Glyphosate and Saflufenacil: Elucidating Their Combined Action on the Control of Glyphosate-Resistant Conyza bonariensis

 ,
, 
Abstract
:1. Introduction
2. Material and Methods
2.1. Biotype Selection, Plant Growth, and Herbicide Treatments
- (1)
- Saflufenacil (Heat® 700 g active ingredient (a.i.) kg−1; BASF, São Paulo, SP, Brazil): 0; 3.75; 7.5; 15; 30; 60; 120 g ha−1;
- (2)
- Glyphosate (Roundup Original® DI 370 g acid equivalent (ae) L−1; Monsanto, São Paulo, SP, Brazil): 0; 185; 370; 740; 1480; 2960; 5920; 11,840; and 23,680 g ae ha−1;
- (3)
- Glyphosate and saflufenacil: glyphosate 1480 g ae ha−1 plus one of the following saflufenacil rates: 3.75, 7.5, 15, 30, 60, or 120 g ha−1 per treatment. A treatment control without herbicide was added.
2.2. Time-Course Plant Metabolism Bioassays
- (1)
- Saflufenacil: 2 × 7 × 9;
- (2)
- Glyphosate: 2 × 9 × 9;
- (3)
- Glyphosate + saflufenacil: 2 × 7 × 9.
2.3. Whole-Plant Shikimic Acid Bioassay
2.4. Oxidative Stress and Lipid Peroxidation
2.5. Statistical Analysis of the Metabolism Experiment
3. Results and Discussion
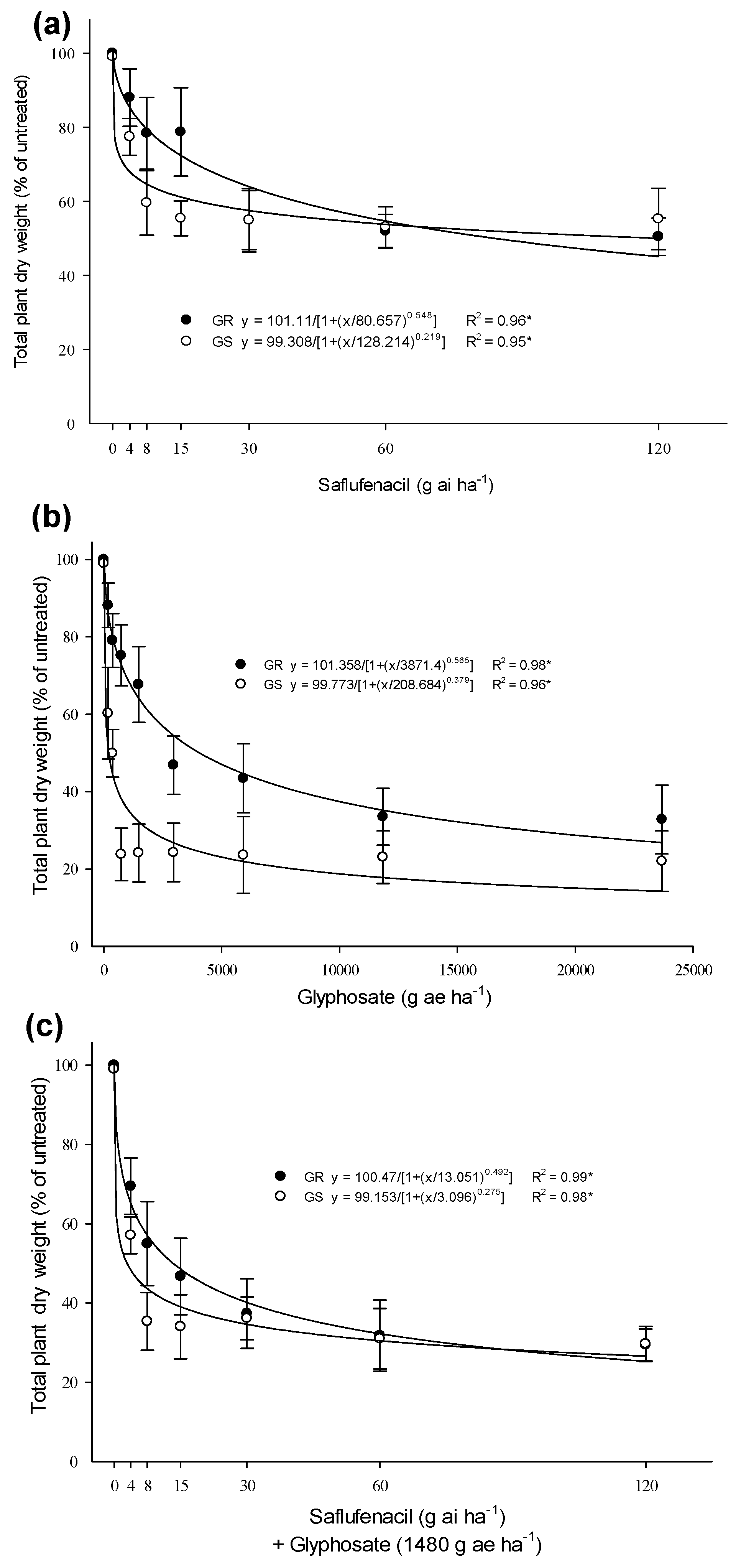
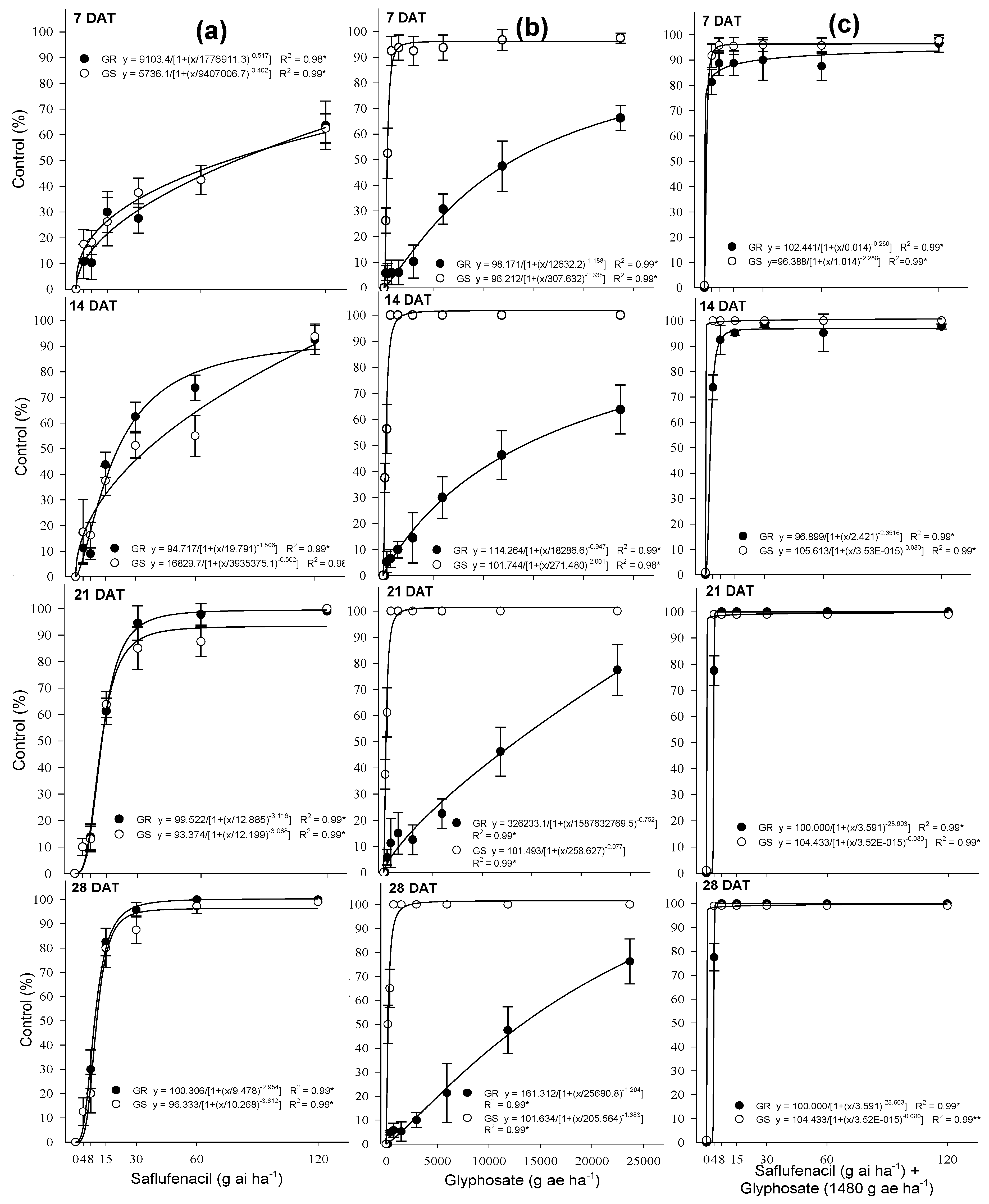
3.1. Herbicides Dose—Response and Control
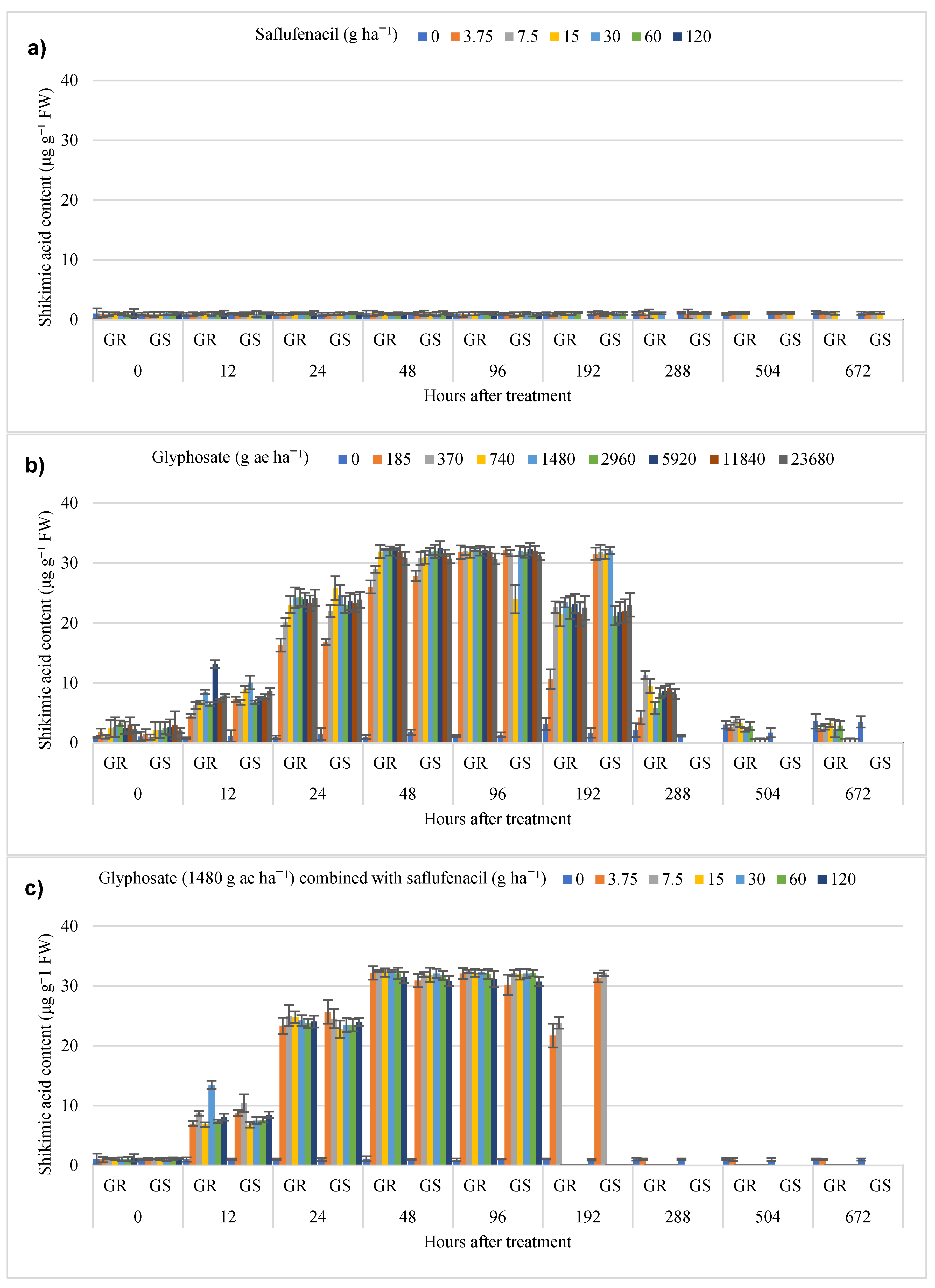
3.2. Whole-Plant Shikimic Acid Bioassay
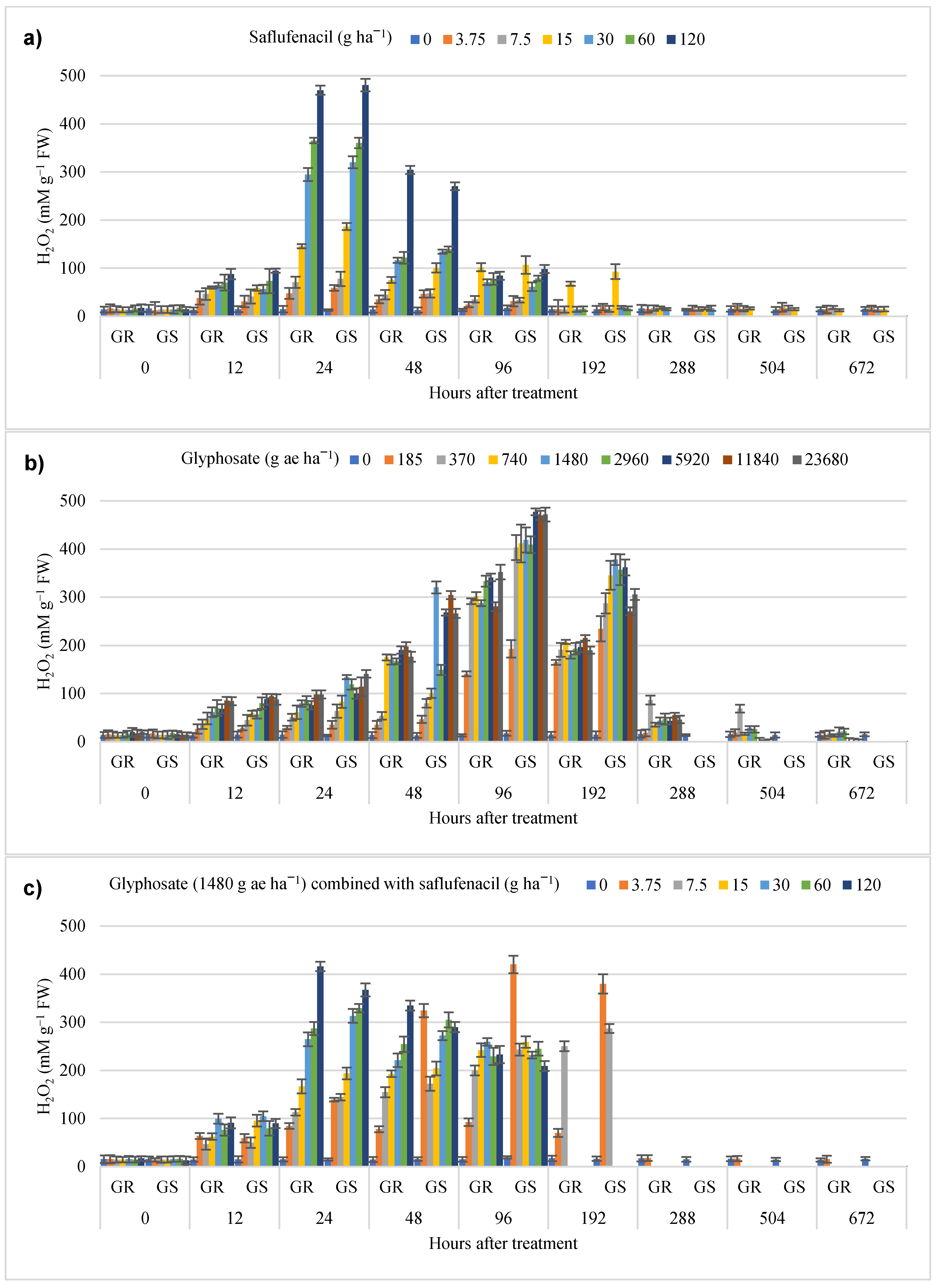
3.3. Oxidative Stress and Lipid Peroxidation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oerk, E.-C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Jeschke, P. Progress of modern agricultural chemistry and future prospects. Pest. Manag. Sci. 2016, 72, 433–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heap, I.; Duke, S.O. Overview of glyphosate-resistant weeds worldwide. Pest. Manag. Sci. 2018, 74, 1040–1049. [Google Scholar] [CrossRef] [PubMed]
- Powles, S.B.; Yu, Q. Evolution in Action: Plants Resistant to Herbicides. Annu. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef] [Green Version]
- Stewart, C.N.; Tranel, P.J.; Horvath, D.P.; Anderson, J.V.; Rieseberg, L.H.; Westwood, J.H.; Mallory-Smith, C.A.; Zapiola, M.L.; Dlugosch, K.M. Evolution of Weediness and Invasiveness: Charting the Course for Weed Genomics. Weed Sci. 2009, 57, 451–462. [Google Scholar] [CrossRef] [Green Version]
- Busi, R.; Goggin, D.E.; Heap, I.M.; Horak, M.J.; Jugulam, M.; Masters, R.A.; Napier, R.M.; Riar, D.S.; Satchivi, N.M.; Torra, J.; et al. Weed resistance to synthetic auxin herbicides. Pest. Manag. Sci. 2018, 74, 2265–2276. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, J.; Gianessi, L. Economic impacts of glyphosate-resistant weeds. In Glyphosate Resistance in Crops and Weeds: History, Development, and Management; Nandula, V.K., Ed.; Wiley: Hoboken, NJ, USA, 2010; p. 344. ISBN 978-0-470-41031-8. [Google Scholar]
- Adegas, F.; Vargas, L.; Gazziero, D.; Karam, D.; Silva, A.; Agostinetto, D. Impacto econômico da resistência de plantas daninhas a herbicidas no Brasil. Circ. Técnica 2017, 132, 1–12. [Google Scholar]
- Heap, I. The International Survey of Herbicide Resistant Weeds. Available online: www.weedscience.com (accessed on 1 May 2019).
- Bajwa, A.A.; Sadia, S.; Ali, H.H.; Jabran, K.; Peerzada, A.M.; Chauhan, B.S. Biology and management of two important Conyza weeds: A global review. Environ. Sci. Pollut. Res. 2016, 23, 24694–24710. [Google Scholar] [CrossRef] [PubMed]
- Van Gessel, M. Glyphosate-resistant horseweed from Delaware. Weed Sci. 2001, 49, 703–705. [Google Scholar] [CrossRef]
- Vargas, L.; Bianchi, M.; Rizzardi, M.; Agostinetto, D.; Dal Magro, T. Conyza bonariensis Biotypes Resistant to the Glyphosate in Southern Brazil. Planta Daninha 2007, 25, 573–578. [Google Scholar] [CrossRef]
- Silva, D.R.O.; Agostinetto, D.; Vargas, L. Molecular characterization of hairy fleabane using RAPD. Planta Daninha 2016, 34, 433–442. [Google Scholar] [CrossRef] [Green Version]
- Vargas, L.; Adegas, F.; Gazziero, D.; Karam, D.; Agostinetto, D.; da Silva, W.T. Resistência de plantas daninhas a herbicidas no Brasil: Histórico, distribuição, impacto econômico, manejo e prevenção. In A era Glyphosate: Agricultura, Meio Ambiente e Homem; Midiograf: Londrina, Brazil, 2016; pp. 219–239. [Google Scholar]
- Piasecki, C.; Yang, Y.; Benemann, D.P.; Kremer, F.S.; Galli, V.; Millwood, R.J.; Cechin, J.; Agostinetto, D.; Maia, L.C.; Vargas, L.; et al. Transcriptomic Analysis Identifies New Non-Target Site Glyphosate-Resistance Genes in Conyza bonariensis. Plants 2019, 8, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piasecki, C.; Carvalho, I.R.; Cechin, J.; Goulart, F.A.P.; da Maia, L.C.; Agostinetto, D.; Caverzan, A.; Stewart, C.N., Jr.; Vargas, L. Oxidative stress and differential antioxidant enzyme activity in glyphosate-resistant and -sensitive hairy fleabane in response to glyphosate treatment. Bragantia 2019, 78, 379–396. [Google Scholar] [CrossRef] [Green Version]
- Dalazen, G.; Kruse, N.D.; de Oliveira Machado, S.L.; Balbinot, A. Sinergismo na combinação de glifosato e saflufenacil para o controle de buva. Pesq. Agropec. Trop. 2015, 45, 249–256. [Google Scholar] [CrossRef] [Green Version]
- Knezevic, S.Z.; Datta, A.; Scott, J.; Charvat, L.D. Interactions Between Saflufenacil and Glyphosate on Selected Broadleaf Weeds. Crop. Manag. 2009, 8, 1–15. [Google Scholar] [CrossRef]
- Mellendorf, T.G.; Young, J.M.; Matthews, J.L.; Young, B.G. Influence of Plant Height and Glyphosate on Saflufenacil Efficacy on Glyphosate-Resistant Horseweed (Conyza canadensis). Weed Technol. 2013, 27, 463–467. [Google Scholar] [CrossRef]
- Piasecki, C.; Bilibio, M.I.; Cechin, J.; Schmitz, M.F.; Henckes, J.R.; Costa, L.O. da Technical effectiveness and economic return of the glyphosate-resistant hairy fleabane management in soybean pre-emergence. Rev. Bras. Herbic. 2017, 16, 20. [Google Scholar]
- Grossmann, K.; Niggeweg, R.; Christiansen, N.; Looser, R.; Ehrhardt, T. The Herbicide Saflufenacil (KixorTM) is a New Inhibitor of Protoporphyrinogen IX Oxidase Activity. Weed Sci. 2010, 58, 1–9. [Google Scholar] [CrossRef]
- Beale, S.I.; Weinstein, J.D. Tetrapyrrole metabolism in photosynthetic organisms. In Biosynthesis of Heme and Chlorophylls; Dailey, H.A., Ed.; McGraw-Hill: New York, NY, USA, 1990; pp. 287–391. [Google Scholar]
- Cobb, A.; Reade, P. Herbicides and Plant Physiology, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2010; ISBN 9781405129350. [Google Scholar]
- Caverzan, A.; Piasecki, C.; Chavarria, G.; Stewart, C.; Vargas, L. Defenses Against ROS in Crops and Weeds: The Effects of Interference and Herbicides. Int. J. Mol. Sci. 2019, 20, 1086. [Google Scholar] [CrossRef] [Green Version]
- Tzin, V.; Galili, G. New Insights into the Shikimate and Aromatic Amino Acids Biosynthesis Pathways in Plants. Mol. Plant. 2010, 3, 956–972. [Google Scholar] [CrossRef]
- Schönbrunn, E.; Eschenburg, S.; Shuttleworth, W.A.; Schloss, J.V.; Amrhein, N.; Evans, J.N.; Kabsch, W. Interaction of the herbicide glyphosate with its target enzyme 5-enolpyruvylshikimate 3-phosphate synthase in atomic detail. Proc. Natl. Acad. Sci. USA 2001, 98, 1376–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maroli, A.S.; Nandula, V.K.; Dayan, F.E.; Duke, S.O.; Gerard, P.; Tharayil, N. Metabolic Profiling and Enzyme Analyses Indicate a Potential Role of Antioxidant Systems in Complementing Glyphosate Resistance in an Amaranthus palmeri Biotype. J. Agric. Food Chem. 2015, 63, 9199–9209. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.K.; Shaner, D.L. Rapid Determination of Glyphosate Injury to Plants and Identification of Glyphosate resistant plants. Weed Sci. Soc. Am. 1998, 12, 527–530. [Google Scholar] [CrossRef]
- Perez-Jones, A.; Park, K.-W.; Polge, N.; Colquhoun, J.; Malory-Smith, C.A. Investigating the mechanisms of glyphosate resistance in Lolium multiflorum. Planta 2007, 226, 395–404. [Google Scholar] [CrossRef]
- Sergiev, I.; Alexieva, V.; Karanov, E. Effect of spermine, atrazine and combination between them on some endogenous protective systems and stress markers in plants. C. R. Acad. Bulg. Sci. 1997, 51, 121–124. [Google Scholar]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Piasecki, C.; (Federal University of Pelotas-UFPel, Pelotas, RS, Brazil). Unpublished work. 2020.
- Van Horn, C.R.; Moretti, M.L.; Robertson, R.R.; Segobye, K.; Weller, S.C.; Young, B.G.; Johnson, W.G.; Schulz, B.; Green, A.C.; Jeffery, T.; et al. Glyphosate resistance in Ambrosia trifida: Part 1. Novel rapid cell death response to glyphosate. Pest. Manag. Sci. 2018, 74, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- De Queiroz, A.R.S.; Delatorre, C.A.; Lucio, F.R.; Rossi, C.V.S.; Zobiole, L.H.S.; Merotto, A. Rapid necrosis: A novel plant resistance mechanism to 2,4-D. Weed Sci. 2019, 68, 6–18. [Google Scholar] [CrossRef]
- de Pinho, C.F.; Lourenço Leal, J.F.; dos Santos Souza, A.; de Oliveira, G.F.P.B.; de Oliveira, C.; Langaro, A.C.; Lopes Machado, A.F.; Christoffoleti, P.J.; Saes Zobiole, L.H. First evidence of multiple resistance of Sumatran Fleabane (Conyza sumatrensis (Retz.) E.Walker) to five- mode-of-action herbicides. Aust. J. Crop. Sci. 2019, 13, 1688–1697. [Google Scholar] [CrossRef]
- Waggoner, B.S.; Mueller, T.C.; Bond, J.A.; Steckel, L.E. Control of Glyphosate-Resistant Horseweed (Conyza canadensis) with Saflufenacil Tank Mixtures in No-Till Cotton. Weed Technol. 2011, 25, 310–315. [Google Scholar] [CrossRef]
- VanGessel, M.J.; Scott, B.A.; Johnson, Q.R.; White-Hansen, S.E. Influence of Glyphosate-Resistant Horseweed (Conyza canadensis) Growth Stage on Response to Glyphosate Applications. Weed Technol. 2009, 23, 49–53. [Google Scholar] [CrossRef]
- Matzenbacher, F.O.; Vidal, R.A.; Merotto Jr., A.; Trezzi, M.M. Environmental and physiological factors that affect the efficacy of herbicides that inhibit the enzyme protoporphyrinogen oxidase: A literature review. Planta Daninha 2014, 32, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Mueller, T.C.; Massey, J.H.; Hayes, R.M.; Main, C.L.; Stewart, C.N. Shikimate accumulates in both glyphosate-sensitive and glyphosate-resistant horseweed (Conyza canadensis L. Cronq.). J. Agric. Food Chem. 2003, 51, 680–684. [Google Scholar] [CrossRef]
- Ahsan, N.; Lee, D.G.; Lee, K.W.; Alam, I.; Lee, S.H.; Bahk, J.D.; Lee, B.H. Glyphosate-induced oxidative stress in rice leaves revealed by proteomic approach. Plant. Physiol. Biochem. 2008, 46, 1062–1070. [Google Scholar] [CrossRef] [PubMed]
- Nol, N.; Tsikou, D.; Eid, M.; Livieratos, I.C.; Giannopolitis, C.N. Shikimate leaf disc assay for early detection of glyphosate resistance in Conyza canadensis and relative transcript levels of EPSPS and ABC transporter genes. Weed Res. 2012, 52, 233–241. [Google Scholar] [CrossRef]
- Ye, B.; Muller, H.H.; Zhang, J.; Gressel, J. Constitutively Elevated Levels of Putrescine and Putrescine-Generating Enzymes Correlated with Oxidant Stress Resistance in Conyza bonariensis and Wheat. Plant. Physiol. 1997, 115, 1443–1451. [Google Scholar] [CrossRef] [Green Version]
- Harre, N.T.; Nie, H.; Jiang, Y.; Young, B.G. Differential antioxidant enzyme activity in rapid-response glyphosate-resistant Ambrosia trifida. Pest. Manag. Sci. 2018, 74, 2125–2132. [Google Scholar] [CrossRef]
- Ge, X.; D’Avignon, D.A.; Ackerman, J.J.H.; Sammons, R.D. Rapid vacuolar sequestration: The horseweed glyphosate resistance mechanism. Pest. Manag. Sci. 2010, 66, 345–348. [Google Scholar] [CrossRef] [Green Version]
- Starke, R.J.; Oliver, L.R. Interaction of glyphosate with chlorimuron, fomesafen, imazetaphyr, and sulfentrazone. Weed Sci. 1998, 46, 652–660. [Google Scholar] [CrossRef]
- Shaw, D.R.; Arnold, J.C. Weed control from herbicide combinations with glyphosate. Weed Technol. 2002, 16, 1–6. [Google Scholar] [CrossRef]
- Ashigh, J.; Hall, J.C. Bases for interactions between saflufenacil and glyphosate in plants. J. Agric. Food Chem. 2010, 58, 7335–7343. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Herbicide | GR50 a | RF b | |
|---|---|---|---|
| GR | GS | ||
| Saflufenacil | 83.8 ns | 120.2 ns | 0.7 c |
| Glyphosate | 4050 a | 206.2 b | 19.6 a |
| Saflufenacil + Glyphosate * | 13.2 a | 2.9 b | 4.6 b |
| Treatment | Rate (ai ha−1) | Biomass Reduction at 28 DAT (%) a | Control (%) b | |||
|---|---|---|---|---|---|---|
| 7 DAT c | 14 DAT | 21 DAT | 28 DAT | |||
| Saflufenacil | 3.75 | 15 b | 11 Ad | 7 Af | 2 Bd | 3 Bd |
| 7.5 | 21 b | 15 Bd | 18 Be | 16 Bc | 30 Ac | |
| 15 | 28 b | 22 Bc | 38 Bd | 61 Ab | 84 Ab | |
| 30 | 36 a | 31 Cc | 62 Bc | 93 Aa | 99 Aa | |
| 60 | 45 a | 44 Cb | 80 Bb | 99 Aa | 100 Aa | |
| 120 | 55 a | 63 Ba | 89 Aa | 99 Aa | 100 Aa | |
| Glyphosate 1480 g ae ha−1 | 1480 | 36 | 7 AB | 10 A | 10 A | 5 B |
| Glyphosate 1480 g ae ha−1 + Saflufenacil rates | 3.75 | 35 c | 83 Aa | 74 Ab | 78 Ab | 78 Ab |
| 7.5 | 43 bc | 86 Ba | 92 Aa | 100 Aa | 100 Aa | |
| 15 | 51 ab | 88 Ba | 96 Aa | 100 Aa | 100 Aa | |
| 30 | 60 a | 90 Aa | 97 Aa | 100 Aa | 100 Aa | |
| 60 | 68 a | 92 Aa | 97 Aa | 100 Aa | 100 Aa | |
| 120 | 75 a | 94 Aa | 97 Aa | 100 Aa | 100 Aa | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piasecki, C.; Carvalho, I.R.; Avila, L.A.; Agostinetto, D.; Vargas, L. Glyphosate and Saflufenacil: Elucidating Their Combined Action on the Control of Glyphosate-Resistant Conyza bonariensis. Agriculture 2020, 10, 236. https://doi.org/10.3390/agriculture10060236
Piasecki C, Carvalho IR, Avila LA, Agostinetto D, Vargas L. Glyphosate and Saflufenacil: Elucidating Their Combined Action on the Control of Glyphosate-Resistant Conyza bonariensis. Agriculture. 2020; 10(6):236. https://doi.org/10.3390/agriculture10060236
Chicago/Turabian StylePiasecki, Cristiano, Ivan R. Carvalho, Luis A. Avila, Dirceu Agostinetto, and Leandro Vargas. 2020. "Glyphosate and Saflufenacil: Elucidating Their Combined Action on the Control of Glyphosate-Resistant Conyza bonariensis" Agriculture 10, no. 6: 236. https://doi.org/10.3390/agriculture10060236
APA StylePiasecki, C., Carvalho, I. R., Avila, L. A., Agostinetto, D., & Vargas, L. (2020). Glyphosate and Saflufenacil: Elucidating Their Combined Action on the Control of Glyphosate-Resistant Conyza bonariensis. Agriculture, 10(6), 236. https://doi.org/10.3390/agriculture10060236