Novel Insights into PARK7 (DJ-1), a Potential Anti-Cancer Therapeutic Target, and Implications for Cancer Progression


Abstract
:1. Introduction
2. Role of PARK7 in Cancer Progression
3. Correlation between PARK7 Secretion and the Progression of Carcinoma
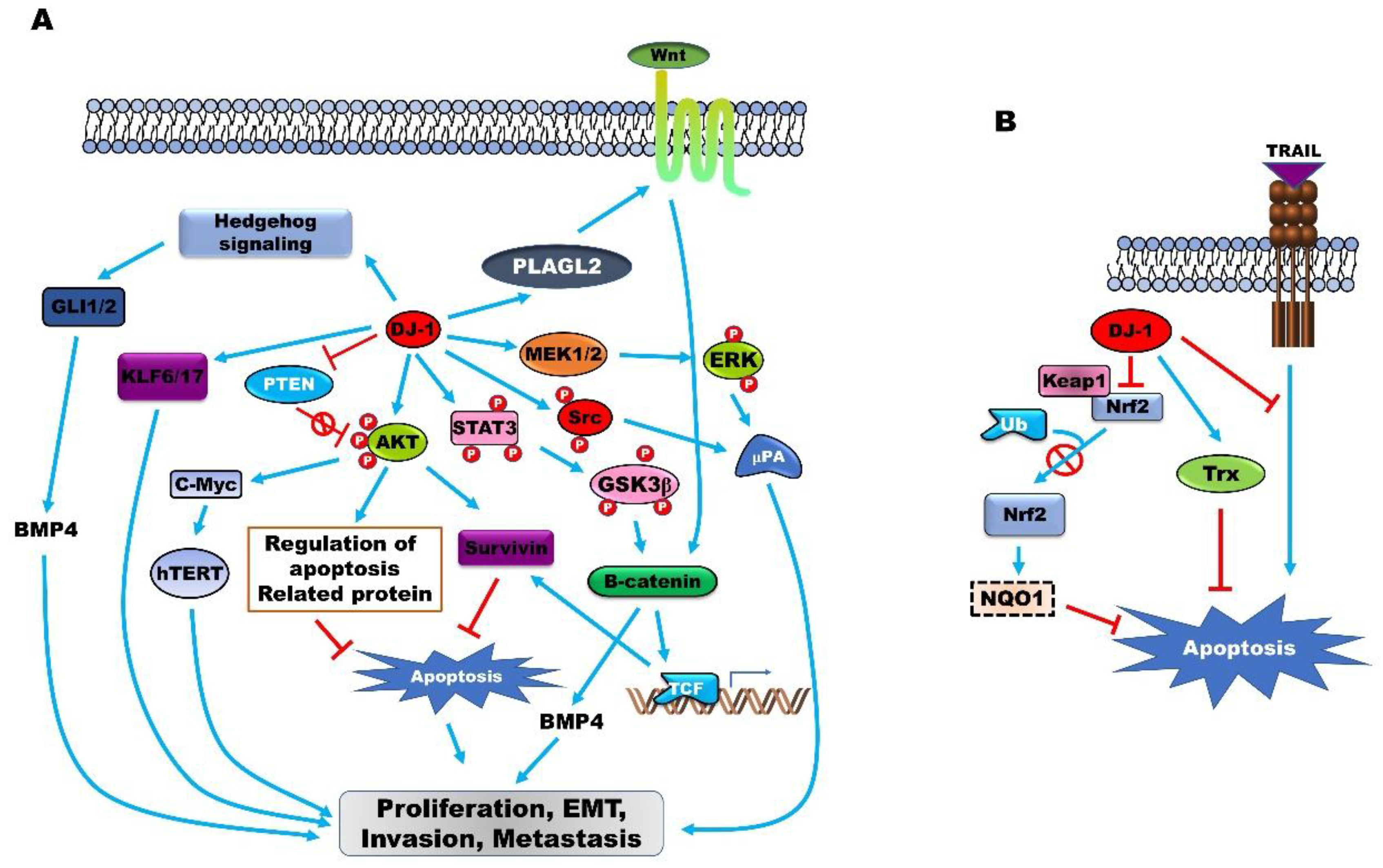
4. Functional Roles of PARK7 in Cancer Progression
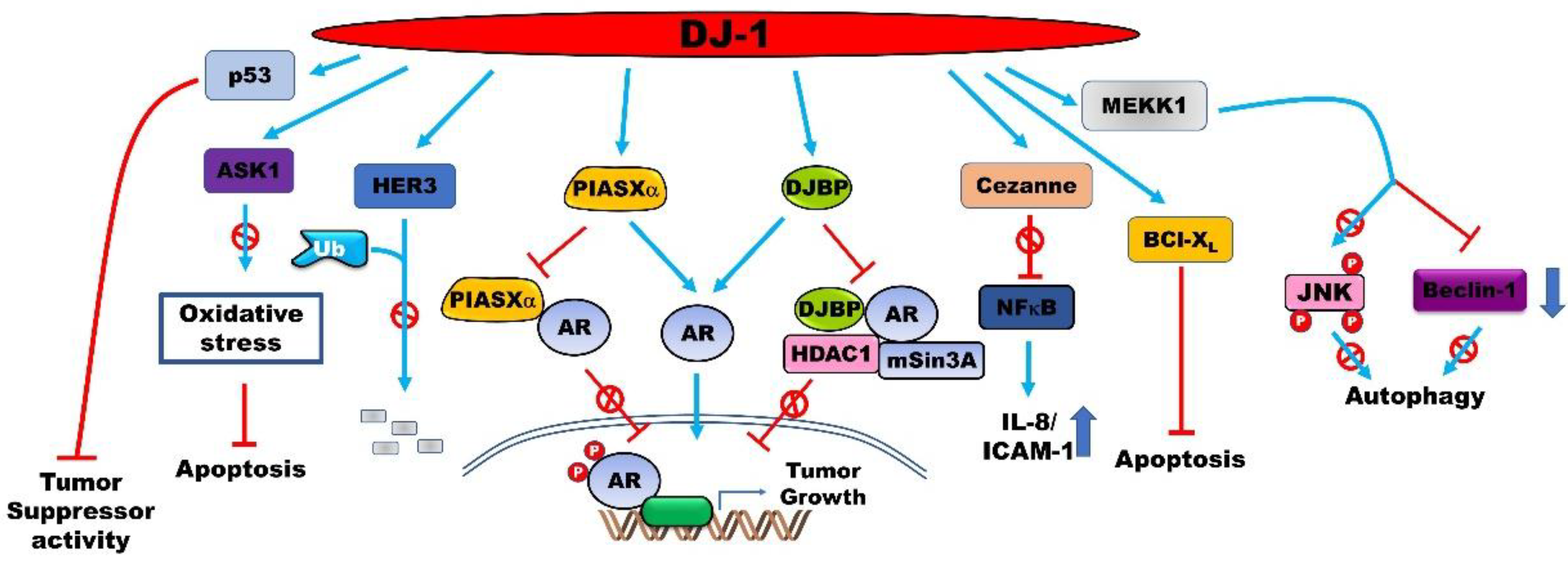
4.1. PARK7 is a Positive Regulator of the Androgen Receptor (AR)-Signaling Pathway
4.2. PARK7 Suppresses Apoptosis in Cancer Cells
4.3. PARK7 Modulates the Expression of Oncoproteins and Tumor Suppressors
4.4. PARK7 is an Oxidative-Stress Sensor
4.5. PARK7 Acts as an Orchestrator of Cellular Activity by Interacting with Various Proteins
4.6. PARK7 Acts as a Marker of Chemoresistance
5. Modulators of PARK7 Expression in Cancer Progression
5.1. Positive Modulators of PARK7 Expression
5.2. Negative Modulators of PARK7 Expression
6. Role of microRNAs (miRNAs) in the Regulation of PARK7 Expression
7. Therapies Targeting PARK7
8. Conclusions
Funding
Conflicts of Interest
References
- Smith, N.; Wilson, M.A. Structural Biology of the DJ-1 Superfamily. Adv. Exp. Med. Biol. 2017, 1037, 5–24. [Google Scholar] [CrossRef] [PubMed]
- Neumann, M.; Muller, V.; Gorner, K.; Kretzschmar, H.A.; Haass, C.; Kahle, P.J. Pathological properties of the Parkinson’s disease-associated protein DJ-1 in alpha-synucleinopathies and tauopathies: Relevance for multiple system atrophy and Pick’s disease. Acta Neuropathol. 2004, 107, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Mullett, S.J.; Hamilton, R.L.; Hinkle, D.A. DJ-1 immunoreactivity in human brain astrocytes is dependent on infarct presence and infarct age. Neuropathology 2009, 29, 125–131. [Google Scholar] [CrossRef]
- Wagenfeld, A.; Yeung, C.H.; Shivaji, S.; Sundareswaran, V.R.; Ariga, H.; Cooper, T.G. Expression and cellular localization of contraception-associated protein. J. Androl. 2000, 21, 954–963. [Google Scholar]
- Bonifati, V.; Rizzu, P.; van Baren, M.J.; Schaap, O.; Breedveld, G.J.; Krieger, E.; Dekker, M.C.; Squitieri, F.; Ibanez, P.; Joosse, M.; et al. Mutations in the DJ-1 gene associated with autosomal recessive early-onset parkinsonism. Science 2003, 299, 256–259. [Google Scholar] [CrossRef] [Green Version]
- Antipova, D.; Bandopadhyay, R. Expression of DJ-1 in Neurodegenerative Disorders. Adv. Exp. Med. Biol. 2017, 1037, 25–43. [Google Scholar] [CrossRef]
- Bonifati, V.; Rizzu, P.; Squitieri, F.; Krieger, E.; Vanacore, N.; van Swieten, J.C.; Brice, A.; van Duijn, C.M.; Oostra, B.; Meco, G.; et al. DJ-1( PARK7), a novel gene for autosomal recessive, early onset parkinsonism. Neurol. Sci. 2003, 24, 159–160. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Petrie, T.G.; Liu, Y.; Liu, J.; Fujioka, H.; Zhu, X. Parkinson’s disease-associated DJ-1 mutations impair mitochondrial dynamics and cause mitochondrial dysfunction. J. Neurochem. 2012, 121, 830–839. [Google Scholar] [CrossRef]
- Repici, M.; Giorgini, F. DJ-1 in Parkinson’s Disease: Clinical Insights and Therapeutic Perspectives. J. Clin. Med. 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Abou-Sleiman, P.M.; Healy, D.G.; Quinn, N.; Lees, A.J.; Wood, N.W. The role of pathogenic DJ-1 mutations in Parkinson’s disease. Ann. Neurol. 2003, 54, 283–286. [Google Scholar] [CrossRef]
- Bonifati, V.; Oostra, B.A.; Heutink, P. Linking DJ-1 to neurodegeneration offers novel insights for understanding the pathogenesis of Parkinson’s disease. J. Mol. Med. 2004, 82, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Hague, S.; Rogaeva, E.; Hernandez, D.; Gulick, C.; Singleton, A.; Hanson, M.; Johnson, J.; Weiser, R.; Gallardo, M.; Ravina, B.; et al. Early-onset Parkinson’s disease caused by a compound heterozygous DJ-1 mutation. Ann. Neurol. 2003, 54, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Taipa, R.; Pereira, C.; Reis, I.; Alonso, I.; Bastos-Lima, A.; Melo-Pires, M.; Magalhaes, M. DJ-1 linked parkinsonism (PARK7) is associated with Lewy body pathology. Brain 2016, 139, 1680–1687. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, C.P.; Giasson, B.I. L10P and P158DEL DJ-1 Mutations Cause Protein Instability, Aggregation, and Dimerization Impairments. J. Neurosci. Res. 2010, 88, 3111–3124. [Google Scholar] [CrossRef] [Green Version]
- Moore, D.J.; Zhang, L.; Dawson, T.M.; Dawson, V.L. A missense mutation (L166P) in DJ-1, linked to familial Parkinson’s disease, confers reduced protein stability and impairs homo-oligomerization. J. Neurochem. 2003, 87, 1558–1567. [Google Scholar] [CrossRef]
- Olzmann, J.A.; Brown, K.; Wilkinson, K.D.; Rees, H.D.; Huai, Q.; Ke, H.; Levey, A.I.; Li, L.; Chin, L.S. Familial Parkinson’s disease-associated L166P mutation disrupts DJ-1 protein folding and function. J. Biol. Chem. 2004, 279, 8506–8515. [Google Scholar] [CrossRef] [Green Version]
- Giaime, E.; Sunyach, C.; Druon, C.; Scarzello, S.; Robert, G.; Grosso, S.; Auberger, P.; Goldberg, M.S.; Shen, J.; Heutink, P.; et al. Loss of function of DJ-1 triggered by Parkinson’s disease-associated mutation is due to proteolytic resistance to caspase-6. Cell Death Differ. 2010, 17, 158–169. [Google Scholar] [CrossRef] [Green Version]
- Nural, H.; He, P.; Beach, T.; Sue, L.; Xia, W.M.; Shen, Y. Dissembled DJ-1 high molecular weight complex in cortex mitochondria from Parkinson’s disease patients. Mol. Neurodegener. 2009, 4. [Google Scholar] [CrossRef] [Green Version]
- Meulener, M.C.; Graves, C.L.; Sampathu, D.M.; Armstrong-Gold, C.E.; Bonini, N.M.; Giasson, B.I. DJ-1 is present in a large molecular complex in human brain tissue and interacts with alpha-synuclein. J. Neurochem. 2005, 93, 1524–1532. [Google Scholar] [CrossRef]
- Solti, K.; Kuan, W.L.; Forizs, B.; Kustos, G.; Mihaly, J.; Varga, Z.; Herberth, B.; Moravcsik, E.; Kiss, R.; Karpati, M.; et al. DJ-1 can form beta-sheet structured aggregates that co-localize with pathological amyloid deposits. Neurobiol. Dis. 2019, 134, 104629. [Google Scholar] [CrossRef]
- Zhu, M.; Patel, S.H.; Han, S. DJ-1, a Parkinson’s disease related protein, aggregates under denaturing conditions and co-aggregates with alpha-synuclein through hydrophobic interaction. BBA Gen. Subj. 2017, 1861, 1759–1769. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Lee, S.; Yun, S.; Suh, P.G.; Park, J.; Cui, M.H.; Choi, S.; Cha, S.S.; Jin, W. Inhibitory effect of tartrate against phosphate-induced DJ-1 aggregation. Int. J. Biol. Macromol. 2018, 107, 1650–1658. [Google Scholar] [CrossRef] [PubMed]
- Herrera, F.E.; Zucchelli, S.; Jezierska, A.; Lavina, Z.S.; Gustincich, S.; Carloni, P. On the oligomeric state of DJ-1 protein and its mutants associated with Parkinson Disease. A combined computational and in vitro study. J. Biol. Chem. 2007, 282, 24905–24914. [Google Scholar] [CrossRef] [Green Version]
- Shendelman, S.; Jonason, A.; Martinat, C.; Leete, T.; Abeliovich, A. DJ-1 is a redox-dependent molecular chaperone that inhibits alpha-synuclein aggregate formation. PLoS Biol. 2004, 2, 1764–1773. [Google Scholar] [CrossRef] [Green Version]
- Nagakubo, D.; Taira, T.; Kitaura, H.; Ikeda, M.; Tamai, K.; Iguchi-Ariga, S.M.; Ariga, H. DJ-1, a novel oncogene which transforms mouse NIH3T3 cells in cooperation with ras. Biochem. Biophys. Res. Commun. 1997, 231, 509–513. [Google Scholar] [CrossRef]
- Abd El Atti, R.M.; Abou Gabal, H.H.; Osman, W.M.; Saad, A.S. Insights into the prognostic value of DJ-1 and MIB-1 in astrocytic tumors. Diagn. Pathol. 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Hinkle, D.A.; Mullett, S.J.; Gabris, B.E.; Hamilton, R.L. DJ-1 expression in glioblastomas shows positive correlation with p53 expression and negative correlation with epidermal growth factor receptor amplification. Neuropathology 2011, 31, 29–37. [Google Scholar] [CrossRef]
- Lin, J.P.; Pan, B.C.; Li, B.; Li, Y.; Tian, X.Y.; Li, Z. DJ-1 is activated in medulloblastoma and is associated with cell proliferation and differentiation. World J. Surg. Oncol. 2014, 12, 373. [Google Scholar] [CrossRef] [Green Version]
- Kim, R.H.; Peters, M.; Jang, Y.; Shi, W.; Pintilie, M.; Fletcher, G.C.; DeLuca, C.; Liepa, J.; Zhou, L.; Snow, B.; et al. DJ-1, a novel regulator of the tumor suppressor PTEN. Cancer Cell 2005, 7, 263–273. [Google Scholar] [CrossRef] [Green Version]
- MacKeigan, J.P.; Clements, C.M.; Lich, J.D.; Pope, R.M.; Hod, Y.; Ting, J.P. Proteomic profiling drug-induced apoptosis in non-small cell lung carcinoma: Identification of RS/DJ-1 and RhoGDIalpha. Cancer Res. 2003, 63, 6928–6934. [Google Scholar]
- Bai, J.; Guo, C.; Sun, W.; Li, M.; Meng, X.; Yu, Y.; Jin, Y.; Tong, D.; Geng, J.; Huang, Q.; et al. DJ-1 may contribute to metastasis of non-small cell lung cancer. Mol. Biol. Rep. 2012, 39, 2697–2703. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Y.; Wang, H.Q.; Liu, H.M.; Guan, Y.; Du, Z.X. Regulation of tumor necrosis factor-related apoptosis-inducing ligand-induced apoptosis by DJ-1 in thyroid cancer cells. Endocr. Relat. Cancer 2008, 15, 535–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Zheng, Z.; Li, J.; Ben, Q.; Liu, J.; Zhang, J.; Ji, J.; Yu, B.; Chen, X.; Su, L.; et al. DJ-1 promotes invasion and metastasis of pancreatic cancer cells by activating SRC/ERK/uPA. Carcinogenesis 2012, 33, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Fang, B.; Yan, S.; Wang, W. Comparison of Tumor Markers for Predicting Disease-Free Survival in Surgically Resected Pancreatic Neuroendocrine Tumors. Clin. Lab. 2017, 63, 1251–1259. [Google Scholar] [CrossRef]
- Wu, F.; Liang, Y.Q.; Huang, Z.M. The expression of DJ-1 gene in human hepatocellular carcinoma and its relationship with tumor invasion and metastasis. Zhonghua Gan Zang Bing Za Zhi 2009, 17, 203–206. [Google Scholar]
- Abdalla, M.A.; Haj-Ahmad, Y. Promising Urinary Protein Biomarkers for the Early Detection of Hepatocellular Carcinoma among High-Risk Hepatitis C Virus Egyptian Patients. J. Cancer 2012, 3, 390–403. [Google Scholar] [CrossRef] [Green Version]
- Davidson, B.; Hadar, R.; Schlossberg, A.; Sternlicht, T.; Slipicevic, A.; Skrede, M.; Risberg, B.; Florenes, V.A.; Kopolovic, J.; Reich, R. Expression and clinical role of DJ-1, a negative regulator of PTEN, in ovarian carcinoma. Hum. Pathol. 2008, 39, 87–95. [Google Scholar] [CrossRef]
- Yuen, H.F.; Chan, Y.P.; Law, S.; Srivastava, G.; El-Tanani, M.; Mak, T.W.; Chan, K.W. DJ-1 could predict worse prognosis in esophageal squamous cell carcinoma. Cancer Epidemiol. Biomark. Prev. 2008, 17, 3593–3602. [Google Scholar] [CrossRef] [Green Version]
- Kawase, H.; Fujii, K.; Miyamoto, M.; Kubota, K.C.; Hirano, S.; Kondo, S.; Inagaki, F. Differential LC-MS-based proteomics of surgical human cholangiocarcinoma tissues. J. Proteome. Res. 2009, 8, 4092–4103. [Google Scholar] [CrossRef]
- Zhu, X.L.; Wang, Z.F.; Lei, W.B.; Zhuang, H.W.; Jiang, H.Y.; Wen, W.P. DJ-1: A novel independent prognostic marker for survival in glottic squamous cell carcinoma. Cancer Sci. 2010, 101, 1320–1325. [Google Scholar] [CrossRef]
- Merikallio, H.; Paakko, P.; Kinnula, V.L.; Harju, T.; Soini, Y. Nuclear factor erythroid-derived 2-like 2 (Nrf2) and DJ1 are prognostic factors in lung cancer. Hum. Pathol. 2012, 43, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Ciregia, F.; Giusti, L.; Molinaro, A.; Niccolai, F.; Mazzoni, M.R.; Rago, T.; Tonacchera, M.; Vitti, P.; Giannaccini, G.; Lucacchini, A. Proteomic analysis of fine-needle aspiration in differential diagnosis of thyroid nodules. Transl. Res. 2016, 176, 81–94. [Google Scholar] [CrossRef]
- Hod, Y. Differential control of apoptosis by DJ-1 in prostate benign and cancer cells. J. Cell Biochem. 2004, 92, 1221–1233. [Google Scholar] [CrossRef] [PubMed]
- Pardo, M.; Garcia, A.; Thomas, B.; Pineiro, A.; Akoulitchev, A.; Dwek, R.A.; Zitzmann, N. The characterization of the invasion phenotype of uveal melanoma tumour cells shows the presence of MUC18 and HMG-1 metastasis markers and leads to the identification of DJ-1 as a potential serum biomarker. Int. J. Cancer 2006, 119, 1014–1022. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.L.; Tian, J.J.; Su, L.; Jing, Y.; Zhang, S.C.; Zhang, H.X.; Wang, X.Q.; Zhu, C.B. DJ-1: A promising marker in metastatic uveal melanoma. J. Cancer Res. Clin. Oncol. 2015, 141, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Melle, C.; Ernst, G.; Escher, N.; Hartmann, D.; Schimmel, B.; Bleul, A.; Thieme, H.; Kaufmann, R.; Felix, K.; Friess, H.M.; et al. Protein profiling of microdissected pancreas carcinoma and identification of HSP27 as a potential serum marker. Clin. Chem. 2007, 53, 629–635. [Google Scholar] [CrossRef] [Green Version]
- Tian, M.; Cui, Y.Z.; Song, G.H.; Zong, M.J.; Zhou, X.Y.; Chen, Y.; Han, J.X. Proteomic analysis identifies MMP-9, DJ-1 and A1BG as overexpressed proteins in pancreatic juice from pancreatic ductal adenocarcinoma patients. BMC Cancer 2008, 8, 241. [Google Scholar] [CrossRef] [Green Version]
- He, X.Y.; Liu, B.Y.; Yao, W.Y.; Zhao, X.J.; Zheng, Z.; Li, J.F.; Yu, B.Q.; Yuan, Y.Z. Serum DJ-1 as a diagnostic marker and prognostic factor for pancreatic cancer. J. Dig. Dis. 2011, 12, 131–137. [Google Scholar] [CrossRef]
- Qiu, B.; Wang, J.; Yu, Y.; Zhen, C.; Gu, J.; Liu, W.; Wen, Y.; Chen, L.; Gao, Y.; Xia, Q.; et al. DJ-1 promotes development of DEN-induced hepatocellular carcinoma and proliferation of liver cancer cells. Oncotarget 2017, 8, 8499–8511. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Long, G.; Wei, H.; Shi, L.; Yang, Z.; Liu, D.; Hu, G.; Qiu, H. DJ-1 knockdown inhibits growth and xenograftinduced tumor generation of human hepatocellular carcinoma cells. Oncol. Rep. 2015, 33, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Bunkholt Elstrand, M.; Dong, H.P.; Odegaard, E.; Holth, A.; Elloul, S.; Reich, R.; Trope, C.G.; Davidson, B. Mammalian target of rapamycin is a biomarker of poor survival in metastatic serous ovarian carcinoma. Hum. Pathol. 2010, 41, 794–804. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, M.; Li, M.; Wang, D.; Rao, Q.; Wang, Y.; Xu, Z.; Wang, J. Expression and role of DJ-1 in leukemia. Biochem. Biophys. Res. Commun. 2008, 375, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Raninga, P.V.; Di Trapani, G.; Vuckovic, S.; Tonissen, K.F. Targeted knockdown of DJ-1 induces multiple myeloma cell death via KLF6 upregulation. Apoptosis 2016, 21, 1422–1437. [Google Scholar] [CrossRef] [PubMed]
- Sitaram, R.T.; Cairney, C.J.; Grabowski, P.; Keith, W.N.; Hallberg, B.; Ljungberg, B.; Roos, G. The PTEN regulator DJ-1 is associated with hTERT expression in clear cell renal cell carcinoma. Int. J. Cancer 2009, 125, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Arnouk, H.; Merkley, M.A.; Podolsky, R.H.; Stoppler, H.; Santos, C.; Alvarez, M.; Mariategui, J.; Ferris, D.; Lee, J.R.; Dynan, W.S. Characterization of Molecular Markers Indicative of Cervical Cancer Progression. Proteom. Clin. Appl. 2009, 3, 516–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Gao, W.W. DJ-1 Expression in Cervical Carcinoma and its Effects on Cell Viability and Apoptosis. Med. Sci. Monit. 2016, 22. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.L.; Wen, W.P.; Lei, W.B.; Chai, L.P.; Hou, W.J.; Wen, Y.H.; Wang, X.R. DJ-1 expression in laryngeal squamous cell carcinoma and its relationship with tumor recurrence and metastasis. Zhonghua Er Bi Yan Hou Tou Jing Wai Ke Za Zhi 2010, 45, 497–501. [Google Scholar]
- Shen, Z.; Ren, Y.; Ye, D.; Guo, J.; Kang, C.; Ding, H. Significance and relationship between DJ-1 gene and surviving gene expression in laryngeal carcinoma. Eur. J. Histochem. 2011, 55, e9. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.L.; Sun, W.; Lei, W.B.; Zhuang, H.W.; Hou, W.J.; Wen, W.P. DJ-1-induced phosphatase and tensin homologue downregulation is associated with proliferative and invasive activity of laryngeal cancer cells. Mol. Med. Rep. 2015, 12, 2003–2008. [Google Scholar] [CrossRef]
- Shu, K.Y.; Xiao, Z.Q.; Long, S.G.; Yan, J.J.; Yu, X.H.; Zhu, Q.Z.; Mei, T. Expression of DJ-1 in Endometrial Cancer Close Correlation With Clinicopathological Features and Apoptosis. Int. J. Gynecol. Cancer 2013, 23, 1029–1035. [Google Scholar] [CrossRef]
- Morelli, M.; Scumaci, D.; Di Cello, A.; Venturella, R.; Donato, G.; Faniello, M.C.; Quaresima, B.; Cuda, G.; Zullo, F.; Costanzo, F. DJ-1 in Endometrial Cancer A Possible Biomarker to Improve Differential Diagnosis Between Subtypes. Int. J. Gynecol. Cancer 2014, 24, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Gao, J.; Yu, X.; Luo, H.; Xiong, X.; Huang, O. Expression of DJ-1 and mTOR in eutopic and ectopic endometria of patients with endometriosis and adenomyosis. Gynecol. Obs. Investig. 2015, 79, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Kuramitsu, Y.; Baron, B.; Yoshino, S.; Zhang, X.; Tanaka, T.; Yashiro, M.; Hirakawa, K.; Oka, M.; Nakamura, K. Proteomic differential display analysis shows up-regulation of 14-3-3 sigma protein in human scirrhous-type gastric carcinoma cells. Anticancer Res. 2010, 30, 4459–4465. [Google Scholar] [PubMed]
- Alvarez-Chaver, P.; Rodriguez-Pineiro, A.M.; Rodriguez-Berrocal, F.J.; Garcia-Lorenzo, A.; Paez de la Cadena, M.; Martinez-Zorzano, V.S. Selection of putative colorectal cancer markers by applying PCA on the soluble proteome of tumors: NDK A as a promising candidate. J. Proteom. 2011, 74, 874–886. [Google Scholar] [CrossRef]
- Lei, Y.; Huang, K.; Gao, C.; Lau, Q.C.; Pan, H.; Xie, K.; Li, J.; Liu, R.; Zhang, T.; Xie, N.; et al. Proteomics identification of ITGB3 as a key regulator in reactive oxygen species-induced migration and invasion of colorectal cancer cells. Mol. Cell Proteom. 2011, 10, M110005397. [Google Scholar] [CrossRef] [Green Version]
- Phueaouan, T.; Chaiyawat, P.; Netsirisawan, P.; Chokchaichamnankit, D.; Punyarit, P.; Srisomsap, C.; Svasti, J.; Champattanachai, V. Aberrant O-GlcNAc-modified proteins expressed in primary colorectal cancer. Oncol. Rep. 2013, 30, 2929–2936. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Chen, Q.; Liu, Q.X.; Zhou, D.; Lu, X.; Deng, X.F.; Yang, H.; Zheng, H.; Qiu, Y. High expression of DJ-1 promotes growth and invasion via the PTEN-AKT pathway and predicts a poor prognosis in colorectal cancer. Cancer Med. 2018, 7, 809–819. [Google Scholar] [CrossRef]
- Zhou, J.; Liu, H.; Zhang, L.; Liu, X.; Zhang, C.D.; Wang, Y.T.; He, Q.; Zhang, Y.; Li, Y.; Chen, Q.M.; et al. DJ-1 promotes colorectal cancer progression through activating PLAGL2/Wnt/BMP4 axis. Cell Death Dis. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Cui, J.; Zhang, C.H.; Yang, D.J.; Chen, J.H.; Zan, W.H.; Li, B.; Li, Z.; He, Y.L. High-expression of DJ-1 and loss of PTEN associated with tumor metastasis and correlated with poor prognosis of gastric carcinoma. Int. J. Med. Sci. 2013, 10, 1689–1697. [Google Scholar] [CrossRef] [Green Version]
- Tsuchiya, B.; Iwaya, K.; Kohno, N.; Kawate, T.; Akahoshi, T.; Matsubara, O.; Mukai, K. Clinical significance of DJ-1 as a secretory molecule: Retrospective study of DJ-1 expression at mRNA and protein levels in ductal carcinoma of the breast. Histopathology 2012, 61, 69–77. [Google Scholar] [CrossRef]
- Kawate, T.; Iwaya, K.; Kikuchi, R.; Kaise, H.; Oda, M.; Sato, E.; Hiroi, S.; Matsubara, O.; Kohno, N. DJ-1 protein expression as a predictor of pathological complete remission after neoadjuvant chemotherapy in breast cancer patients. Breast Cancer Res. Tr. 2013, 139, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Kawate, T.; Iwaya, K.; Koshikawa, K.; Moriya, T.; Yamasaki, T.; Hasegawa, S.; Kaise, H.; Fujita, T.; Matsuo, H.; Nakamura, T.; et al. High levels of DJ-1 protein and isoelectric point 6.3 isoform in sera of breast cancer patients. Cancer Sci. 2015, 106, 938–943. [Google Scholar] [CrossRef]
- Fan, J.P.; Yu, H.Y.; Lv, Y.; Yin, L.G. Diagnostic and prognostic value of serum thioredoxin and DJ-1 in non-small cell lung carcinoma patients. Tumor. Biol. 2016, 37, 1949–1958. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Wang, J.; Gao, J.; Feng, S.; Zhu, Y.; Li, X.; Xiao, T.; Qi, J.; Cui, W. DJ-1 as a potential biomarker for the early diagnosis in lung cancer patients. Tumour Biol. 2017, 39, 1010428317714625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabata, Y.; Nakanishi, Y.; Hatanaka, K.C.; Hatanaka, Y.; Tsuchikawa, T.; Okamura, K.; Noji, T.; Shichinohe, T.; Matsuno, Y.; Hirano, S. DJ-1 is a useful biomarker for invasive extrahepatic cholangiocarcinoma. Hum. Pathol. 2018, 76, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Di Cello, A.; Di Sanzo, M.; Perrone, F.M.; Santamaria, G.; Rania, E.; Angotti, E.; Venturella, R.; Mancuso, S.; Zullo, F.; Cuda, G.; et al. DJ-1 is a reliable serum biomarker for discriminating high-risk endometrial cancer. Tumour Biol. 2017, 39, 1010428317705746. [Google Scholar] [CrossRef] [Green Version]
- Shimwell, N.J.; Ward, D.G.; Mohri, Y.; Mohri, T.; Pallan, L.; Teng, M.; Miki, Y.C.; Kusunoki, M.; Tucker, O.; Wei, W.; et al. Macrophage migration inhibitory factor and DJ-1 in gastric cancer: Differences between high-incidence and low-incidence areas. Brit. J. Cancer 2012, 107, 1595–1601. [Google Scholar] [CrossRef]
- Chen, Y.; Kang, M.; Lu, W.; Guo, Q.; Zhang, B.; Xie, Q.; Wu, Y. DJ-1, a novel biomarker and a selected target gene for overcoming chemoresistance in pancreatic cancer. J. Cancer Res. Clin. Oncol. 2012, 138, 1463–1474. [Google Scholar] [CrossRef]
- Le Naour, F.; Misek, D.E.; Krause, M.C.; Deneux, L.; Giordano, T.J.; Scholl, S.; Hanash, S.M. Proteomics-based identification of RS/DJ-1 as a novel circulating tumor antigen in breast cancer. Clin. Cancer Res. 2001, 7, 3328–3335. [Google Scholar]
- Oda, M.; Makita, M.; Iwaya, K.; Akiyama, F.; Kohno, N.; Tsuchiya, B.; Iwase, T.; Matsubara, O. High levels of DJ-1 protein in nipple fluid of patients with breast cancer. Cancer Sci. 2012, 103, 1172–1176. [Google Scholar] [CrossRef]
- Balk, S.P. Androgen receptor as a target in androgen-independent prostate cancer. Urology 2002, 60, 132–138. [Google Scholar] [CrossRef]
- Taplin, M.E.; Bubley, G.J.; Ko, Y.J.; Small, E.J.; Upton, M.; Rajeshkumar, B.; Balk, S.P. Selection for androgen receptor mutations in prostate cancers treated with androgen antagonist. Cancer Res. 1999, 59, 2511–2515. [Google Scholar] [PubMed]
- Takahashi, K.; Taira, T.; Niki, T.; Seino, C.; Iguchi-Ariga, S.M.; Ariga, H. DJ-1 positively regulates the androgen receptor by impairing the binding of PIASx alpha to the receptor. J. Biol. Chem. 2001, 276, 37556–37563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niki, T.; Takahashi-Niki, K.; Taira, T.; Iguchi-Ariga, S.M.M.; Ariga, H. DJBP: A novel DJ-1-binding protein, negatively regulates the androgen receptor by recruiting histone deacetylase complex, and DJ-1 antagonizes this inhibition by abrogation of this complex. Mol. Cancer Res. 2003, 1, 247–261. [Google Scholar]
- Shen, Z.; Jiang, Z.; Ye, D.; Xiao, B.; Zhang, X.; Guo, J. Growth inhibitory effects of DJ-1-small interfering RNA on laryngeal carcinoma Hep-2 cells. Med. Oncol. 2011, 28, 601–607. [Google Scholar] [CrossRef]
- Ren, H.; Fu, K.; Mu, C.; Li, B.; Wang, D.; Wang, G. DJ-1, a cancer and Parkinson’s disease associated protein, regulates autophagy through JNK pathway in cancer cells. Cancer Lett. 2010, 297, 101–108. [Google Scholar] [CrossRef]
- Sun, H.; Lesche, R.; Li, D.M.; Liliental, J.; Zhang, H.; Gao, J.; Gavrilova, N.; Mueller, B.; Liu, X.; Wu, H. PTEN modulates cell cycle progression and cell survival by regulating phosphatidylinositol 3,4,5,-trisphosphate and Akt/protein kinase B signaling pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 6199–6204. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Fang, M.; Zhang, M.; Li, W.P.; Guan, H.; Sun, Y.H.; Xie, S.M.; Zhong, X.Y. The positive correlation between DJ-1 and beta-catenin expression shows prognostic value for patients with glioma. Neuropathology 2013, 33, 628–636. [Google Scholar] [CrossRef]
- Kim, Y.C.; Kitaura, H.; Taira, T.; Iguchi-Ariga, S.M.; Ariga, H. Oxidation of DJ-1-dependent cell transformation through direct binding of DJ-1 to PTEN. Int. J. Oncol. 2009, 35, 1331–1341. [Google Scholar]
- Klawitter, J.; Shokati, T.; Moll, V.; Christians, U.; Klawitter, J. Effects of lovastatin on breast cancer cells: A proteo-metabonomic study. Breast Cancer Res. 2010, 12, R16. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.C.; Kitaura, H.; Iguchi-Ariga, S.M.; Ariga, H. DJ-1, an oncogene and causative gene for familial Parkinson’s disease, is essential for SV40 transformation in mouse fibroblasts through up-regulation of c-Myc. FEBS Lett. 2010, 584, 3891–3895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismail, I.A.; Kang, H.S.; Lee, H.J.; Kim, J.K.; Hong, S.H. DJ-1 upregulates breast cancer cell invasion by repressing KLF17 expression. Brit. J. Cancer 2014, 110, 1298–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, L.S.; Huang, R.H.; Lai, H.W.; Hsu, H.T.; Sung, W.W.; Hsieh, M.J.; Wu, C.Y.; Lin, Y.M.; Chen, M.K.; Lo, Y.S.; et al. KLF6 inhibited oral cancer migration and invasion via downregulation of mesenchymal markers and inhibition of MMP-9 activities. Int. J. Med. Sci. 2017, 14, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Clements, C.M.; McNally, R.S.; Conti, B.J.; Mak, T.W.; Ting, J.P. DJ-1, a cancer-and Parkinson’s disease-associated protein, stabilizes the antioxidant transcriptional master regulator Nrf2. Proc. Natl. Acad. Sci. USA 2006, 103, 15091–15096. [Google Scholar] [CrossRef] [Green Version]
- Motani, K.; Tabata, K.; Kimura, Y.; Okano, S.; Shibata, Y.; Abiko, Y.; Nagai, H.; Akihisa, T.; Suzuki, T. Proteomic analysis of apoptosis induced by xanthoangelol, a major constituent of Angelica keiskei, in neuroblastoma. Biol. Pharm. Bull. 2008, 31, 618–626. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Sullards, M.C.; Olzmann, J.A.; Rees, H.D.; Weintraub, S.T.; Bostwick, D.E.; Gearing, M.; Levey, A.I.; Chin, L.S.; Li, L. Oxidative damage of DJ-1 is linked to sporadic Parkinson and Alzheimer diseases. J. Biol. Chem. 2006, 281, 10816–10824. [Google Scholar] [CrossRef] [Green Version]
- Fu, C.; Wu, C.; Liu, T.; Ago, T.; Zhai, P.; Sadoshima, J.; Li, H. Elucidation of thioredoxin target protein networks in mouse. Mol. Cell Proteom. 2009, 8, 1674–1687. [Google Scholar] [CrossRef] [Green Version]
- Tillman, J.E.; Yuan, J.; Gu, G.; Fazli, L.; Ghosh, R.; Flynt, A.S.; Gleave, M.; Rennie, P.S.; Kasper, S. DJ-1 binds androgen receptor directly and mediates its activity in hormonally treated prostate cancer cells. Cancer Res. 2007, 67, 4630–4637. [Google Scholar] [CrossRef] [Green Version]
- Ren, H.; Fu, K.; Wang, D.; Mu, C.; Wang, G. Oxidized DJ-1 interacts with the mitochondrial protein BCL-XL. J. Biol. Chem. 2011, 286, 35308–35317. [Google Scholar] [CrossRef] [Green Version]
- McNally, R.S.; Davis, B.K.; Clements, C.M.; Accavitti-Loper, M.A.; Mak, T.W.; Ting, J.P. DJ-1 enhances cell survival through the binding of Cezanne, a negative regulator of NF-kappaB. J. Biol. Chem. 2011, 286, 4098–4106. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Mukherjee, S.; Fan, X.J.; Salameh, A.; Mujoo, K.; Huang, Z.; Li, L.K.; Salazar, G.T.; Zhang, N.Y.; An, Z.Q. Novel association of DJ-1 with HER3 potentiates HER3 activation and signaling in cancer. Oncotarget 2016, 7, 65758–65769. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Lu, Q.; Wang, Y.; Sun, X.; Zhang, S. NRG-1 Stimulates Serum DJ-1 Increase in Breast Cancers. Pathol. Oncol. Res. 2019, 25, 71–79. [Google Scholar] [CrossRef]
- Cao, J.; Ying, M.; Xie, N.; Lin, G.; Dong, R.; Zhang, J.; Yan, H.; Yang, X.; He, Q.; Yang, B. The oxidation states of DJ-1 dictate the cell fate in response to oxidative stress triggered by 4-hpr: Autophagy or apoptosis? Antioxid. Redox. Signal 2014, 21, 1443–1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitkanen-Arsiola, T.; Tillman, J.E.; Gu, G.; Yuan, J.; Roberts, R.L.; Wantroba, M.; Coetzee, G.A.; Cookson, M.S.; Kasper, S. Androgen and anti-androgen treatment modulates androgen receptor activity and DJ-1 stability. Prostate 2006, 66, 1177–1193. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.Z.; Qu, Y.Q.; Zhang, W.J.; Xiu, B.; Deng, A.M.; Liang, A.B. Proteomic analysis identified DJ-1 as a cisplatin resistant marker in non-small cell lung cancer. Int. J. Mol. Sci. 2011, 12, 3489–3499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.Y.; Niu, Y.C.; Li, M.; Fang, S.; Guo, L.L. Identification of DJ-1 as a contributor to multidrug resistance in human small-cell lung cancer using proteomic analysis. Int. J. Exp. Pathol. 2017, 98, 67–74. [Google Scholar] [CrossRef]
- Trivedi, R.; Dihazi, G.H.; Eltoweissy, M.; Mishra, D.P.; Mueller, G.A.; Dihazi, H. The antioxidant protein PARK7 plays an important role in cell resistance to Cisplatin-induced apoptosis in case of clear cell renal cell carcinoma. Eur. J. Pharm. 2016, 784, 99–110. [Google Scholar] [CrossRef]
- Pan, X.K.; Su, F.; Xu, L.H.; Yang, Z.S.; Wang, D.W.; Yang, L.J.; Kong, F.Z.; Xie, W.; Feng, M.H. DJ-1 Alters Epirubicin-induced Apoptosis via Modulating Epirubicinactivated Autophagy in Human Gastric Cancer Cells. Curr. Med. Sci. 2018, 38, 1018–1024. [Google Scholar] [CrossRef]
- Liu, H.Y.; Duan, G.L.; Xu, R.Y.; Li, X.R.; Xiao, L.; Zhao, L.; Ma, Z.X.; Xu, X.W.; Qiu, L.J.; Zhu, Z.M.; et al. DJ-1 overexpression confers the multidrug resistance phenotype to SGC7901 cells by upregulating P-gp and Bcl-2. Biochem. Bioph. Res. Co. 2019, 519, 73–80. [Google Scholar] [CrossRef]
- Zhang, G.Q.; He, C.; Tao, L.; Liu, F. Role of DJ-1 siRNA in reverse sensitivity of breast cancer cells to chemotherapy and its possible mechanism. Int. J. Clin. Exp. Patho. 2015, 8, 6944–6951. [Google Scholar]
- Feng, X.; Li, L.; Jiang, H.; Jiang, K.; Jin, Y.; Zheng, J. Dihydroartemisinin potentiates the anticancer effect of cisplatin via mTOR inhibition in cisplatin-resistant ovarian cancer cells: Involvement of apoptosis and autophagy. Biochem. Biophys. Res. Commun. 2014, 444, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Liao, S.D.; Shi, J.J.; Chang, L.L.; Tong, Y.G.; Cao, J.; Fu, Y.Y.; Chen, X.P.; Ying, M.D.; Yang, B.; et al. DJ-1 mediates the resistance of cancer cells to dihydroartemisinin through reactive oxygen species removal. Free Radic. Biol. Med. 2014, 71, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Sekito, A.; Taira, T.; Niki, T.; Iguchi-Ariga, S.M.; Ariga, H. Stimulation of transforming activity of DJ-1 by Abstrakt, a DJ-1-binding protein. Int. J. Oncol. 2005, 26, 685–689. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.H.; Lee, S.H.; Chang, H.C.; Tseng, Y.L.; Lai, W.W.; Liao, C.C.; Tsay, Y.G.; Liao, P.C. Comparative Secretome Analyses Using a Hollow Fiber Culture System with Label-Free Quantitative Proteomics Indicates the Influence of PARK7 on Cell Proliferation and Migration/Invasion in Lung Adenocarcinoma. J. Proteome. Res. 2012, 11, 5167–5185. [Google Scholar] [CrossRef]
- Cholez, E.; Debuysscher, V.; Bourgeais, J.; Boudot, C.; Leprince, J.; Tron, F.; Brassart, B.; Regnier, A.; Bissac, E.; Pecnard, E.; et al. Evidence for a protective role of the STAT5 transcription factor against oxidative stress in human leukemic pre-B cells. Leukemia 2012, 26, 2390–2397. [Google Scholar] [CrossRef] [Green Version]
- Tanti, G.K.; Goswami, S.K. SG2NA recruits DJ-1 and Akt into the mitochondria and membrane to protect cells from oxidative damage. Free Radic. Bio. Med. 2014, 75, 1–13. [Google Scholar] [CrossRef]
- Tanti, G.K.; Pandey, S.; Goswami, S.K. SG2NA enhances cancer cell survival by stabilizing DJ-1 and thus activating Akt. Biochem. Biophis. Res. Co. 2015, 463, 524–531. [Google Scholar] [CrossRef]
- Vasseur, S.; Afzal, S.; Tomasini, R.; Guillaumond, F.; Tardivel-Lacombe, J.; Mak, T.W.; Iovanna, J.L. Consequences of DJ-1 upregulation following p53 loss and cell transformation. Oncogene 2012, 31, 664–670. [Google Scholar] [CrossRef] [Green Version]
- Rahman-Roblick, R.; Hellman, U.; Becker, S.; Bader, F.G.; Auer, G.; Wiman, K.G.; Roblick, U.J. Proteomic identification of p53-dependent protein phosphorylation. Oncogene 2008, 27, 4854–4859. [Google Scholar] [CrossRef] [Green Version]
- Butler, G.S.; Dean, R.A.; Tam, E.M.; Overall, C.M. Pharmacoproteomics of a metalloproteinase hydroxamate inhibitor in breast cancer cells: Dynamics of membrane type 1 matrix metalloproteinase-mediated membrane protein shedding. Mol. Cell Biol. 2008, 28, 4896–4914. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.M.; Li, Z.R.; Huang, Y.; Yu, H.H.; Huang, X.S.; Yan, Y.F.; Shao, J.H.; Chen, H.P. DJ-1 is involved in the peritoneal metastasis of gastric cancer through activation of the Akt signaling pathway. Oncol. Rep. 2014, 31, 1489–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.P.; Fan, S.J.; An, Y.; Wang, X.; Pan, Y.; Xiaokaiti, Y.L.X.T.; Duan, J.H.; Li, X.; Tie, L.; Ye, M.; et al. Bisdemethoxycurcumin exerts pro-apoptotic effects in human pancreatic adenocarcinoma cells through mitochondrial dysfunction and a GRP78-dependent pathway. Oncotarget 2016, 7, 83641–83656. [Google Scholar] [CrossRef] [PubMed]
- Bindukumar, B.; Schwartz, S.; Aalinkeel, R.; Mahajan, S.; Lieberman, A.; Chadha, K. Proteomic profiling of the effect of prostate-specific antigen on prostate cancer cells. Prostate 2008, 68, 1531–1545. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.L.; Wang, H.B.; Yong, J.K.; Zhong, J.; Li, Q.H. MiR-128-3p overexpression sensitizes hepatocellular carcinoma cells to sorafenib induced apoptosis through regulating DJ-1. Eur. Rev. Med. Pharm. 2018, 22, 6667–6677. [Google Scholar]
- Lu, Y.B.; Ji, N.D.; Wei, W.; Sun, W.J.; Gong, X.J.; Wang, X.T. MiR-142 modulates human pancreatic cancer proliferation and invasion by targeting hypoxia-inducible factor 1 (HIF-1 alpha) in the tumor microenvironments. Biol. Open 2017, 6, 252–259. [Google Scholar] [CrossRef] [Green Version]
- Han, G.Y.; Cui, J.H.; Liang, S.; Li, H.L. Increased miR-142 and decreased DJ-1 enhance the sensitivity of pancreatic cancer cell to adriamycin. Eur. Rev. Med. Pharm. 2018, 22, 7696–7703. [Google Scholar]
- Du, S.L.; Xu, L.Y.; Gao, P.; Liu, Q.S.; Lu, F.F.; Mo, Z.H.; Fan, Z.Z.; Cheng, X.L.; Dong, Z.H. MiR-203 regulates DJ-1 expression and affects proliferation, apoptosis and DDP resistance of pancreatic cancer cells. Eur. Rev. Med. Pharm. 2019, 23, 8833–8840. [Google Scholar] [CrossRef]
- Ling, H.; He, J.; Tan, H.; Yi, L.; Liu, F.; Ji, X.X.; Wu, Y.H.; Hu, H.B.; Zeng, X.; Ai, X.H.; et al. Identification of potential targets for differentiation in human leukemia cells induced by diallyl disulfide. Int. J. Oncol. 2017, 50, 697–707. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Yang, Y.N.; Yi, L.; Qing, J.; Li, Q.Y.; Wang, W.S.; Wang, J.; Tang, Y.X.; Tan, H. Diallyl disulfide effect on the invasion and migration ability of HL-60 cells with a high expression of DJ-1 in the nucleus through the suppression of the Src signaling pathway. Oncol. Lett. 2018, 15, 6377–6385. [Google Scholar] [CrossRef]
- Saidu, N.E.B.; Kavian, N.; Leroy, K.; Jacob, C.; Nicco, C.; Batteux, F.; Alexandre, J. Dimethyl fumarate, a two-edged drug: Current status and future directions. Med. Res. Rev. 2019, 39, 1923–1952. [Google Scholar] [CrossRef]
- Saidu, N.E.B.; Noe, G.; Cerles, O.; Cabel, L.; Kavian-Tessler, N.; Chouzenoux, S.; Bahuaud, M.; Chereau, C.; Nicco, C.; Leroy, K.; et al. Dimethyl Fumarate Controls the NRF2/DJ-1 Axis in Cancer Cells: Therapeutic Applications. Mol. Cancer 2017, 16, 529–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett Saidu, N.E.; Bretagne, M.; Mansuet, A.L.; Just, P.A.; Leroy, K.; Cerles, O.; Chouzenoux, S.; Nicco, C.; Damotte, D.; Alifano, M.; et al. Dimethyl fumarate is highly cytotoxic in KRAS mutated cancer cells but spares non-tumorigenic cells. Oncotarget 2018, 9, 9088–9099. [Google Scholar] [CrossRef] [PubMed]
- Ismail, I.A.; El-Sokkary, G.H.; Saber, S.H. Low doses of Paclitaxel repress breast cancer invasion through DJ-1/KLF17 signalling pathway. Clin. Exp. Pharm. Physiol. 2018, 45, 961–968. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, L.; Wang, M.; Zhou, L.; Feng, X.P.; Yu, L.L.; Lan, J.; Gao, W.; Zhang, C.D.; Bu, Y.Q.; et al. CPX Targeting DJ-1 Triggers ROS-induced Cell Death and Protective Autophagy in Colorectal Cancer. Theranostics. 2019, 9, 5577–5594. [Google Scholar] [CrossRef]
- Kawabe, S.; Nishikawa, T.; Munshi, A.; Roth, J.A.; Chada, S.; Meyn, R.E. Adenovirus-mediated mda-7 gene expression radiosensitizes non-small cell lung cancer cells via TP53-independent mechanisms. Mol. Ther. 2002, 6, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Wang, Z.; Cui, T.; Yi, F.; Bu, Y.; Ding, S.; Ma, Y.; Song, F. Proteomic analysis of cervical cancer cells treated with adenovirus-mediated MDA-7. Cancer Biol. 2008, 7, 510–516. [Google Scholar] [CrossRef] [Green Version]
- Tashiro, S.; Caaveiro, J.M.M.; Nakakido, M.; Tanabe, A.; Nagatoishi, S.; Tamura, Y.; Matsuda, N.; Liu, D.; Hoang, Q.Q.; Tsumoto, K. Discovery and Optimization of Inhibitors of the Parkinson’s Disease Associated Protein DJ-1. ACS Chem. Biol. 2018, 13, 2783–2793. [Google Scholar] [CrossRef]
- Ogino, S.; Nowak, J.A.; Hamada, T.; Milner, D.A.; Nishihara, R. Insights into Pathogenic Interactions Among Environment, Host, and Tumor at the Crossroads of Molecular Pathology and Epidemiology. Annu. Rev. Pathol. Mech. 2019, 14, 83–103. [Google Scholar] [CrossRef]
- Ogino, S.; Chan, A.T.; Fuchs, C.S.; Giovannucci, E. Molecular pathological epidemiology of colorectal neoplasia: An emerging transdisciplinary and interdisciplinary field. Gut 2011, 60, 397–411. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Type of Cancer | Frequency | Refs |
|---|---|---|
| Astrocytoma | 103/111 (92.8%) | [26] |
| Glioma | 34/40 (85%) | [27] |
| Medulloblastomas | 32/66 (48.5%) | [28] |
| Breast cancer | 22/28 (79%) | [29] |
| Non-small cell lung carcinoma | 6/7 (86%) 13/18 (72.2%) | [30,31] |
| Thyroid cancer | 70/74 (94.6%) | [32] |
| Prostate cancer | 66/76 (86%) | [33] |
| Pancreatic neuroendocrine tumors | 21/40 (52.5%) | [34] |
| Hepatocellular carcinoma Hepatitis C virus-infected hepatocellular carcinoma (HCC) | 32/46 (69.6%) 30/32 (93.75%) | [35] [36] |
| Ovarian cancer | 63/72 (87%) | [37] |
| Esophageal squamous cell carcinoma | 11/21 (46%) | [38] |
| Cholangiocarcinoma | 5/6 (83.3%) | [39] |
| Laryngeal squamous cell cancer | 51/60 (85%) | [40] |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, W. Novel Insights into PARK7 (DJ-1), a Potential Anti-Cancer Therapeutic Target, and Implications for Cancer Progression. J. Clin. Med. 2020, 9, 1256. https://doi.org/10.3390/jcm9051256
Jin W. Novel Insights into PARK7 (DJ-1), a Potential Anti-Cancer Therapeutic Target, and Implications for Cancer Progression. Journal of Clinical Medicine. 2020; 9(5):1256. https://doi.org/10.3390/jcm9051256
Chicago/Turabian StyleJin, Wook. 2020. "Novel Insights into PARK7 (DJ-1), a Potential Anti-Cancer Therapeutic Target, and Implications for Cancer Progression" Journal of Clinical Medicine 9, no. 5: 1256. https://doi.org/10.3390/jcm9051256
APA StyleJin, W. (2020). Novel Insights into PARK7 (DJ-1), a Potential Anti-Cancer Therapeutic Target, and Implications for Cancer Progression. Journal of Clinical Medicine, 9(5), 1256. https://doi.org/10.3390/jcm9051256