Elevated Hedgehog-Interacting Protein Levels in Subjects with Prediabetes and Type 2 Diabetes

 , ,
, ,
Abstract
:1. Introduction
2. Methods
2.1. Study Participants
2.2. Statistics
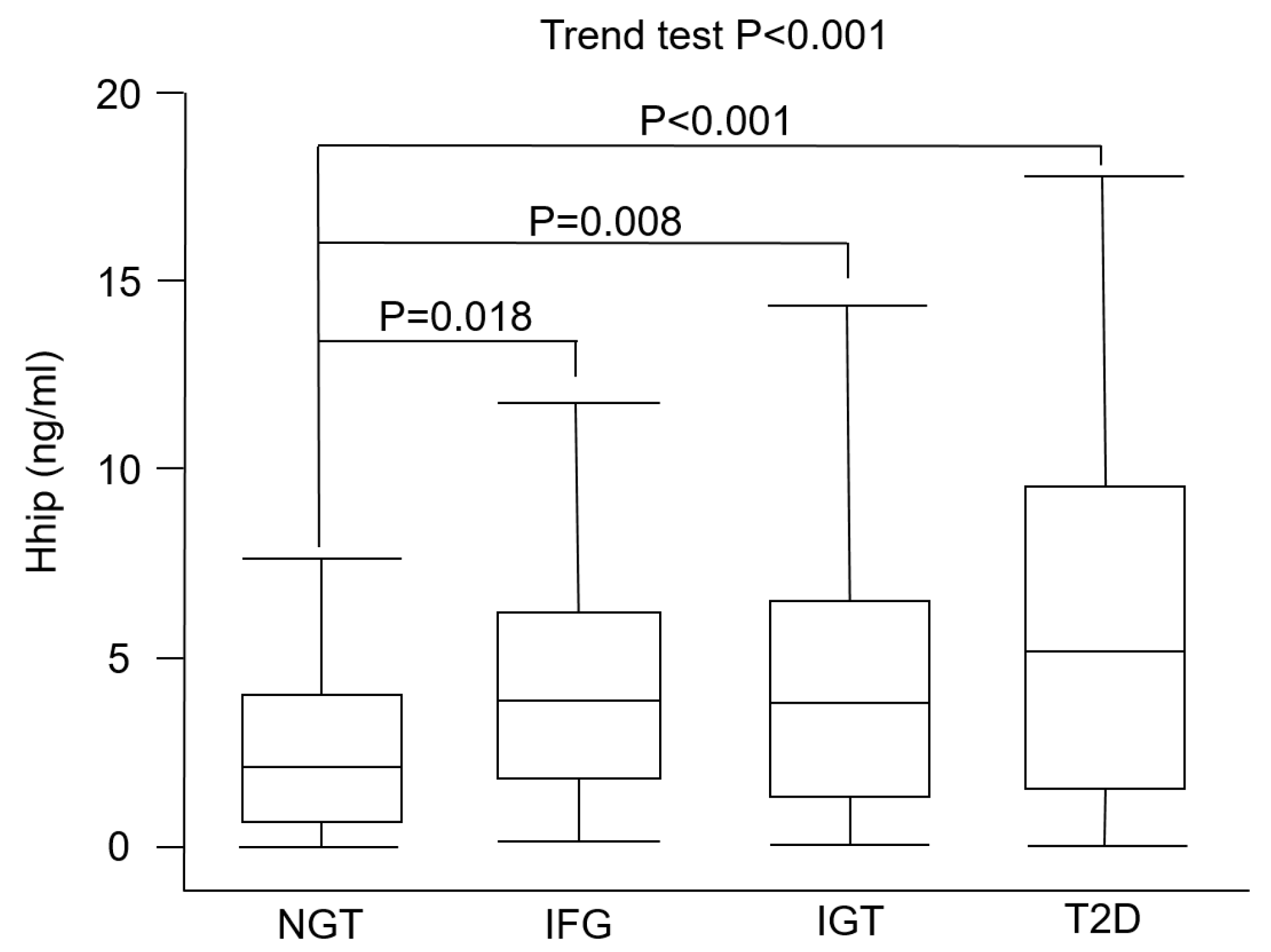
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IDF Diabetes Atlas Eighth Edition 2017. Available online: http://www.diabetesatlas.org (accessed on 14 August 2019).
- Diabetes.co.uk. Diabetes and Cancer. Available online: https://www.diabetes.co.uk/diabetes-complications/diabetes-and-cancer.html (accessed on 14 August 2019).
- American Diabetes Association. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2019. Diabetes Care 2019, 42, S13–S28. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, W.A.; Orloski, L.; Wyne, K. Prediabetes: The importance of early identification and intervention. Postgrad Med. 2010, 122, 129–143. [Google Scholar] [CrossRef] [PubMed]
- Davidson, M.B. Counterpoint: The oral glucose tolerance test is superfluous. Diabetes Care. 2002, 25, 1883–1885. [Google Scholar] [CrossRef] [PubMed]
- Chuang, P.T.; McMahon, A.P. Vertebrate Hedgehog signalling modulated by induction of a Hedgehog-binding protein. Nature 1999, 397, 617–621. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.P.; Chang, S.Y.; Liao, M.C.; Lo, C.S.; Chenier, I.; Luo, H.; Chiasson, J.L.; Ingelfinger, J.R.; Chan, J.S.D.; Zhang, S.L. Hedgehog Interacting Protein Promotes Fibrosis and Apoptosis in Glomerular Endothelial Cells in Murine Diabetes. Sci. Rep. 2018, 8, 5958. [Google Scholar] [CrossRef]
- Agrawal, V.; Kim, D.Y.; Kwon, Y.-G. Hhip regulates tumor-stroma-mediated upregulation of tumor angiogenesis. Exp. Mol. Med. 2017, 49, e289. [Google Scholar] [CrossRef]
- Olsen, C.L.; Hsu, P.P.; Glienke, J.; Rubanyi, G.M.; Brooks, A.R. Hedgehog-interacting protein is highly expressed in endothelial cells but down-regulated during angiogenesis and in several human tumors. BMC Cancer 2004, 4, 43. [Google Scholar] [CrossRef]
- Onishi, H.; Katano, M. Hedgehog signaling pathway as a therapeutic target in various types of cancer. Cancer Sci. 2011, 102, 1756–1760. [Google Scholar] [CrossRef]
- Skoda, A.M.; Simovic, D.; Karin, V.; Kardum, V.; Vranic, S.; Serman, L. The role of the Hedgehog signaling pathway in cancer: A comprehensive review. Bosn. J. Basic Med. Sci. 2018, 18, 8–20. [Google Scholar] [CrossRef]
- Sun, H.; Ni, S.j.; Ye, M.; Weng, W.; Zhang, Q.; Zhang, M.; Tan, C.; Wang, L.; Huang, D.; Du, X.; et al. Hedgehog Interacting Protein 1 is a Prognostic Marker and Suppresses Cell Metastasis in Gastric Cancer. J. Cancer. 2018, 9, 4642–4649. [Google Scholar] [CrossRef]
- Thayer, S.P.; di Magliano, M.P.; Heiser, P.W.; Nielsen, C.M.; Roberts, D.J.; Lauwers, G.Y.; Qi, Y.P.; Gysin, S.; Fernández-del Castillo, C.; Yajnik, V.; et al. Hedgehog is an early and late mediator of pancreatic cancer tumorigenesis. Nature 2003, 425, 851–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Interactive Database of Gene Expression and Diabetes-related Clinical Phenotypes. Available online: http://diabetes.wisc.edu (accessed on 14 August 2019).
- Nchienzia, H.; Liao, M.C.; Zhao, X.P.; Chang, S.Y.; Lo, C.S.; Chenier, I.; Ingelfinger, J.R.; Chan, J.S.D.; Zhang, S.L. Hedgehog Interacting Protein (Hhip) Regulates Insulin Secretion in Mice Fed High Fat Diets. Sci. Rep. 2019, 9, 11183. [Google Scholar] [CrossRef] [PubMed]
- Apelqvist, Å.; Ahlgren, U.; Edlund, H. Sonic hedgehog directs specialised mesoderm differentiation in the intestine and pancreas. Curr. Biol. 1997, 7, 801–804. [Google Scholar] [CrossRef] [Green Version]
- Hebrok, M.; Kim, S.K.; Melton, D.A. Notochord repression of endodermal Sonic hedgehog permits pancreas development. Genes Dev. 1998, 12, 1705–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawahira, H.; Ma, N.H.; Tzanakakis, E.S.; McMahon, A.P.; Chuang, P.T.; Hebrok, M. Combined activities of hedgehog signaling inhibitors regulate pancreas development. Development 2003, 130, 4871–4879. [Google Scholar] [CrossRef] [Green Version]
- Landsman, L.; Parent, A.; Hebrok, M. Elevated Hedgehog/Gli signaling causes beta-cell dedifferentiation in mice. Proc. Natl. Acad. Sci. USA 2011, 108, 17010–17015. [Google Scholar] [CrossRef]
- Suzuki, T.; Dai, P.; Hatakeyama, T.; Harada, Y.; Tanaka, H.; Yoshimura, N.; Takamatsu, T. TGF-β Signaling Regulates Pancreatic β-Cell Proliferation through Control of Cell Cycle Regulator p27 Expression. Acta Histochem. Cytochem. 2013, 46, 51–58. [Google Scholar] [CrossRef]
- Meier, J.J.; Breuer, T.G.; Bonadonna, R.C.; Tannapfel, A.; Uhl, W.; Schmidt, W.E.; Schrader, H.; Menge, B.A. Pancreatic diabetes manifests when beta cell area declines by approximately 65% in humans. Diabetologia 2012, 55, 1346–1354. [Google Scholar] [CrossRef] [Green Version]
- Jonas, J.C.; Sharma, A.; Hasenkamp, W.; Ilkova, H.; Patanè, G.; Laybutt, R.; Bonner-Weir, S.; Weir, G.C. Chronic hyperglycemia triggers loss of pancreatic beta cell differentiation in an animal model of diabetes. J. Biol. Chem. 1999, 274, 14112–14121. [Google Scholar] [CrossRef]
- Accili, D. Insulin Action Research and the Future of Diabetes Treatment: The 2017 Banting Medal for Scientific Achievement Lecture. Diabetes 2018, 67, 1701–1709. [Google Scholar] [CrossRef] [Green Version]
- Ochi, H.; Pearson, B.J.; Chuang, P.T.; Hammerschmidt, M.; Westerfield, M. Hhip regulates zebrafish muscle development by both sequestering Hedgehog and modulating localization of Smoothened. Dev. Biol. 2006, 297, 127–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, H.; Li, J.; Shi, S.; Zhang, L.; Xiang, A.; Shi, X.; Yang, G.; Chu, G. Hhip inhibits proliferation and promotes differentiation of adipocytes through suppressing hedgehog signaling pathway. Biochem. Biophys. Res. Commun. 2019, 514, 148–156. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| NGT | IFG | IGT | NDD | p Value | |
|---|---|---|---|---|---|
| N | 75 | 66 | 86 | 87 | |
| Age (years) | 61.5 ± 12.4 | 62.4 ± 12.0 | 62.4 ± 11.9 | 62.7 ± 11.6 | NS |
| Male (%) | 50.7 | 65.2 | 55.8 | 57.5 | NS |
| Hhip (ng/mL) | 2.45 ± 2.12 | 4.40 ± 3.22 | 4.44 ± 3.64 | 6.31 ± 5.35 | <0.001 |
| BMI (kg/m2) | 22.3 ± 2.7 | 23.6 ± 2.8 | 23.4 ± 3.0 | 23.3 ± 3.2 | 0.049 |
| SBP (mmHg) | 121.4 ± 17.2 | 126.9 ± 17.2 | 128.7 ± 18.2 | 131.9 ± 19.8 | 0.003 |
| DBP (mmHg) | 70.4 ± 9.9 | 73.4 ± 10.2 | 73.7 ± 10.7 | 75.4 ± 10.9 | 0.025 |
| FPG (mmol/L) | 85.8 ± 7.3 | 104.2 ± 4.8 | 88.5 ± 11.6 | 138.4 ± 61.6 | <0.001 |
| Post-load 2-h glucose (mmol/L) | 97.0 ± 24.2 | 107.0 ± 22.7 | 161.7 ± 16.7 | 262.1 ± 87.5 | <0.001 |
| A1C (%) | 5.7 ± 0.3 | 5.8 ± 0.4 | 5.8 ± 0.3 | 7.3 ± 2.0 | <0.001 |
| AST (U/L) | 26.6 ± 8.0 | 27.1 ± 14.0 | 24.7 ± 7.9 | 30.7 ± 43.6 | NS |
| ALT (U/L) | 23.5 ± 10.5 | 27.5 ± 21.1 | 22.5 ± 10.7 | 32.7 ± 54.9 | NS |
| hsCRP (mg/L) | 1.9 ± 3.0 | 3.0 ± 5.5 | 3.0 ± 5.3 | 5.9 ± 10.0 | 0.001 |
| Total cholesterol (mmol/L) | 197.1 ± 36.8 | 205.8 ± 31.6 | 196.2 ± 36.3 | 211.4 ± 48.7 | 0.038 |
| Triglyceride (mmol/L) * | 93.9 ± 38.1 | 119.0 ± 67.5 | 113.4 ± 60.2 | 131.3 ± 81.8 | 0.006 |
| HDL-C (mmol/L) | 60.5 ± 19.2 | 53.5 ± 15.1 | 53.5 ± 14.5 | 52.6 ± 13.8 | 0.006 |
| LDL-C (mmol/L) | 117.8 ± 35.7 | 128.5 ± 29.7 | 120.0 ± 31.9 | 132.5 ± 39.5 | 0.021 |
| Variable | Model 1 | Model 2 | Model 3 | |||
|---|---|---|---|---|---|---|
| β (95% CI) | p | β (95% CI) | p | β (95% CI) | p | |
| Age | −0.015 (−0.043, 0.032) | NS | −0.027 (−0.047, 0.028) | NS | −0.085 (−0.072, 0.015) | NS |
| Sex | 0.171 (0.513, 2.298) | 0.002 | 0.173 (0.519, 2.321) | 0.002 | 0.124 (0.059, 1.938) | 0.037 |
| A1C (%) | 0.175 (0.207, 0.894) | 0.002 | ||||
| FPG (mmol/L) | 0.120 (0.001, 0.024) | 0.032 | ||||
| IFG vs. NGT | 0.236 (1.086, 3.864) | 0.001 | ||||
| IGT vs. NGT | 0.214 (0.690, 3.103) | 0.002 | ||||
| NDD vs. NGT | 0.404 (2.305, 4.830) | <0.001 | ||||
| Insulin (mIU/L) | −0.143 (−0.397, 0.045) | 0.014 | ||||
| hsCRP (mg/L) | 0.067 (−0.027, 0.106) | NS | ||||
| BMI (kg/m2) | 0.003 (−0.160, 0.169) | NS | ||||
| SBP (mmHg) | −0.004 (−0.027, 0.025) | NS | ||||
| eGFR | 0.010 (−0.023, 0.027) | NS | ||||
| ALT (U/L) | −0.064 (−0.022, 0.006) | NS | ||||
| Total cholesterol (mmol/L) | −0.027 (−0.096, 0.091) | NS | ||||
| * Triacylglycerol (mmol/L) | −0.048 (−6.922, 5.091) | NS | ||||
| HDL-C (mmol/L) | −0.070 (−0.117, 0.103) | NS | ||||
| LDL-C (mmol/L) | 0.076 (−0.086, 0.103) | NS | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, A.-C.; Hung, H.-C.; Chen, Y.-W.; Cheng, K.-P.; Li, C.-H.; Lin, C.-H.; Chang, C.-J.; Wu, H.-T.; Ou, H.-Y. Elevated Hedgehog-Interacting Protein Levels in Subjects with Prediabetes and Type 2 Diabetes. J. Clin. Med. 2019, 8, 1635. https://doi.org/10.3390/jcm8101635
Lin A-C, Hung H-C, Chen Y-W, Cheng K-P, Li C-H, Lin C-H, Chang C-J, Wu H-T, Ou H-Y. Elevated Hedgehog-Interacting Protein Levels in Subjects with Prediabetes and Type 2 Diabetes. Journal of Clinical Medicine. 2019; 8(10):1635. https://doi.org/10.3390/jcm8101635
Chicago/Turabian StyleLin, An-Chi, Hao-Chang Hung, Yun-Wen Chen, Kai-Pi Cheng, Chung-Hao Li, Ching-Han Lin, Chih-Jen Chang, Hung-Tsung Wu, and Horng-Yih Ou. 2019. "Elevated Hedgehog-Interacting Protein Levels in Subjects with Prediabetes and Type 2 Diabetes" Journal of Clinical Medicine 8, no. 10: 1635. https://doi.org/10.3390/jcm8101635
APA StyleLin, A. -C., Hung, H. -C., Chen, Y. -W., Cheng, K. -P., Li, C. -H., Lin, C. -H., Chang, C. -J., Wu, H. -T., & Ou, H. -Y. (2019). Elevated Hedgehog-Interacting Protein Levels in Subjects with Prediabetes and Type 2 Diabetes. Journal of Clinical Medicine, 8(10), 1635. https://doi.org/10.3390/jcm8101635