Development and Disease-Dependent Dynamics of Spermatogonial Subpopulations in Human Testicular Tissues

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Testicular Tissue Collection
2.2.1. Immature Patients
2.2.2. Adult Controls
2.3. Histological and Immunohistochemical Evaluation of Human Testicular Tissue
2.4. Spermatogonial Quantification
2.5. Spermatogonial Labeling Index
2.6. Statistical Analyses
3. Results
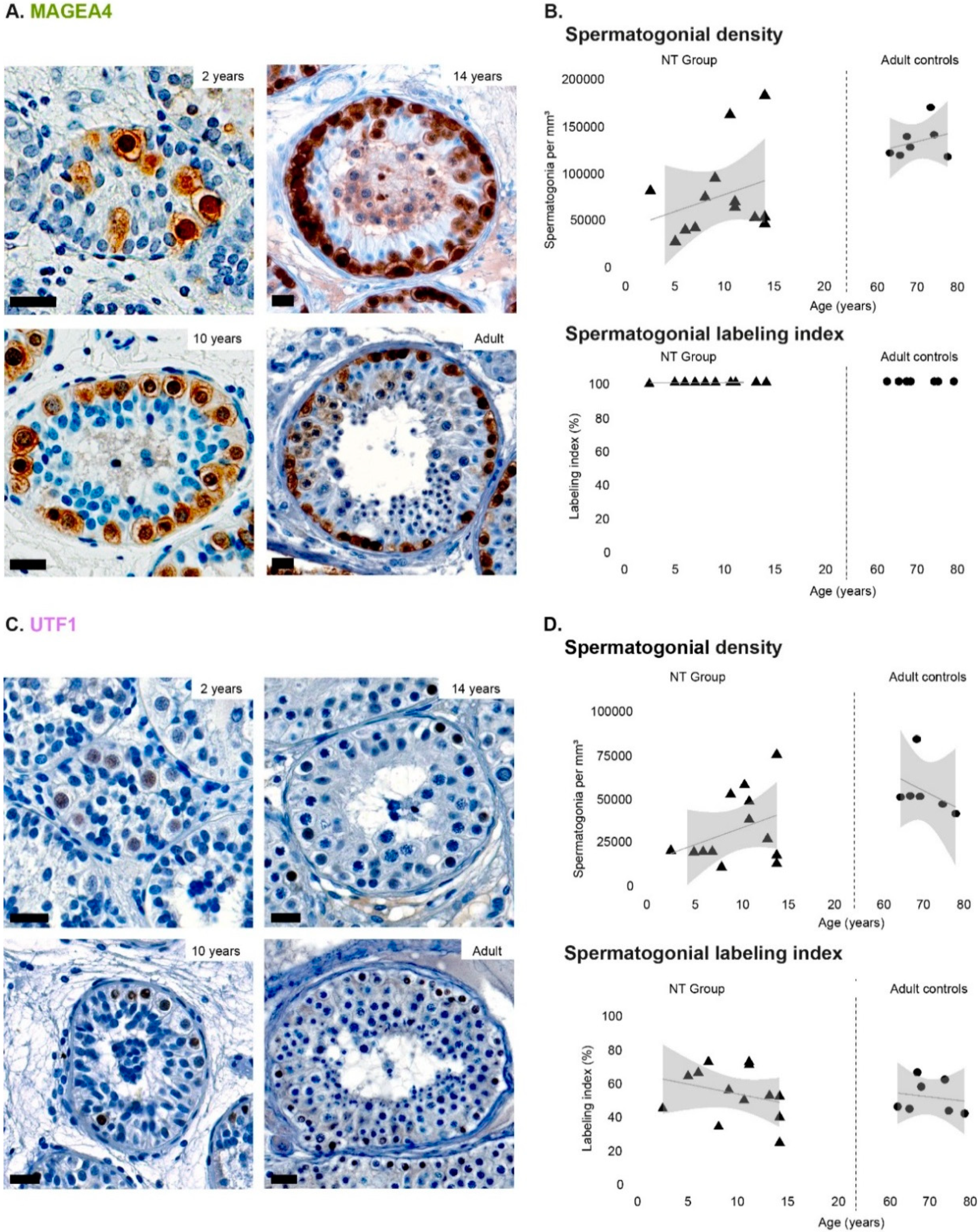
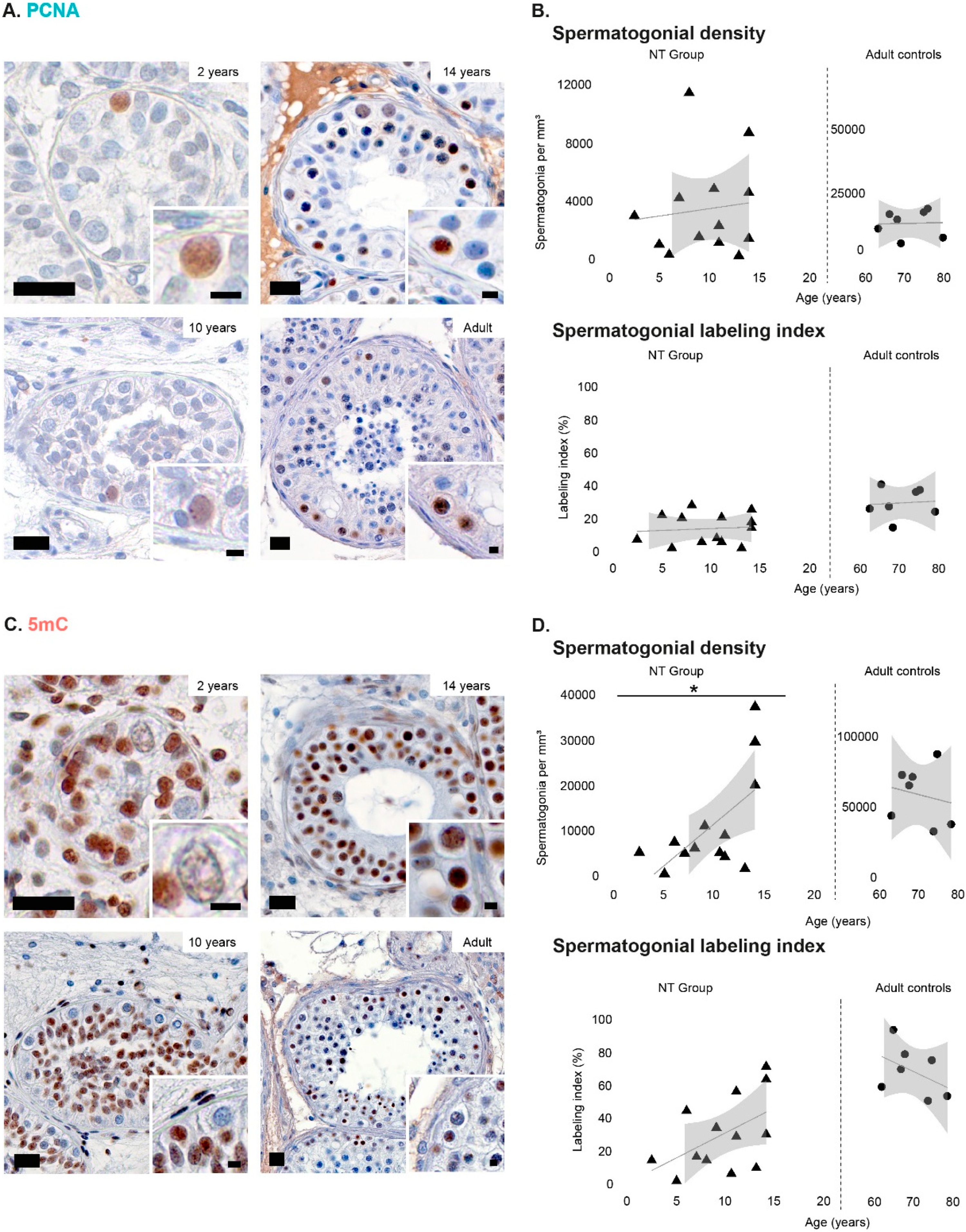
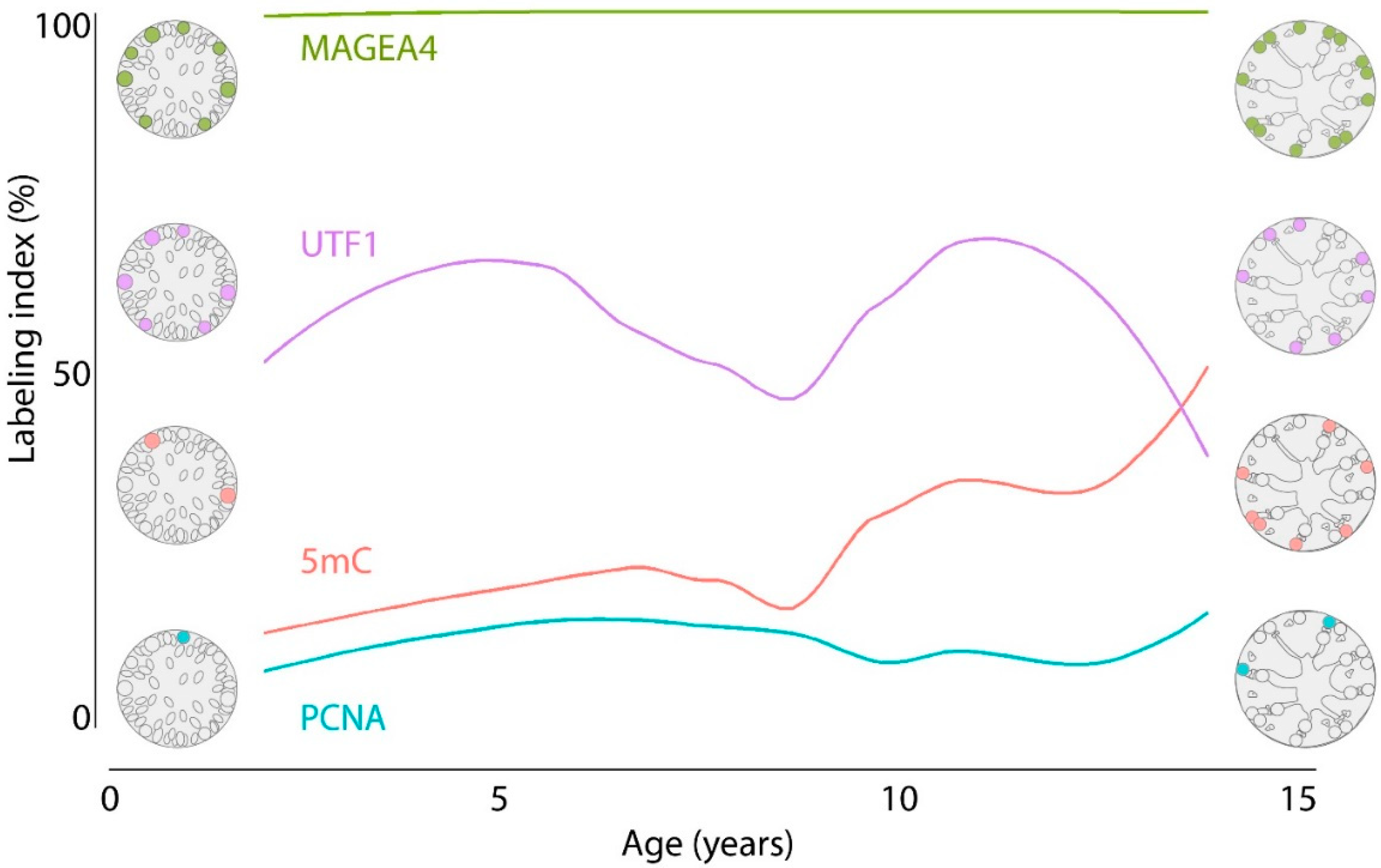
3.1. Density of Spermatogonial Subpopulations during Normal Testicular Development
3.2. Spermatogonial Labeling Indices during Normal Testicular Development
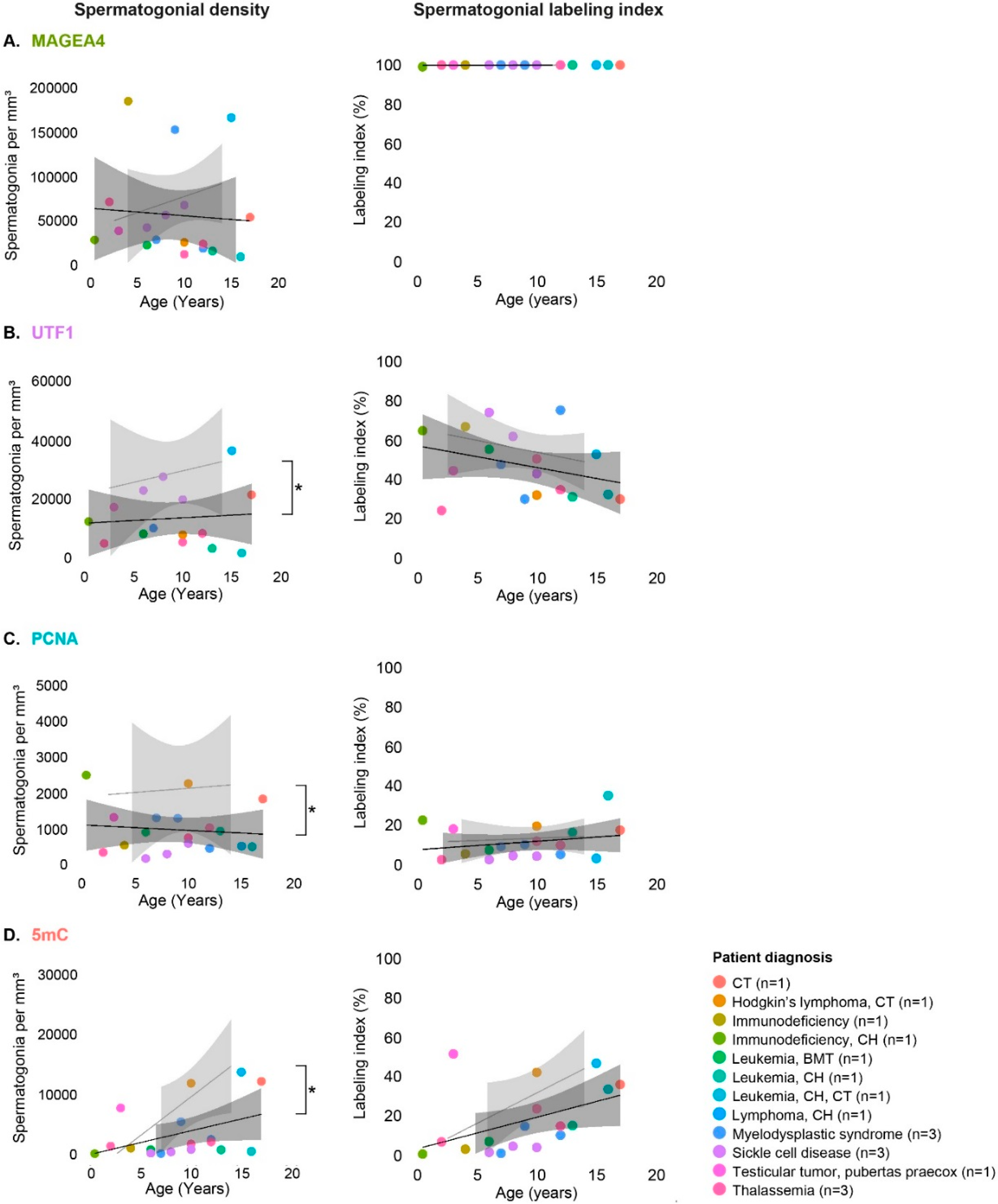
3.3. Spermatogonial Subpopulations in Testicular Tissues of Patients Affected by Disease or Treatment
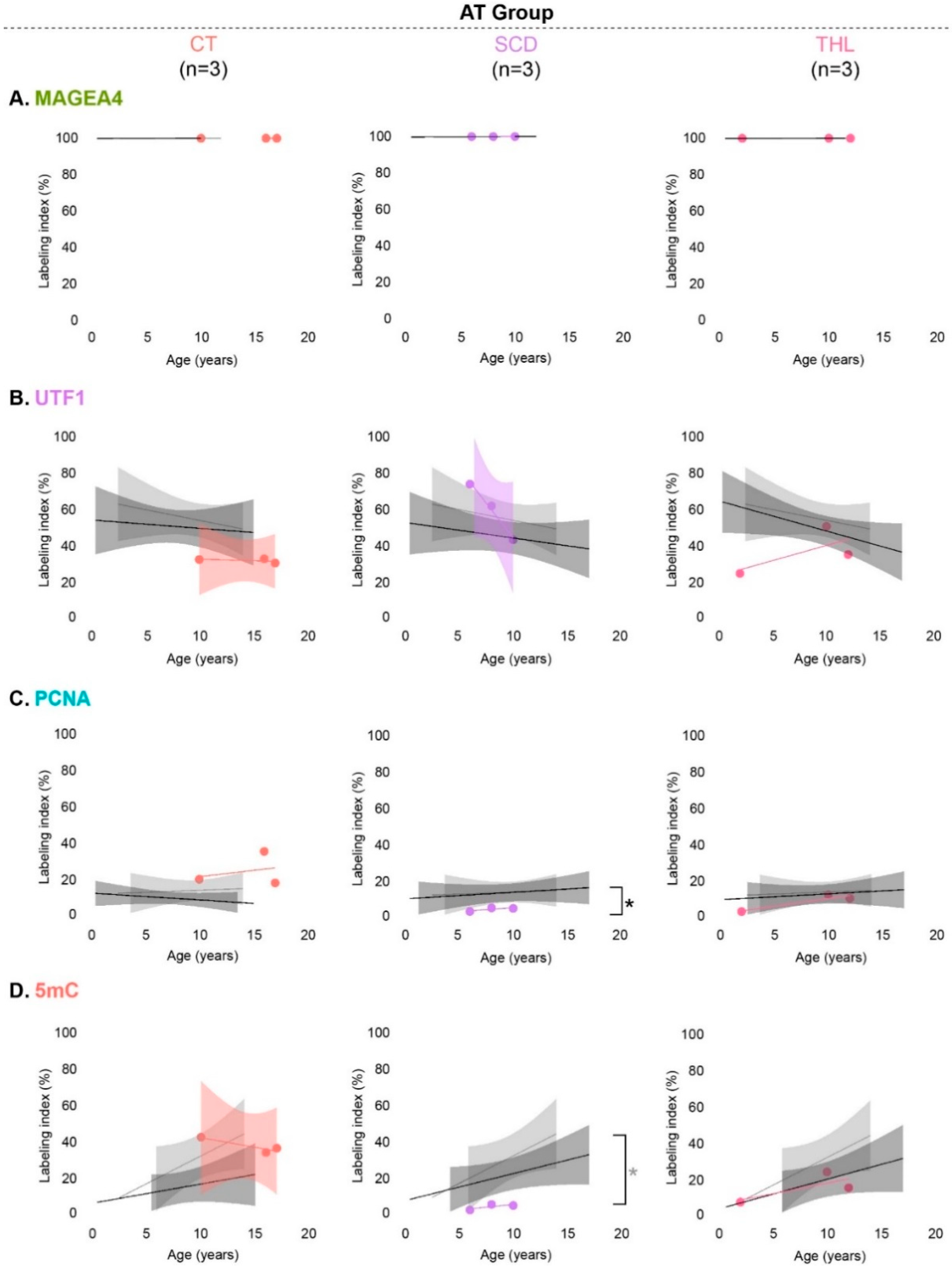
3.4. Spermatogonial Marker Expression in Patients with Cryptorchidism, Sickle Cell Disease and Thalassemia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wikstrom, A.M.; Raivo, T.; Hadziselimovic, F.; Wikstrom, S.; Tuuri, T.; Dunkel, L. Klinefelter syndrome in adolescence: Onset of puberty is associated with accelerated germ cell depletion. J. Clin. Endocrinol. Metab. 2004, 89, 2263–2270. [Google Scholar] [CrossRef] [PubMed]
- Aksglaede, L.; Wikström, A.M.; Rajpert-De Meyts, E.; Dunkel, L.; Skakkebaek, N.E.; Juul, A. Natural history of seminiferous tubule degeneration in Klinefelter syndrome. Hum. Reprod. Update 2006, 12, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Stukenborg, J.B.; Kjartansdóttir, K.R.; Reda, A.; Colon, E.; Albersmeier, J.P.; Söder, O. Male germ cell development in humans. Horm. Res. Paediatr. 2014, 81, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Van Saen, D.; Pino Sánchez, J.; Ferster, A.; Van Der Werff Ten Bosch, J.; Tournaye, H.; Goossens, E. Is the protein expression window during testicular development affected in patients at risk for stem cell loss? Hum. Reprod. 2015, 30, 2859–2870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heckmann, L.; Langenstroth-Röwer, D.; Pock, T.; Wistuba, J.; Stukenborg, J.B.; Zitzmann, M.; Kliesch, S.; Schlatt, S.; Neuhaus, N. A diagnostic germ cell score for immature testicular tissue at risk of germ cell loss. Hum. Reprod. 2018, 33, 636–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crofton, P.M.; Thomson, A.B.; Evans, A.E.; Groome, N.P.; Bath, L.E.; Kelnar, C.J.; Wallace, W.H. Is inhibin B a potential marker of gonadotoxicity in prepubertal children treated for cancer? Clin. Endocrinol. 2003, 58, 296–301. [Google Scholar] [CrossRef]
- Wigny, K.M.; Van Dorp, W.; Van Der Kooi, A.L.; De Rijke, Y.B.; De Vries, A.C.; Smit, M.; Pluijm, S.M.; Van Den Akker, E.L.; Pieters, R.; Laven, J.S.; et al. Gonadal function in boys with newly diagnosed cancer before the start of treatment. Hum. Reprod. 2016, 31, 2613–2618. [Google Scholar] [CrossRef]
- Maymon, B.B.S.; Yogev, L.; Marks, A.; Hauser, R.; Botchan, A.; Yavetz, H. Sertoli cell inactivation by cytotoxic damage to the human testis after cancer chemotherapy. Fertil. Steril. 2014, 81, 1391–1394. [Google Scholar] [CrossRef]
- Poganitsch-Korhonen, M.; Masliukaite, I.; Nurmio, M.; Lähteenmäki, P.; Van Wely, M.; Van Pelt, A.M.M.; Jahnukainen, K.; Stukenborg, J.B. Decreased spermatogonial quantity in prepubertal boys with leukaemia treated with alkylating agents. Leukemia 2017, 31, 1460–1463. [Google Scholar] [CrossRef]
- Stukenborg, J.B.; Alves-Lopes, J.P.; Kurek, M.; Albalushi, H.; Reda, A.; Keros, V.; Töhönen, V.; Bjarnason, R.; Romerius, P.; Sundin, M.; et al. Spermatogonial quantity in human prepubertal testicular tissue collected for fertility preservation prior to potentially sterilizing therapy. Hum. Reprod. 2018, 33, 1677–1683. [Google Scholar] [CrossRef]
- Jahnukainen, K.; Ehmcke, J.; Hou, M.; Schlatt, S. Testicular function and fertility preservation in male cancer patients. Best Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 287–302. [Google Scholar] [CrossRef] [PubMed]
- Jahnukainen, K.; Mitchell, R.T.; Stukenborg, J.B. Testicular function and fertility preservation after treatment for haematological cancer. Curr. Opin. Endocrinol. Diabetes Obes. 2015, 22, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Green, D.M.; Nolan, V.G.; Goodman, P.J.; Whitton, J.A.; Srivastava, D.; Leisenring, W.M.; Neglia, J.P.; Sklar, C.A.; Kaste, S.C.; Hudson, M.M.; et al. The cyclophosphamide equivalent dose as an approach for quantifying alkylating agent exposure: A report from the Childhood Cancer Survivor Study. Pediatr. Blood Cancer 2014, 61, 53–67. [Google Scholar] [CrossRef] [Green Version]
- Picton, H.M.; Wyns, C.; Anderson, R.A.; Goossens, E.; Jahnukainen, K.; Kliesch, S.; Mitchell, R.T.; Pennings, G.; Rives, N.; Tournaye, H.; et al. A European perspective on testicular tissue cryopreservation for fertility preservation in prepubertal and adolescent boys. Hum. Reprod. 2015, 30, 2463–2475. [Google Scholar] [CrossRef] [PubMed]
- Valli-Pulaski, H.; Peters, K.A.; Gassei, K.; Steimer, S.R.; Sukhwani, M.; Hermann, B.P.; Dwomor, L.; David, S.; Fayomi, A.P.; Munyoki, S.K.; et al. Testicular tissue cryopreservation: 8 years of experience from a coordinated network of academic centers. Hum. Reprod. 2019, 34, 966–977. [Google Scholar] [CrossRef]
- Gaskell, T.L.; Esnal, A.; Robinson, L.L.; Anderson, R.A.; Saunders, P.T. Immunohistochemical profiling of germ cells within the human fetal testis: Identification of three subpopulations. Biol. Reprod. 2004, 71, 2012–2021. [Google Scholar] [CrossRef] [PubMed]
- Honecker, F.; Stoop, H.; De Krijger, R.R.; Chris Lau, Y.F.; Bokemeyer, C.; Looijenga, L.H. Pathobiological implications of the expression of markers of testicular carcinoma in situ by fetal germ cells. J. Pathol. 2004, 203, 849–857. [Google Scholar] [CrossRef]
- Mitchell, R.T.; Cowan, G.; Morris, K.D.; Anderson, R.A.; Fraser, H.M.; Mckenzie, K.J.; Wallace, W.H.; Kelnar, C.J.; Saunders, P.T.; Sharpe, R.M. Germ cell differentiation in the marmoset (Callithrix jacchus) during fetal and neonatal life closely parallels that in the human. Hum. Reprod. 2008, 23, 2755–2765. [Google Scholar] [CrossRef] [Green Version]
- Steger, K.; Aleithe, I.; Behre, H.; Bergmann, M. The proliferation of spermatogonia in normal and pathological human seminiferous epithelium: An immunohistochemical study using monoclonal antibodies against Ki-67 protein and proliferating cell nuclear antigen. Mol. Hum. Reprod. 1998, 4, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Berensztein, E.B.; Sciara, M.I.; Rivarola, M.A.; Belgorosky, A. Apoptosis and proliferation of human testicular somatic and germ cells during prepuberty: High rate of testicular growth in newborns mediated by decreased apoptosis. J. Clin. Endocrinol. Metab. 2002, 87, 5113–5118. [Google Scholar] [CrossRef] [Green Version]
- Masliukaite, I.; Hagen, J.M.; Jahnukainen, K.; Stukenborg, J.B.; Repping, S.; Van Der Veen, F.; Van Wely, M.; Van Pelt, A.M. Establishing reference values for age-related spermatogonial quantity in prepubertal human testes: A systematic review and meta-analysis. Fertil. Steril. 2016, 106, 1652–1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Kopylow, K.; Staege, H.; Spiess, A.N.; Schulze, W.; Will, H.; Primig, M.; Kirchhoff, C. Differential marker protein expression specifies rarefaction zone-containing human Adark spermatogonia. Reproduction 2012, 143, 45–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Persio, S.; Saracino, R.; Fera, S.; Muciaccia, B.; Esposito, V.; Boitani, C.; Berloco, B.P.; Nudo, F.; Spadetta, G.; Stefanini, M.; et al. Spermatogonial kinetics in humans. Development 2017, 144, 3430–3439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Grow, E.J.; Mlcochova, H.; Maher, G.J.; Lindskog, C.; Nie, X.; Guo, Y.; Takei, Y.; Yun, J.; Cai, L.; et al. The adult human testis; transcriptional cell atlas. Cell Res. 2018, 28, 1141–1157. [Google Scholar] [CrossRef]
- Hermann, B.P.; Cheng, K.; Singh, A.; Roa-De La Cruz, L.; Mutoji, K.N.; Chen, I.C.; Gildersleeve, H.; Lehle, J.D.; Mayo, M.; Westernströer, B.; et al. The mammalian spermatogenesis single-cell transcriptome, from spermatogonial stem cells to spermatids. Cell Rep. 2018, 25, 1650–1667. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Liu, X.; Chang, G.; Chen, Y.; An, G.; Yan, L.; Gao, S.; Xu, Y.; Cui, Y.; Dong, J.; et al. Single-cell RNA sequencing analysis reveals sequential cell fate transition during human spermatogenesis. Cell Stem Cell 2018, 23, 599–614. [Google Scholar] [CrossRef] [Green Version]
- Sohni, A.; Tan, K.; Song, H.W.; Burow, D.; De Rooij, D.G.; Laurent, L.; Hsieh, T.C.; Rabah, R.; Hammoud, S.S.; Vicini, E.; et al. The neonatal and adult human testis defined at the single-cell level. Cell Rep. 2019, 26, 1501–1517. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Shichijo, S.; Noguchi, M.; Hirohata, M.; Itoh, K. Identification of MAGE-1 and MAGE-4 proteins in spermatogonia and primary spermatocytes of testis. Cancer Res. 1995, 55, 3478–3482. [Google Scholar]
- Aubry, F.; Satie, A.P.; Rioux-Leclercq, N.; Rajpert-De Meyts, E.; Spagnoli, G.C.; Chomez, P.; De Backer, O.; Jégou, B.; Samson, M. MAGE-A4, a germ cell specific marker, is expressed differentially in testicular tumors. Cancer 2001, 92, 2778–2785. [Google Scholar] [CrossRef]
- Pohl, E.; Höffken, V.; Schlatt, S.; Kliesch, S.; Gromoll, J.; Wistuba, J. Ageing in men with normal spermatogenesis alters spermatogonial dynamics and nuclearmorphology in Sertoli cells. Andrology 2019, in press. [Google Scholar] [CrossRef]
- Kristensen, D.M.; Nielsen, J.E.; Skakkebaek, N.E.; Graem, N.; Jacobsen, G.K.; Rajpert-De Meyts, E.; Leffers, H. Presumed pluripotency markers UTF-1 and REX-1 are expressed in human adult testes and germ cell neoplasms. Hum. Reprod. 2008, 23, 775–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Kopylow, K.; Kirchhoff, C.; Jezek, D.; Schulze, W.; Feig, C.; Primig, M.; Steinkraus, V.; Spiess, A.N. Screening for biomarkers of spermatogonia within the human testis: A whole genome approach. Hum. Reprod. 2010, 25, 1104–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Grow, E.J.; Yi, C.; Mlcochova, H.; Maher, G.J.; Lindskog, C.; Murphy, P.J.; Wike, C.L.; Carrell, D.T.; Goriely, A.; et al. Chromatin and single-cell RNA-seq profiling reveal dynamic signaling and metabolic transitions during human spermatogonial stem cell development. Cell Stem Cell 2017, 21, 533–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adiga, S.K.; Ehmcke, J.; Schlatt, S.; Kliesch, S.; Westernströer, B.; Luetjens, C.M.; Wistuba, J.; Gromoll, J. Reduced expression of DNMT3B in the germ cells of patients with bilateral spermatogenic arrest does not lead to changes in the global methylation status. Mol. Hum. Reprod. 2011, 17, 545–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nettersheim, D.; Heukamp, L.C.; Fronhoffs, F.; Grewe, M.J.; Haas, N.; Waha, A.; Honecker, F.; Waha, A.; Kristiansen, G.; Schorle, H. Analysis of TET expression/activity and 5mC oxidation during normal and malignant germ cell development. PLoS ONE 2013, 8, e82881. [Google Scholar] [CrossRef] [PubMed]
- Fend-Guella, D.L.; Von Kopylow, K.; Spiess, A.N.; Schulze, W.; Salzbrunn, A.; Diederich, S.; El Hajj, N.; Haaf, T.; Zechner, U.; Linke, M. The DNA methylation profile of human spermatogonia at single-cell-and single-allele-resolution refutes its role in spermatogonial stem cell function and germ cell differentiation. Mol. Hum. Reprod. 2019, 25, 283–294. [Google Scholar] [CrossRef]
- Kaneda, M.; Okano, M.; Hata, K.; Sado, T.; Tsujimoto, N.; Li, E.; Sasaki, H. Essential role for de novo DNA methyltransferase Dnmt3a in paternal and maternal imprinting. Nature 2004, 429, 900–903. [Google Scholar] [CrossRef]
- Urdinguio, R.G.; Bayón, G.F.; Dmitrijeva, M.; Toraño, E.G.; Bravo, C.; Fraga, M.F.; Bassas, L.; Larriba, S.; Fernández, A.F. Aberrant DNA methylation patterns of spermatozoa in men with unexplained infertility. Hum. Reprod. 2015, 30, 1014–1028. [Google Scholar] [CrossRef] [Green Version]
- Langenstroth-Röwer, D.; Gromoll, J.; Wistuba, J.; Tröndle, I.; Laurentino, S.; Schlatt, S.; Neuhaus, N. De novo methylation in male germ cells of the common marmoset monkey occurs during postnatal development and is maintained in vitro. Epigenetics 2017, 12, 527–539. [Google Scholar] [CrossRef]
- Behre, H.M.; Nashan, D.; Nieschlag, E. Objective measurement of testicular volume by ultrasonography: Evaluation of the technique and comparison with orchidometer estimates. Int. J. Androl. 1989, 12, 395–403. [Google Scholar] [CrossRef]
- Uijldert, M.; Meißner, A.; De Melker, A.A.; Van Pelt, A.M.M.; Van De Wetering, M.D.; Van Rijn, R.R.; Van Wely, M.; Van Der Veen, F.; Repping, S. Development of the testis in pre-pubertal boys with cancer after biopsy for fertility preservation. Hum. Reprod. 2017, 32, 2366–2372. [Google Scholar] [CrossRef] [PubMed]
- Pohl, E.; Gromoll, J.; Kliesch, S.; Wistuba, J. An alternative interpretation of cellular ‘selfish spermatogonial selection’-clusters in the human testis indicates the need for 3-D-analyses. Andrology 2016, 4, 213–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, M.; Kliesch, S. Testicular biopsy and histology. In Andrology Male Reproductive Health and Dysfunction; Nieschlag, E., Behre, H.M., Nieschlag, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 155–167. [Google Scholar]
- Albert, S.; Wistuba, J.; Eildermann, K.; Ehmcke, J.; Schlatt, S.; Gromoll, J.; Kossack, N. Comparative marker analysis after isolation and culture of testicular cells from the immature marmoset. Cells Tissues Organs 2012, 196, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Paniagua, R.; Nistal, M. Morphological and histometric study of human spermatogonia from birth to the onset of puberty. J. Anat. 1984, 139, 535–552. [Google Scholar]
- Gundersen, H.J.G.; Jensen, E.B. Stereological estimation of the volume weighted mean volume of arbitrary particles observed on random sections. J. Microsc. 1985, 138, 127–142. [Google Scholar] [CrossRef]
- Laurentino, S.; Borgmann, J.; Gromoll, J. On the origin of sperm epigenetic heterogeneity. Reproduction 2016, 151, R71–R78. [Google Scholar] [CrossRef] [Green Version]
- Kafri, T.; Ariel, M.; Brandeis, M.; Shemer, R.; Urven, L.; McCarrey, J.; Cedar, H.; Razin, A. Developmental pattern of gene-specific DNA methylation in the mouse embryo and germ line. Genes Dev. 1992, 6, 705–714. [Google Scholar] [CrossRef] [Green Version]
- Kubo, N.; Toh, H.; Shirane, K.; Shirakawa, T.; Kobayashi, H.; Sato, T.; Sone, H.; Sato, Y.; Tomizawa, S.; Tsurusaki, Y.; et al. DNA methylation and gene expression dynamics during spermatogonial stem cell differentiation in the early postnatal mouse testis. BMC Genom. 2015, 16, 624. [Google Scholar] [CrossRef] [Green Version]
- Marques, C.J.; Joao Pinho, M.; Carvalho, F.; Bieche, I.; Barros, A.; Sousa, M. DNA methylation imprinting marks and DNA methyltransferase expression in human spermatogenic cell stages. Epigenetics 2011, 6, 1354–1361. [Google Scholar] [CrossRef] [Green Version]
- Nagano, M.C. Homing efficiency and proliferation kinetics of male germ line stem cells following transplantation in mice. Biol. Reprod. 2003, 69, 701–707. [Google Scholar] [CrossRef]
- Hermann, B.P.; Sukhwani, M.; Winkler, F.; Pascarella, J.N.; Peters, K.A.; Sheng, Y.; Valli, H.; Rodriguez, M.; Ezzelarab, M.; Daego, G.; et al. Spermatogonial stem cell transplantation into rhesus testes regenerates spermatogenesis producing functional sperm. Cell Stem Cell 2012, 11, 715–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reda, A.; Hou, M.; Winton, T.R.; Chapin, R.E.; Söder, O.; Stukenborg, J.B. In vitro differentiation of rat spermatogonia into round spermatids in tissue culture. Mol. Hum. Reprod. 2016, 22, 601–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumont, L.; Arkoun, B.; Jumeau, F.; Milazzo, J.P.; Bironneau, A.; Liot, D.; Wils, J.; Rondanino, C.; Rives, N. Assessment of the optimal vitrification protocol for pre-pubertal mice testes leading to successful in vitro production of flagellated spermatozoa. Andrology 2015, 3, 611–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Michele, F.; Poels, J.; Vermeulen, M.; Ambroise, J.; Gruson, D.; Guiot, Y.; Wyns, C. Haploid germ cells generated in organotypic culture of testicular tissue from prepubertal boys. Front. Physiol. 2018, 9, 1413. [Google Scholar] [CrossRef]
- Jahnukainen, K.; Ehmcke, J.; Schlatt, S. Testicular xenografts: A novel approach to study cytotoxic damage in juvenile primate testis. Cancer Res. 2006, 66, 3813–3818. [Google Scholar] [CrossRef] [Green Version]
- Jahnukainen, K.; Ehmcke, J.; Nurmio, M.; Schlatt, S. Autologous ectopic grafting of cryopreserved testicular tissue preserves the fertility of prepubescent monkeys that receive sterilizing cytotoxic therapy. Cancer Res. 2012, 72, 5174–5178. [Google Scholar] [CrossRef] [Green Version]
- Cortes, D.; Thorup, J.M.; Beck, B.L. Quantitative histology of germ cells in the undescended testes of human fetuses, neonates and infants. J. Urol. 1995, 154, 1188–1192. [Google Scholar] [CrossRef]
- Nistal, M.; Riestra, M.L.; Paniagua, R. Correlation between testicular biopsies (prepubertal and postpubertal) and spermiogram in cryptorchid men. Hum. Pathol. 2000, 31, 1022–1030. [Google Scholar] [CrossRef]
- Li, R.; Thorup, J.; Sun, C.; Cortes, D.; Southwell, B.; Hutson, J. Immunofluorescent analysis of testicular biopsies with germ cell and Sertoli cell markers shows significant MVH negative germ cell depletion with older age at orchiopexy. J. Urol. 2014, 191, 458–464. [Google Scholar] [CrossRef]
- Hadziselimovic, F.; Herzog, B. The importance of both an early orchidopexy and germ cell maturation for fertility. Lancet 2001, 358, 1156–1157. [Google Scholar] [CrossRef]
- Seguchi, H.; Hadziselimović, F. Ultramicroscopic studies on the seminiferous tubule in children from birth to puberty. I. Spermatogonia development. Verh. Anat. Ges. 1974, 68, 133–148. [Google Scholar]
- Hadziselimovic, F.; Hocht, B.; Herzog, B.; Buser, M.W. Infertility in cryptorchidism is linked to the stage of germ cell development at orchidopexy. Horm. Res. 2007, 68, 46–52. [Google Scholar] [CrossRef]
- Walker, A.L.; Steward, S.; Howard, T.A.; Mortier, N.; Smeltzer, M.; Wang, Y.; Ware, R.E. Epigenetic and molecular profiles of erythroid cells after hydroxyurea treatment in sickle cell anemia. Blood 2011, 118, 5664–5670. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Patient Number | Age (Years) | Testis Volume (mL) | Diagnosis (Treatment before Testis Biopsy) |
|---|---|---|---|---|
| NT | NT1 | 2.5 | 1.5 | Ependymoma |
| NT2 * | 3 | 0.33 | Acute myeloid leukemia | |
| NT3 | 5 | 1 | Hodgkin’s lymphoma | |
| NT4 | 6 | 1 | Hodgkin’s lymphoma | |
| NT5 | 7 | 0.83 | Rhabdomyosarcoma | |
| NT6 | 8 | 0.40 | Ewing sarcoma | |
| NT7 | 9 | 1.30 | Intracranial germ cell tumor | |
| NT8 | 10.5 | 1.80 | Rhabdomyosarcoma | |
| NT9 | 11 | 1 | Hodgkin’s lymphoma | |
| NT10 | 11 | 2 | Osteosarcoma | |
| NT11 | 13 | 1 | Hodgkin’s lymphoma | |
| NT12 | 14 | 8 # | Sarcoma | |
| NT13 | 14 | 19 | Lymphoma | |
| NT14 | 14 | 9 | Non-Hodgkin’s lymphoma | |
| AT | AT1 | 5 months | - | Immunodeficiency (chemotherapy) |
| AT2 | 2 | 0.3 | Thalassemia | |
| AT3 | 3 | 8 | Testicular tumor, pubertas praecox | |
| AT4 | 4 | 0.4 | Immunodeficiency | |
| AT5 | 6 | - | Leukemia (allogeneic bone marrow transplantation) | |
| AT6 | 6 | 1 | Sickle cell disease (hydroxyurea) | |
| AT7 | 7 | 1 | Myelodysplastic syndrome | |
| AT8 * | 7 | 1 | Sickle cell disease, cryptorchid testes | |
| AT9 | 8 | 0.3 | Sickle cell disease (hydroxyurea) | |
| AT10 | 9 | 1 | Myelodysplastic syndrome | |
| AT11 | 10 | 1 | Thalassemia major | |
| AT12 | 10 | 1 | Hodgkin’s lymphoma, cryptorchid testes | |
| AT13 | 10 | 2 | Sickle cell disease (hydroxyurea) | |
| AT14 | 12 | 1.4 | Myelodysplastic syndrome | |
| AT15 * | 12 | - | Cryptorchid testes, hypogonadism | |
| AT16 | 12 | 1 | Thalassemia major | |
| AT17 | 13 | 8 | Leukemia (chemotherapy) | |
| AT18 * | 14 | 6 | Cryptorchid testes | |
| AT19 | 15 | 6 | Lymphoma (chemotherapy) | |
| AT20 | 16 | 5 | Leukemia, cryptorchid testes (chemotherapies) | |
| AT21 * | 16 | 3 | Testicular tumor, cryptorchid testes | |
| AT22 | 17 | 3 | Cryptorchid testes |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Portela, J.M.D.; Heckmann, L.; Wistuba, J.; Sansone, A.; van Pelt, A.M.M.; Kliesch, S.; Schlatt, S.; Neuhaus, N. Development and Disease-Dependent Dynamics of Spermatogonial Subpopulations in Human Testicular Tissues. J. Clin. Med. 2020, 9, 224. https://doi.org/10.3390/jcm9010224
Portela JMD, Heckmann L, Wistuba J, Sansone A, van Pelt AMM, Kliesch S, Schlatt S, Neuhaus N. Development and Disease-Dependent Dynamics of Spermatogonial Subpopulations in Human Testicular Tissues. Journal of Clinical Medicine. 2020; 9(1):224. https://doi.org/10.3390/jcm9010224
Chicago/Turabian StylePortela, Joana M. D., Laura Heckmann, Joachim Wistuba, Andrea Sansone, Ans M. M. van Pelt, Sabine Kliesch, Stefan Schlatt, and Nina Neuhaus. 2020. "Development and Disease-Dependent Dynamics of Spermatogonial Subpopulations in Human Testicular Tissues" Journal of Clinical Medicine 9, no. 1: 224. https://doi.org/10.3390/jcm9010224
APA StylePortela, J. M. D., Heckmann, L., Wistuba, J., Sansone, A., van Pelt, A. M. M., Kliesch, S., Schlatt, S., & Neuhaus, N. (2020). Development and Disease-Dependent Dynamics of Spermatogonial Subpopulations in Human Testicular Tissues. Journal of Clinical Medicine, 9(1), 224. https://doi.org/10.3390/jcm9010224