Comparative Analysis of Thrombin Calibration Algorithms and Correction for Thrombin-α2macroglobulin Activity

 , , and
, , and
Abstract
:1. Introduction
2. Experimental Section
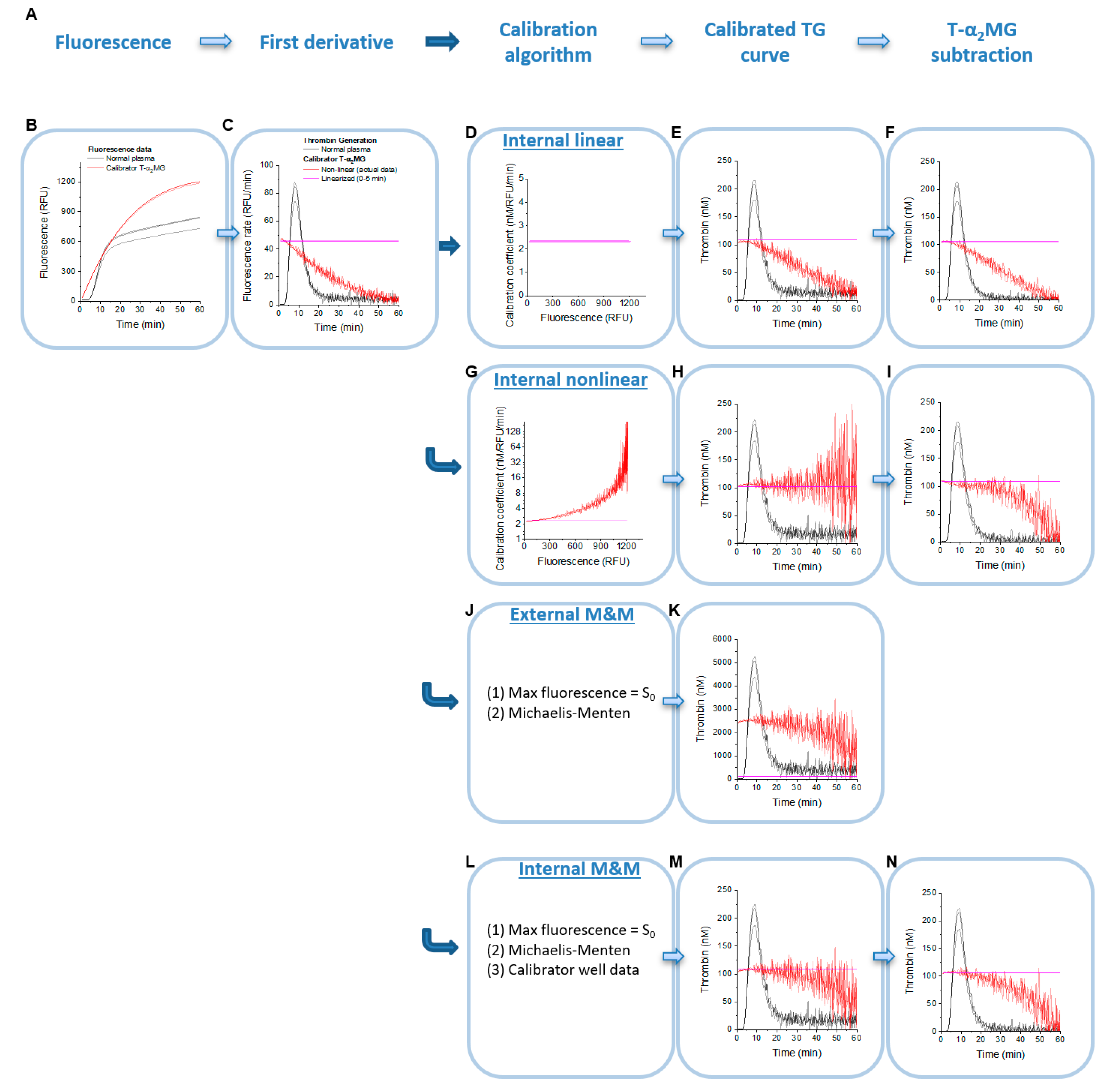
2.1. Principles of TG Data Analysis
2.2. Thrombin Generation Experiments
2.3. Analysis Algorithms and Software
2.4. Internal Linear Thrombin Calibration
2.5. Internal Non-Linear Thrombin Calibration
2.6. External Linear and Non-Linear Thrombin Calibration
2.7. Determination of Km and kcat for Hydrolysis of Z-GGR-AMC by Active Site-Titrated Thrombin
2.8. External and Internal Michaelis–Menten Calibration
3. Results
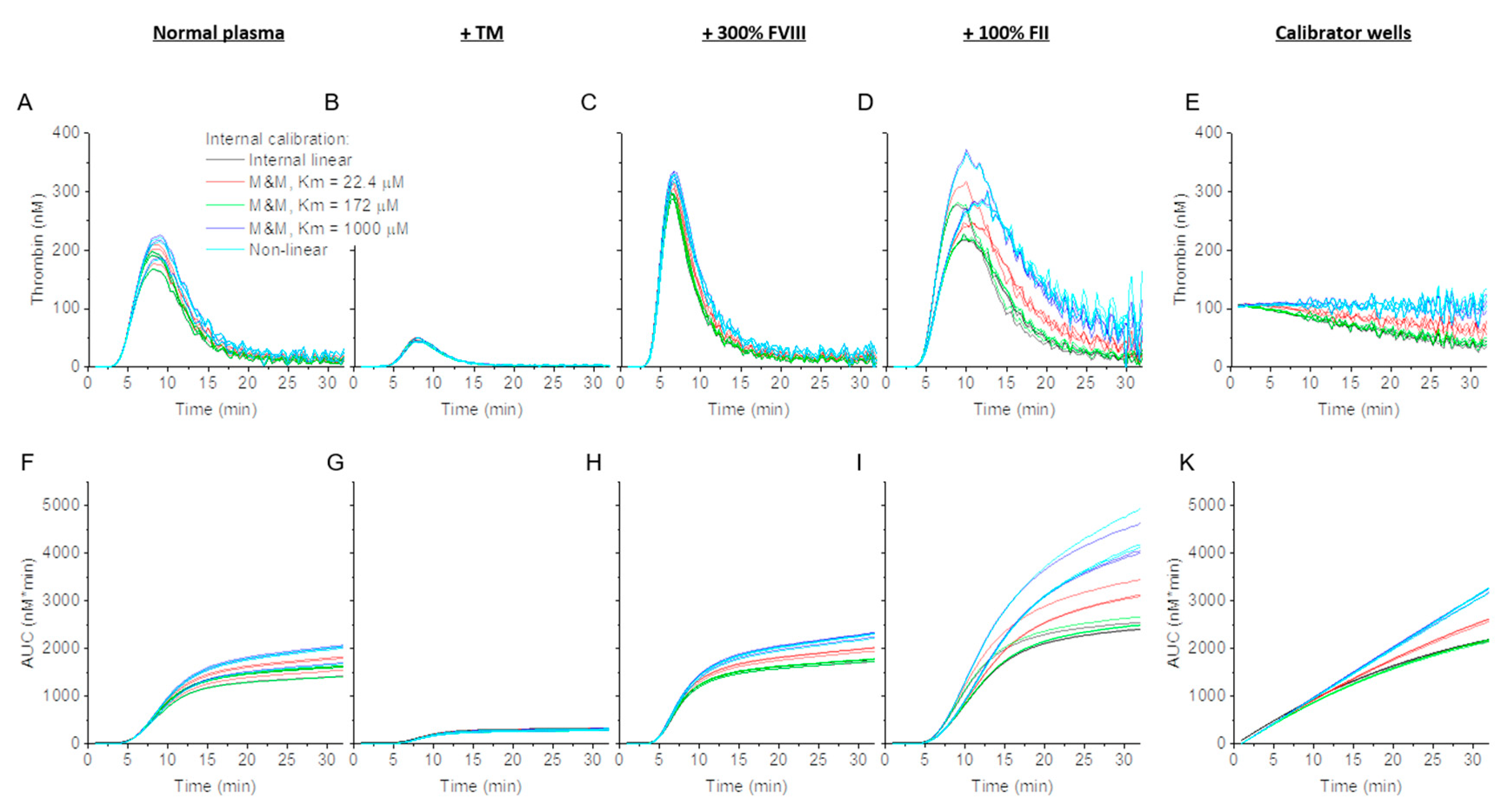
3.1. Effect of Internal Linear and Non-Linear Thrombin Calibration and Michaelis–Menten Formula on Calibrator and Normal TG
3.2. Effect of Thrombin Calibration and Correction on TG in Factor Supplemented Plasma
3.3. Effect of Internal Calibration on TG Parameter Values in PFP
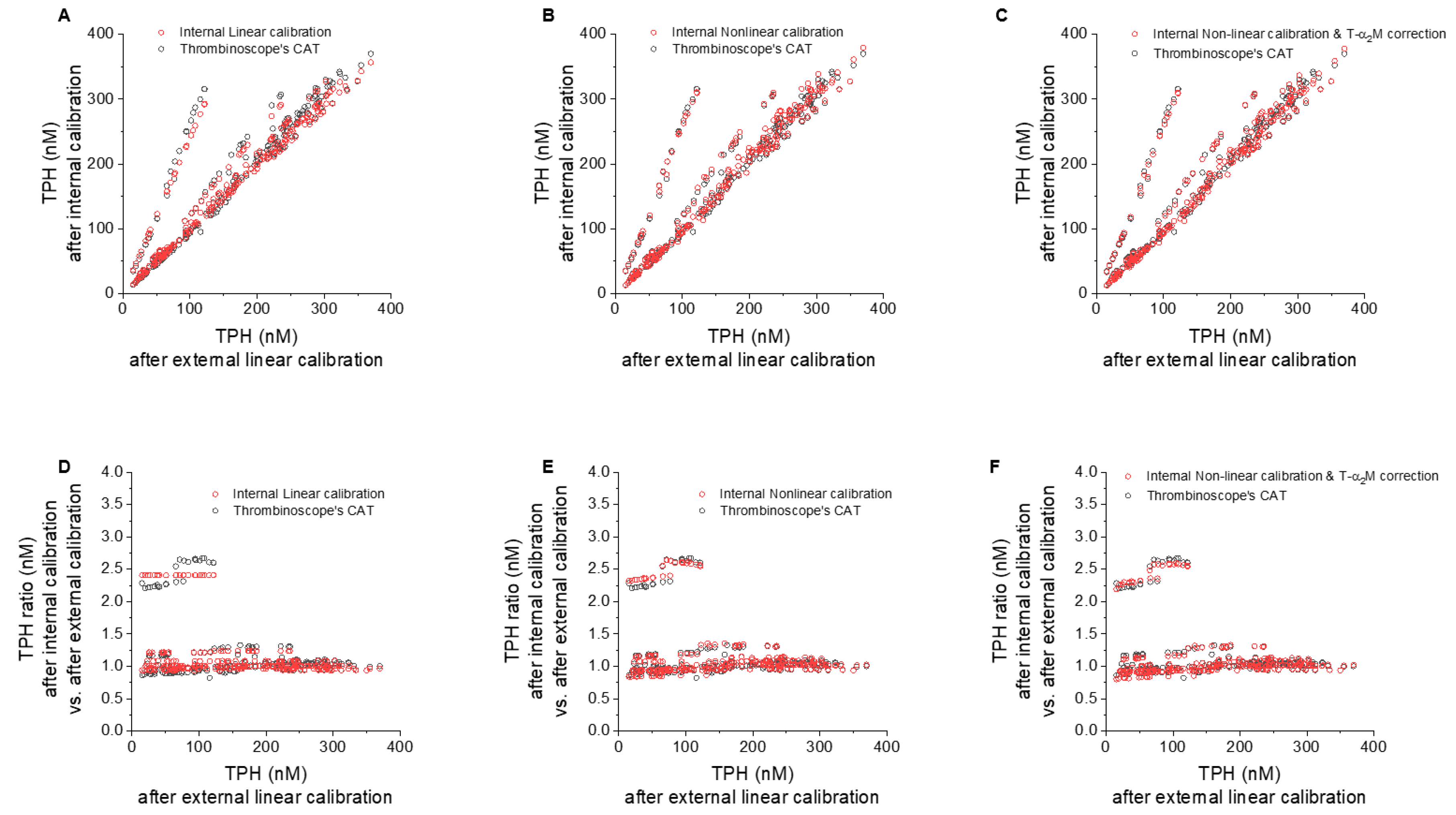
3.4. Individual Contributions of Internal Linear/Non-Linear, and T-α2MG Corrections vs. CAT
3.4.1. Effect of Internal vs. External Linear Thrombin Calibration
3.4.2. Effect of Nonlinear Correction
3.4.3. Effect of T-α2MG Correction
3.5. Effect of Corrections on Run-to-Run and within-Run Variability
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Duarte, R.C.F.; Ferreira, C.N.; Rios, D.R.A.; Reis, H.J.D.; Carvalho, M.D.G. Thrombin generation assays for global evaluation of the hemostatic system: Perspectives and limitations. Rev. Bras. Hematol. Hemoter. 2017, 39, 259–265. [Google Scholar] [CrossRef]
- Tripodi, A. Thrombin generation assay and its application in the clinical laboratory. Clin. Chem. 2016, 62, 699–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdary, P. Anti-tissue factor pathway inhibitor (TFPI) therapy: A novel approach to the treatment of haemophilia. Int. J. Hematol. 2020, 111, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Garland, S.G.; DeRemer, C.E.; Smith, S.M.; Gums, J.G. Betrixaban: A new oral factor Xa inhibitor for extended venous thromboembolism prophylaxis in high-risk hospitalized patients. Ann. Pharmacother. 2018, 52, 554–561. [Google Scholar] [CrossRef] [PubMed]
- Livnat, T.; Sehgal, A.; Qian, K.; Van Nguyen, H.; Madigan, K.; Sorensen, B.; Kenet, G. Thrombin generation in plasma of patients with haemophilia A and B with inhibitors: Effects of bypassing agents and antithrombin reduction. Blood Cells Mol. Dis. 2020, 82, 102416. [Google Scholar] [CrossRef] [PubMed]
- Machin, N.; Ragni, M.V. An investigational RNAi therapeutic targeting antithrombin for the treatment of hemophilia A and B. J. Blood Med. 2018, 9, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Siegal, D.M.; Curnutte, J.T.; Connolly, S.J.; Lu, G.; Conley, P.B.; Wiens, B.L.; Mathur, V.S.; Castillo, J.; Bronson, M.D.; Leeds, J.M.; et al. Andexanet alfa for the reversal of factor Xa inhibitor activity. N. Engl. J. Med. 2015, 373, 2413–2424. [Google Scholar] [CrossRef]
- Dargaud, Y.; Wolberg, A.S.; Gray, E.; Negrier, C.; Hemker, H.C. Proposal for standardized preanalytical and analytical conditions for measuring thrombin generation in hemophilia: Communication from the SSC of the ISTH. J. Thromb. Haemost. 2017, 15, 1704–1707. [Google Scholar] [CrossRef]
- Hemker, H.C. Thrombin generation: Biochemical possibilities and clinical reality. Blood 2015, 126, 288–289. [Google Scholar] [CrossRef] [Green Version]
- Hemker, H.C. The application of thrombin generation in real life clinical situations. Thromb. Res. 2015, 136, 3–4. [Google Scholar] [CrossRef]
- Kintigh, J.; Monagle, P.; Ignjatovic, V. A review of commercially available thrombin generation assays. Res. Pract. Thromb. Haemost. 2018, 2, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Panova-Noeva, M.; van der Meijden, P.E.J.; Ten Cate, H. Clinical applications, pitfalls, and uncertainties of thrombin generation in the presence of platelets. J. Clin. Med. 2019, 9, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panteleev, M.A.; Hemker, H.C. Global/integral assays in hemostasis diagnostics: Promises, successes, problems and prospects. Thromb. J. 2015, 13, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramjee, M.K. The use of fluorogenic substrates to monitor thrombin generation for the analysis of plasma and whole blood coagulation. Anal. Biochem. 2000, 277, 11–18. [Google Scholar] [CrossRef]
- Hemker, H.C.; Giesen, P.; AlDieri, R.; Regnault, V.; de Smed, E.; Wagenvoord, R.; Lecompte, T.; Béguin, S. The calibrated automated thrombogram (CAT): A universal routine test for hyper- and hypocoagulability. Pathophysiol. Haemost Thromb. 2002, 32, 249–253. [Google Scholar] [CrossRef]
- Hemker, H.C.; Giesen, P.; Al, D.R.; Regnault, V.; de Smedt, E.; Wagenvoord, R.; Lecompte, T.; Béguin, S. Calibrated automated thrombin generation measurement in clotting plasma. Pathophysiol. Haemost Thromb. 2003, 33, 4–15. [Google Scholar] [CrossRef]
- Ataullakhanov, F.I.; Guria, G.T.; Sarbash, V.I.; Volkova, R.I. Spatiotemporal dynamics of clotting and pattern formation in human blood. Biochim. Biophys. Acta 1998, 1425, 453–468. [Google Scholar] [CrossRef]
- Krasotkina, Y.V.; Sinauridze, E.I.; Ataullakhanov, F.I. Spatiotemporal dynamics of fibrin formation and spreading of active thrombin entering non-recalcified plasma by diffusion. Biochim. Biophys. Acta 2000, 1474, 337–345. [Google Scholar] [CrossRef]
- Lau, A.; Berry, L.R.; Mitchell, L.G.; Chan, A.K. Effect of substrate and fibrin polymerization inhibitor on determination of plasma thrombin generation in vitro. Thromb. Res. 2007, 119, 667–677. [Google Scholar] [CrossRef]
- van Berkel, S.S.; van der Lee, B.; van Delft, F.L.; Wagenvoord, R.; Hemker, H.C.; Rutjes, F.P. Fluorogenic peptide-based substrates for monitoring thrombin activity. Chem. Med. Chem. 2012, 7, 606–617. [Google Scholar] [CrossRef]
- Chatterjee, M.S.; Denney, W.S.; Jing, H.; Diamond, S.L. Systems biology of coagulation initiation: Kinetics of thrombin generation in resting and activated human blood. PLoS Comput. Biol. 2010, 6, e1000950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarandovskiy, I.D.; Balandina, A.N.; Kopylov, K.G.; Konyashina, N.I.; Kumskova, M.A.; Panteleev, M.A.; Ataullakhanov, F.I. Investigation of the phenotype heterogeneity in severe hemophilia A using thromboelastography, thrombin generation, and thrombodynamics. Thromb. Res. 2013, 131, e274–e280. [Google Scholar] [CrossRef] [PubMed]
- Tarandovskiy, I.D.; Artemenko, E.O.; Panteleev, M.A.; Sinauridze, E.I.; Ataullakhanov, F.I. Antiplatelet agents can promote two-peaked thrombin generation in platelet rich plasma: Mechanism and possible applications. PLoS ONE 2013, 8, e55688. [Google Scholar] [CrossRef] [PubMed]
- Hemker, H.C.; Beguin, S. Thrombin generation in plasma: Its assessment via the endogenous thrombin potential. Thromb. Haemost. 1995, 74, 134–138. [Google Scholar] [CrossRef] [Green Version]
- Turecek, P.L.; Varadi, K.; Keil, B.; Negrier, C.; Berntorp, E.; Astermark, J.; Bordet, J.-C.; Morfini, M.; Linari, S.; Schwarz, H.P. Factor VIII inhibitor-bypassing agents act by inducing thrombin generation and can be monitored by a thrombin generation assay. Pathophysiol. Haemost. Thromb. 2003, 33, 16–22. [Google Scholar] [CrossRef]
- Nagashima, H. Studies on the different modes of action of the anticoagulant protease inhibitors DX-9065a and Argatroban. I. Effects on thrombin generation. J. Biol. Chem. 2002, 277, 50439–50444. [Google Scholar] [CrossRef] [Green Version]
- Palmier, M.O.; Van Doren, S.R. Rapid determination of enzyme kinetics from fluorescence: Overcoming the inner filter effect. Anal. Biochem. 2007, 371, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Hemker, H.C.; Willems, G.M.; Beguin, S. A computer assisted method to obtain the prothrombin activation velocity in whole plasma independent of thrombin decay processes. Thromb. Haemost. 1986, 56, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Woodle, S.A.; Shibeko, A.M.; Lee, T.K.; Ovanesov, M.V. Determining the impact of instrument variation and automated software algorithms on the TGT in hemophilia and normalized plasma. Thromb. Res. 2013, 132, 374–380. [Google Scholar] [CrossRef]
- Varadi, K.; Negrier, C.; Berntorp, E.; Astermark, J.; Bordet, J.C.; Morfini, M.; Linari, S.; Schwarz, H.P.; Turecek, P.L. Monitoring the bioavailability of FEIBA with a thrombin generation assay. J. Thromb. Haemost. 2003, 1, 2374–2380. [Google Scholar] [CrossRef]
- Ovanesov, M.V.; Ananyeva, N.M.; Panteleev, M.A.; Ataullakhanov, F.I.; Saenko, E.L. Initiation and propagation of coagulation from tissue factor-bearing cell monolayers to plasma: Initiator cells do not regulate spatial growth rate. J. Thromb. Haemost. 2005, 3, 321–331. [Google Scholar] [CrossRef]
- Machlus, K.R.; Colby, E.A.; Wu, J.R.; Koch, G.G.; Key, N.S.; Wolberg, A.S. Effects of tissue factor, thrombomodulin and elevated clotting factor levels on thrombin generation in the calibrated automated thrombogram. Thromb. Haemost. 2009, 102, 936–944. [Google Scholar] [CrossRef] [Green Version]
- Kremers, R.M.; Mohamed, A.B.; Pelkmans, L.; Hindawi, S.; Hemker, H.C.; de Laat, H.B.; Huskens, D.; Al Dieri, R. Thrombin generating capacity and phenotypic association in ABO blood groups. PLoS ONE 2015, 10, e0141491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Veen, J.J.; Gatt, A.; Bowyer, A.E.; Cooper, P.C.; Kitchen, S.; Makris, M. Calibrated automated thrombin generation and modified thromboelastometry in haemophilia A. Thromb. Res. 2009, 123, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Luna-Zaizar, H.; Gonzalez-Moncada, A.I.; Padilla-Lopez, E.L.; Ramirez-Anguiano, A.C.; Pacheco-Moises, F.P.; Velasco-Ramirez, S.F.; Zavelia Padilla-Romo, M.G.; Borjas-Gutierrez, C.; Jaloma-Cruz, R. Thrombin generation and international normalized ratio in inherited thrombophilia patients receiving thromboprophylactic therapy. Thromb. Res. 2015, 136, 1291–1298. [Google Scholar] [CrossRef]
- Giesen, P.; van Oerle, R.; Sehgal, A.; Akinc, A.; Spronk, H.M.H. Measurement of thrombin generation in conditions of low antithrombin. In XXVI Congress of the International Society on Thrombosis and Haemostasis; International Society on Thrombosis and Haemostasis: Berlin, Germany, 2017. [Google Scholar]
- De Smedt, E.; Al Dieri, R.; Spronk, H.M.; Hamulyak, K.; ten Cate, H.; Hemker, H.C. The technique of measuring thrombin generation with fluorogenic substrates: 1. Necessity of adequate calibration. Thromb. Haemost. 2008, 100, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Chandler, W.L.; Roshal, M. Optimization of plasma fluorogenic thrombin-generation assays. Am. J. Clin. Pathol. 2009, 132, 169–179. [Google Scholar] [CrossRef] [Green Version]
- Hemker, H.C.; Hemker, P.W.; Al Dieri, R. The technique of measuring thrombin generation with fluorescent substrates: 4. The H-transform, a mathematical procedure to obtain thrombin concentrations without external calibration. Thromb. Haemost. 2009, 101, 171–177. [Google Scholar] [CrossRef] [Green Version]
- Gribkova, I.V.; Lipets, E.N.; Rekhtina, I.G.; Bernakevich, A.I.; Ayusheev, D.B.; Ovsepyan, R.A.; Ataullakhanov, F.I.; Sinauridze, E.I. The modification of the thrombin generation test for the clinical assessment of dabigatran etexilate efficiency. Sci. Rep. 2016, 6, 29242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dargaud, Y.; Wolberg, A.S.; Luddington, R.; Regnault, V.; Spronk, H.; Baglin, T.; Ten Cate, H.; Negrier, C. Evaluation of a standardized protocol for thrombin generation measurement using the calibrated automated thrombogram: An international multicentre study. Thromb. Res. 2012, 130, 929–934. [Google Scholar] [CrossRef]
- Perrin, J.; Depasse, F.; Lecompte, T. Large external quality assessment survey on thrombin generation with CAT: Further evidence for the usefulness of normalisation with an external reference plasma. Thromb. Res. 2015, 136, 125–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Paridon, P.C.S.; Panova-Noeva, M.; van Oerle, R.; Schultz, A.; Hermanns, I.M.; Prochaska, J.H.; Arnold, N.; Binder, H.; Schmidtmann, I.; Beutel, M.E.; et al. Thrombin generation in cardiovascular disease and mortality results from the Gutenberg Health Study. Haematologica 2020, 105, 2327–2334. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Artifact | Description | Potential Impact (Artifacts) of TG Curve |
|---|---|---|
| Fluorescent signal variation between instruments | Instrument-specific characteristics of fluorescent lamp and detector | Thrombin values cannot be compared between instruments |
| Fluorescent signal variation between instruments between runs | Fluorescent lamp aging may cause gradual change as instrument gets older | May increase run-to-run variability in TG values determined on the same sample of plasma on different days |
| Variable quenching of fluorescence by plasma | Occasionally, plasma composition, e.g., hemolysis, may quench fluorescence | May result in the under-estimation of TPH, velocity and AUC |
| Substrate depletion in highly procoagulant samples | Fluorogenic substrate ZGGR-AMC is gradually consumed during thrombin generation | May result in the under-estimation of AUC and, in extreme cases, TPH and TTP |
| Nonlinearity of high fluorescence signal detection (IFE) | High concentrations of AMC are underestimated because fluorescence is quenched in a thick layer of plasma | May result in the under-estimation of AUC and, in extreme cases, TPH and TTP |
| Background signal due to T-α2MG activity | Biologically inactive T-α2MG complex can cleave small peptide substrates almost as efficiently as biologically active thrombin | May result in the over-estimation of AUC and, in extreme cases, TPH |
| Competitive inhibition of thrombin by slow fluorogenic substrate | Supra-physiological concentration of fluorogenic substrate slows thrombin inhibition and prevents access of physiological substrates (e.g., fibrinogen) | Over-estimation of TG curve and potential delay in TG initiation. |
| Poor substrate specificity (e.g., cleaved by FXa and tPA) | Peptide substrate can be cleaved by a variety of coagulation enzymes | Overestimation of TG curve and distortion of TG curve shape |
| Artifact | Evaluated in This Work | References | ||||||
|---|---|---|---|---|---|---|---|---|
| Fluorescent Signal Variation between Instruments | Fluorescent Signal Variation between Runs | Variable Quenching of Fluorescence by Plasma | Substrate Consumption in Highly Procoagulant Samples | Nonlinearity of High Fluorescence Signal Detection (IFE) | ||||
| Fluorescence calibration external to sample plasma | Linear (slope ratio) calibration | Corrected by thrombin calibration coefficient 1 | Not corrected | Not corrected | Yes 2 | Turececk et al. [25] | ||
| Nonlinear calibration (slope ratio calibration, corrected for level of fluorescence 3) | Slope ratio calibration, corrected for level of fluorescence | Yes | Not tried before | |||||
| M&M formula 4 | Corrected by fluorescence calibration coefficient 1 | Corrected by M&M formula | Corrected by IFE formula 6 or calibration curve 7 | No | Nagashima [26] | |||
| M&M formula + IFE correction | Palmier & Van Doren [27] | |||||||
| Fluorescence calibration internal to sample plasma | Linear calibration | Corrected by thrombin calibration coefficient 4 | Not corrected | Yes | Not tried before | |||
| Nonlinear calibration 3 | Corrected using thrombin calibrator’s TG curve | Yes | Hemker et al. [15,16] | |||||
| M&M formula | Corrected by fluorescence calibration coefficient 1 | Corrected by M&M formula | Not corrected | Yes | Nagashima [26] | |||
| M&M formula + IFE curve 5 | Corrected by IFE calibration curve 6 | No | Tarandovskiy et al. [22,23] | |||||
| M&M formula + IFE formula 5 | Corrected by IFE formula 5 | Palmier & Van Doren [27] | ||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, W.C.; Jackson, J.W.; Machlus, K.R.; Wolberg, A.S.; Ovanesov, M.V. Comparative Analysis of Thrombin Calibration Algorithms and Correction for Thrombin-α2macroglobulin Activity. J. Clin. Med. 2020, 9, 3077. https://doi.org/10.3390/jcm9103077
Chang WC, Jackson JW, Machlus KR, Wolberg AS, Ovanesov MV. Comparative Analysis of Thrombin Calibration Algorithms and Correction for Thrombin-α2macroglobulin Activity. Journal of Clinical Medicine. 2020; 9(10):3077. https://doi.org/10.3390/jcm9103077
Chicago/Turabian StyleChang, William C., Joseph W. Jackson, Kellie R. Machlus, Alisa S. Wolberg, and Mikhail V. Ovanesov. 2020. "Comparative Analysis of Thrombin Calibration Algorithms and Correction for Thrombin-α2macroglobulin Activity" Journal of Clinical Medicine 9, no. 10: 3077. https://doi.org/10.3390/jcm9103077
APA StyleChang, W. C., Jackson, J. W., Machlus, K. R., Wolberg, A. S., & Ovanesov, M. V. (2020). Comparative Analysis of Thrombin Calibration Algorithms and Correction for Thrombin-α2macroglobulin Activity. Journal of Clinical Medicine, 9(10), 3077. https://doi.org/10.3390/jcm9103077