Autoantibodies and MS
11. Cellular Aspects of MS
12. A New Theory for MS
13. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Baranzini, S.E. Revealing the genetic basis of multiple sclerosis: Are we there yet? Curr. Opin. Genet. Dev. 2011, 21, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Patsopoulos, N.; Baranzini, S.E.; Santaniello, A.; Shoostari, P.; Cotsapas, C.; Wong, G.; Beecham, A.H.; James, T.; Replogle, J.; Vlachos, I.; et al. The multiple sclerosis genomic map: Role of peripheral immune cells and resident microglia in susceptibility. bioRxiv 2017, 1, 143933. [Google Scholar]
- Verdu, E.F.; Caminero, A. How infection can incite sensitivity to food. Science 2017, 356, 29–30. [Google Scholar] [CrossRef] [PubMed]
- Erny, D.; Prinz, M. Microbiology: Gut microbes augment neurodegeneration. Nature 2017, 544, 304–305. [Google Scholar] [CrossRef] [PubMed]
- Rothhammer, V.; Borucki, D.M.; Tjon, E.C.; Takenaka, M.C.; Chao, C.C.; Ardura-Fabregat, A.; de Lima, K.A.; Gutiérrez-Vázquez, C.; Hewson, P.; Staszewski, O.; et al. Microglial control of astrocytes in response to microbial metabolites. Nature 2018, 557, 724–728. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.; Diering, G.H.; Na, C.H.; Nirujogi, R.S.; Bachman, J.L.; Pandey, A.; Huganir, R.L. Identification of long-lived synaptic proteins by proteomic analysis of synaptosome protein turnover. Proc. Natl. Acad. Sci. USA 2018, 115, E3827–E3836. [Google Scholar] [CrossRef] [PubMed]
- Toyama, B.H.; Savas, J.N.; Park, S.K.; Harris, M.S.; Ingolia, N.T.; Yates, J.R.; Hetzer, M.W. Identification of long-lived proteins reveals exceptional stability of essential cellular structures. Cell 2013, 154, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Lynnerup, N.; Kjeldsen, H.; Heegaard, S.; Jacobsen, C.; Heinemeier, J. Radiocarbon dating of the human eye lens crystallines reveal proteins without carbon turnover throughout life. PLoS ONE 2008, 3, e1529. [Google Scholar] [CrossRef] [PubMed]
- Truscott, R.J.; Schey, K.L.; Friedrich, M.G. Old proteins in man: A field in its infancy. Trends Biochem. Sci. 2016, 41, 654–664. [Google Scholar] [CrossRef] [PubMed]
- Rowley, M.; Buchanan, H.; Mackay, I. Recipocal change with age in antibody to extrinsic and intrinsic antigens. Lancet 1968, 292, 24–26. [Google Scholar] [CrossRef]
- Doyle, H.A.; Aswad, D.W.; Mamula, M.J. Autoimmunity to isomerized histone h2b in systemic lupus erythematosus. Autoimmunity 2012, 46, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Doyle, H.A.; Zhou, J.; Wolff, M.J.; Harvey, B.P.; Roman, R.M.; Gee, R.J.; Koski, R.A.; Mamula, M.J. Isoaspartyl post-translational modification triggers anti-tumor t and b lymphocyte immunity. J. Biol. Chem. 2006, 281, 32676–32683. [Google Scholar] [CrossRef] [PubMed]
- Geiger, T.; Clarke, S. Deamidation, isomerization, and racemization at asparaginyl and aspartyl residues in peptides. Succinimide-linked reactions that contribute to protein degradation. J. Biol. Chem. 1987, 262, 785–794. [Google Scholar] [PubMed]
- Friedrich, M.G.; Wang, Z.; Oakley, A.J.; Schey, K.L.; Truscott, R.J. Hotspots of age-related protein degradation. The importance of neighbouring residues for the formation of non-disulfide crosslinks derived from cysteine. Biochem. J. 2017, 7, BC20170268. [Google Scholar]
- Wang, Z.; Lyons, B.; Truscott, R.J.; Schey, K.L. Human protein aging: Modification and crosslinking through dehydroalanine and dehydrobutyrine intermediates. Aging Cell 2014, 13, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Wilmarth, P.A.; Tanner, S.; Dasari, S.; Nagalla, S.R.; Riviere, M.A.; Bafna, V.; Pevzner, P.A.; David, L.L. Age-related changes in human crystallins determined from comparative analysis of post-translational modifications in young and aged lens: Does deamidation contribute to crystallin insolubility? J. Proteome Res. 2006, 5, 2554–2566. [Google Scholar] [CrossRef] [PubMed]
- Su, S.P.; Lyons, B.; Friedrich, M.; McArthur, J.D.; Song, X.; Xavier, D.; Truscott, R.J.; Aquilina, J.A. Molecular signatures of long-lived proteins: Autolytic cleavage adjacent to serine residues. Aging Cell 2012, 11, 1125–1127. [Google Scholar] [CrossRef] [PubMed]
- Hooi, M.; Truscott, R. Racemisation and human cataract. D-ser, d-asp/asn and d-thr are higher in the lifelong proteins of cataract lenses than in age-matched normal lenses. Age 2011, 33, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Hooi, M.Y.; Raftery, M.J.; Truscott, R.J. Racemization of two proteins over our lifespan: Deamidation of asparagine 76 in γs crystallin is greater in cataract than in normal lenses across the age range. Investig. Ophthalmol. Visual Sci. 2012, 53, 3554–3561. [Google Scholar] [CrossRef] [PubMed]
- Hooi, M.Y.; Raftery, M.J.; Truscott, R.J. Accelerated aging of asp 58 in αa crystallin and human cataract formation. Exp. Eye Res. 2013, 106, 34–39. [Google Scholar] [CrossRef] [PubMed]
- DeBruin, L.S.; Haines, J.D.; Wellhauser, L.A.; Radeva, G.; Schonmann, V.; Bienzle, D.; Harauz, G. Developmental partitioning of myelin basic protein into membrane microdomains. J. Neurosci. Res. 2005, 80, 211–225. [Google Scholar] [CrossRef] [PubMed]
- Sarg, B.; Faserl, K.; Lindner, H.H. Identification of novel site-specific alterations in the modification level of myelin basic protein isolated from mouse brain at different ages using capillary electrophoresis–mass spectrometry. Proteomics 2017, 17, 1700269. [Google Scholar] [CrossRef] [PubMed]
- Vassall, K.A.; Bamm, V.V.; Harauz, G. Myelstones: The executive roles of myelin basic protein in myelin assembly and destabilization in multiple sclerosis. Biochem. J. 2015, 472, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Dendrou, C.A.; Fugger, L.; Friese, M.A. Immunopathology of multiple sclerosis. Nat. Rev. Immunol. 2015, 15, 545–558. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Mastronardi, F.G.; Wood, D.D.; Lubman, D.M.; Zand, R.; Moscarello, M.A. Multiple sclerosis: An important role for post-translational modifications of myelin basic protein in pathogenesis. Mol. Cell. Proteom. 2003, 2, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Moscarello, M.A.; Wood, D.D.; Ackerley, C.; Boulias, C. Myelin in multiple sclerosis is developmentally immature. J. Clin. Investig. 1994, 94, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Steelman, A.J. Infection as an environmental trigger of multiple sclerosis disease exacerbation. Front. Immunol. 2015, 6, 520. [Google Scholar] [CrossRef] [PubMed]
- Hauser, S.L.; Waubant, E.; Arnold, D.L.; Vollmer, T.; Antel, J.; Fox, R.J.; Bar-Or, A.; Panzara, M.; Sarkar, N.; Agarwal, S.; et al. B-cell depletion with rituximab in relapsing–remitting multiple sclerosis. N. Engl. J. Med. 2008, 358, 676–688. [Google Scholar] [CrossRef] [PubMed]
- Kinnunen, T.; Chamberlain, N.; Morbach, H.; Choi, J.; Kim, S.; Craft, J.; Mayer, L.; Cancrini, C.; Passerini, L.; Bacchetta, R.; et al. Accumulation of peripheral autoreactive b cells in the absence of functional human regulatory t cells. Blood 2013, 121, 1595–1603. [Google Scholar] [CrossRef] [PubMed]
- von Büdingen, H.C.; Bar-Or, A.; Zamvil, S.S. B cells in multiple sclerosis: Connecting the dots. Curr. Opin. Immunol. 2011, 23, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Viglietta, V.; Baecher-Allan, C.; Weiner, H.L.; Hafler, D.A. Loss of functional suppression by CD4+ CD25+ regulatory T cells in patients with multiple sclerosis. J. Exp. Med. 2004, 199, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Liu, Y.; Liu, B.; Tan, H.; Lu, H.; Wang, H.; Yan, H. Treatment of surgical brain injury by immune tolerance induced by intrathymic and hepatic portal vein injection of brain antigens. Sci. Rep. 2016, 6, 32030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhagen, J.; Wegner, A.; Wraith, D.C. Extra-thymically induced t regulatory cell subsets: The optimal target for antigen-specific immunotherapy. Immunology 2015, 145, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Gallart-Palau, X.; Lee, B.S.T.; Adav, S.S.; Qian, J.; Serra, A.; Park, J.E.; Lai, M.K.P.; Chen, C.P.; Kalaria, R.N.; Sze, S.K. Gender differences in white matter pathology and mitochondrial dysfunction in alzheimer’s disease with cerebrovascular disease. Mol. Brain 2016, 9, 27. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M. Animal models of multiple sclerosis: The good, the bad and the bottom line. Nat. Neurosci. 2012, 15, 1074–1077. [Google Scholar] [CrossRef] [PubMed]
- Ellwardt, E.; Zipp, F. Molecular mechanisms linking neuroinflammation and neurodegeneration in ms. Exp. Neurol. 2014, 262, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Louveau, A.; Harris, T.H.; Kipnis, J. Revisiting the concept of cns immune privilege. Trends Immunol. 2015, 36, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Cserr, H.F.; Harling-Berg, C.J.; Knopf, P.M. Drainage of brain extracellular fluid into blood and deep cervical lymph and its immunological significance. Brain Pathol. 1992, 2, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Lyons, B.; Kwan, A.H.; Truscott, R.J. Spontaneous cleavage of proteins at serine and threonine is facilitated by zinc. Aging Cell 2016, 15, 237–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korn, T.; Kallies, A. T cell responses in the central nervous system. Nat. Rev. Immunol. 2017, 17, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Locatelli, G.; Wortge, S.; Buch, T.; Ingold, B.; Frommer, F.; Sobottka, B.; Kruger, M.; Karram, K.; Buhlmann, C.; Bechmann, I.; et al. Primary oligodendrocyte death does not elicit anti-cns immunity. Nat. Neurosci. 2012, 15, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Traka, M.; Podojil, J.R.; McCarthy, D.P.; Miller, S.D.; Popko, B. Oligodendrocyte death results in immune-mediated cns demyelination. Nat. Neurosci. 2016, 19, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Berger, T.; Rubner, P.; Schautzer, F.; Egg, R.; Ulmer, H.; Mayringer, I.; Dilitz, E.; Deisenhammer, F.; Reindl, M. Antimyelin antibodies as a predictor of clinically definite multiple sclerosis after a first demyelinating event. N. Engl. J. Med. 2003, 349, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Kuhle, J.; Pohl, C.; Mehling, M.; Edan, G.; Freedman, M.S.; Hartung, H.-P.; Polman, C.H.; Miller, D.H.; Montalban, X.; Barkhof, F.; et al. Lack of association between antimyelin antibodies and progression to multiple sclerosis. N. Engl. J. Med. 2007, 356, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Nemazee, D. Mechanisms of central tolerance for b cells. Nat. Rev. Immunol. 2017, 17, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Kinnunen, T.; Chamberlain, N.; Morbach, H.; Cantaert, T.; Lynch, M.; Preston-Hurlburt, P.; Herold, K.C.; Hafler, D.A.; O’Connor, K.C.; Meffre, E. Specific peripheral b cell tolerance defects in patients with multiple sclerosis. J. Clin. Investig. 2013, 123, 2737–2741. [Google Scholar] [CrossRef] [PubMed]
- Doyle, H.A.; Mamula, M.J. Autoantigenesis: The evolution of protein modifications in autoimmune disease. Curr. Opin. Immunol. 2012, 24, 112–118. [Google Scholar] [CrossRef] [PubMed]




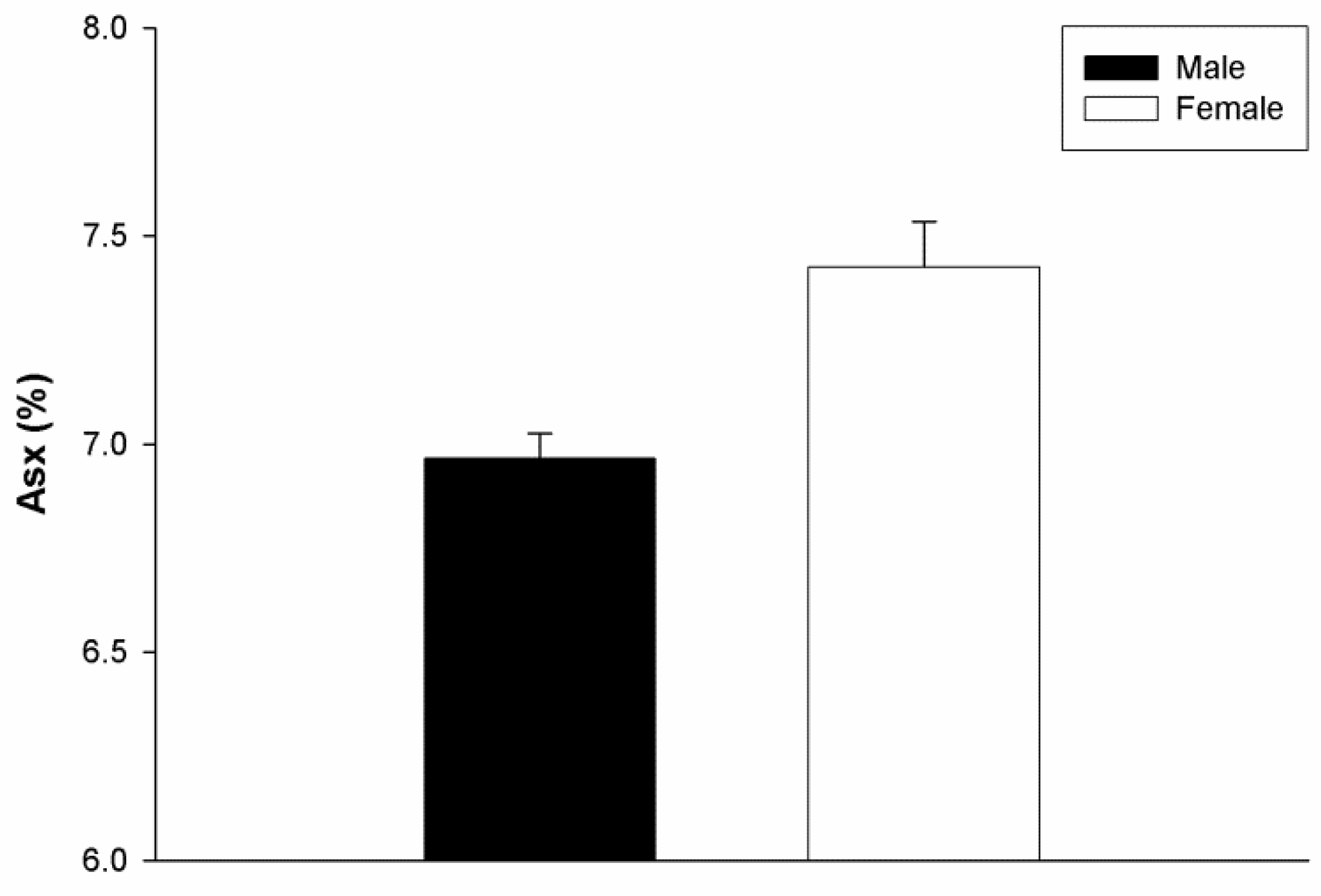
 ) and female (
) and female (  ) subjects. Elevated levels of d-Asx were found in females by comparison to males (p = 0.029, Mann–Whitney U test). Male n = 4, female n = 4, age range 51–87 years. Mean ± SEM.
) and female ( ) subjects. Elevated levels of d-Asx were found in females by comparison to males (p = 0.029, Mann–Whitney U test). Male n = 4, female n = 4, age range 51–87 years. Mean ± SEM.
) subjects. Elevated levels of d-Asx were found in females by comparison to males (p = 0.029, Mann–Whitney U test). Male n = 4, female n = 4, age range 51–87 years. Mean ± SEM.
) and female ( ) subjects. Elevated levels of d-Asx were found in females by comparison to males (p = 0.029, Mann–Whitney U test). Male n = 4, female n = 4, age range 51–87 years. Mean ± SEM.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Truscott, R.J.W.; Friedrich, M.G. Can the Fact That Myelin Proteins Are Old and Break down Explain the Origin of Multiple Sclerosis in Some People? J. Clin. Med. 2018, 7, 281. https://doi.org/10.3390/jcm7090281
Truscott RJW, Friedrich MG. Can the Fact That Myelin Proteins Are Old and Break down Explain the Origin of Multiple Sclerosis in Some People? Journal of Clinical Medicine. 2018; 7(9):281. https://doi.org/10.3390/jcm7090281
Chicago/Turabian StyleTruscott, Roger J. W., and Michael G. Friedrich. 2018. "Can the Fact That Myelin Proteins Are Old and Break down Explain the Origin of Multiple Sclerosis in Some People?" Journal of Clinical Medicine 7, no. 9: 281. https://doi.org/10.3390/jcm7090281
APA StyleTruscott, R. J. W., & Friedrich, M. G. (2018). Can the Fact That Myelin Proteins Are Old and Break down Explain the Origin of Multiple Sclerosis in Some People? Journal of Clinical Medicine, 7(9), 281. https://doi.org/10.3390/jcm7090281