Light Quality Affects Growth and Physiology of Carpesium triste Maxim. Cultured In Vitro



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Sterilization Methods
2.2. Light Treatments and Growth Parameters
2.3. Determination of Contents of Photosynthetic Pigments
2.4. Chlorophyll a Fluorescence Transient Measurement
2.5. Determination of Activities of CAT, APX, POD, and SOD
2.5.1. Catalase (CAT; EC 1.11.1.6)
2.5.2. Ascorbate Peroxidase (APX; EC 1.11.1.11)
2.5.3. Peroxidase (POD; EC 1.11.1.7)
2.5.4. Superoxidase Dismutase (SOD; EC 1.15.1.1)
2.6. Preparation of Plant Extracts
2.7. Total Flavonoids Estimation
2.8. 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Free Radical Scavenging Capacity
2.9. Data Collection and Statistical Analysis
3. Results
3.1. Effect of Light Quality on Growth and Development of Plantlets
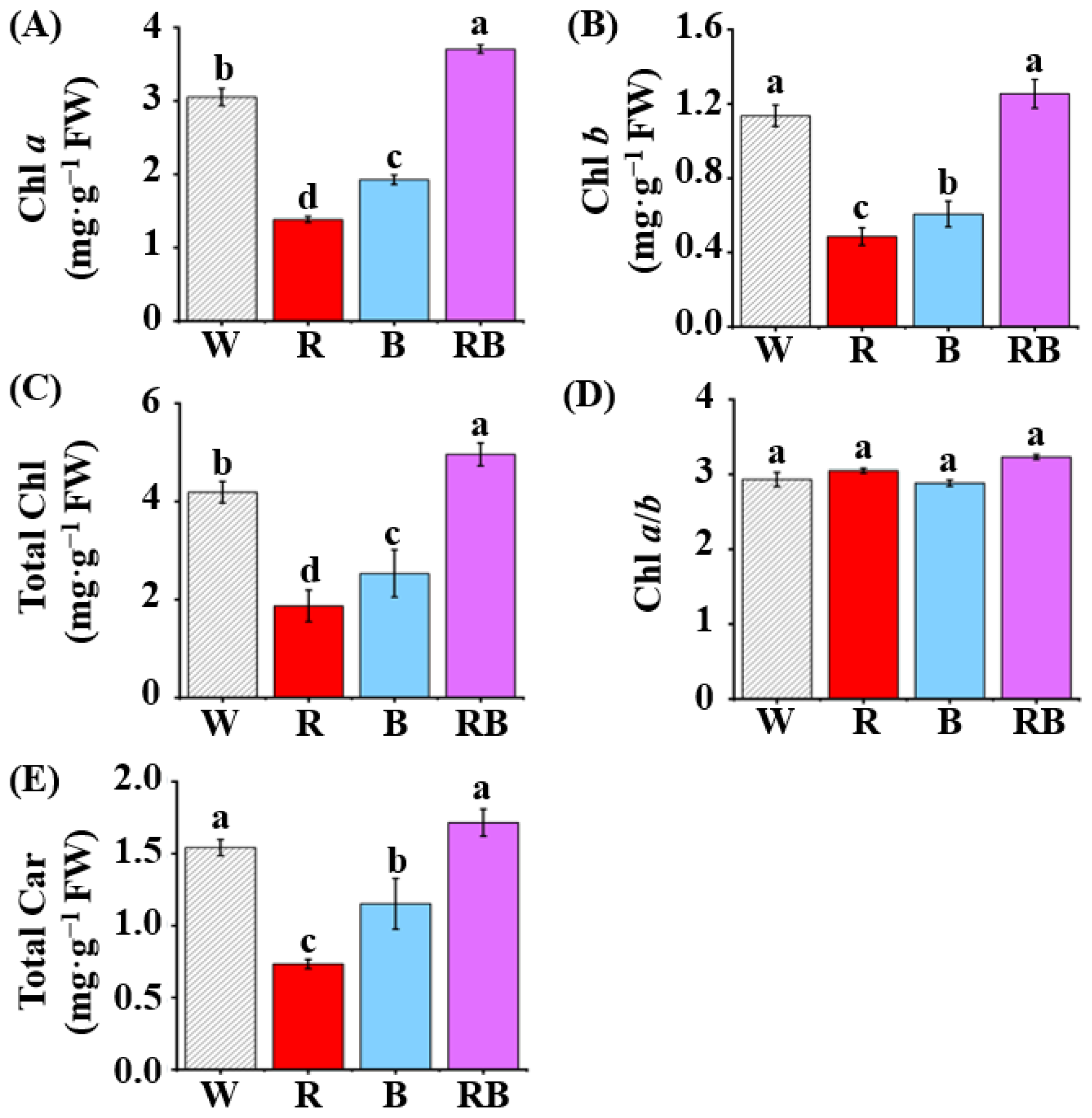
3.2. Effect of Light Quality on Contents of Photosynthetic Pigments
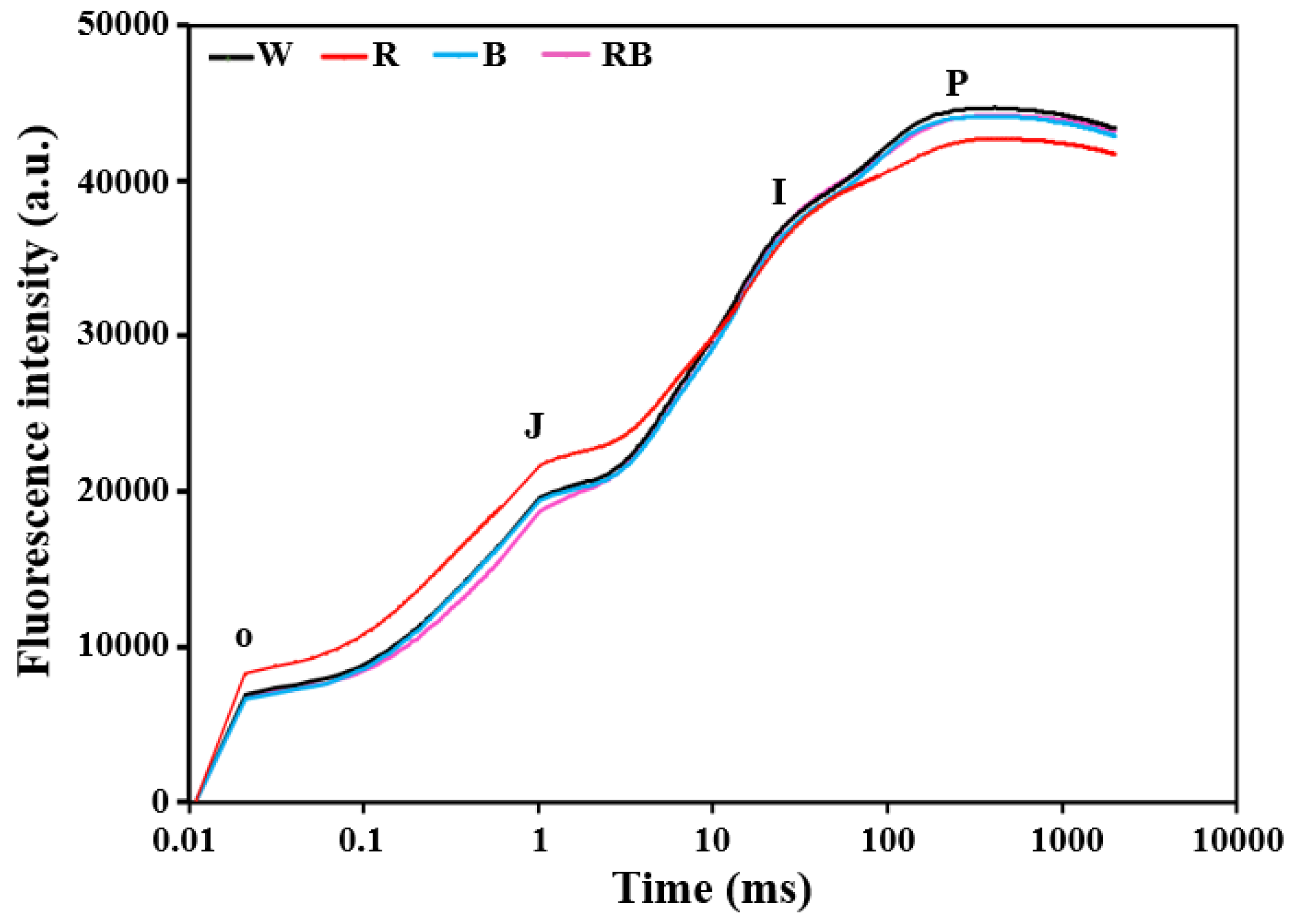
3.3. Effect of Light Quality on Parameters of Chlorophyll a Fluorescence in Leaves
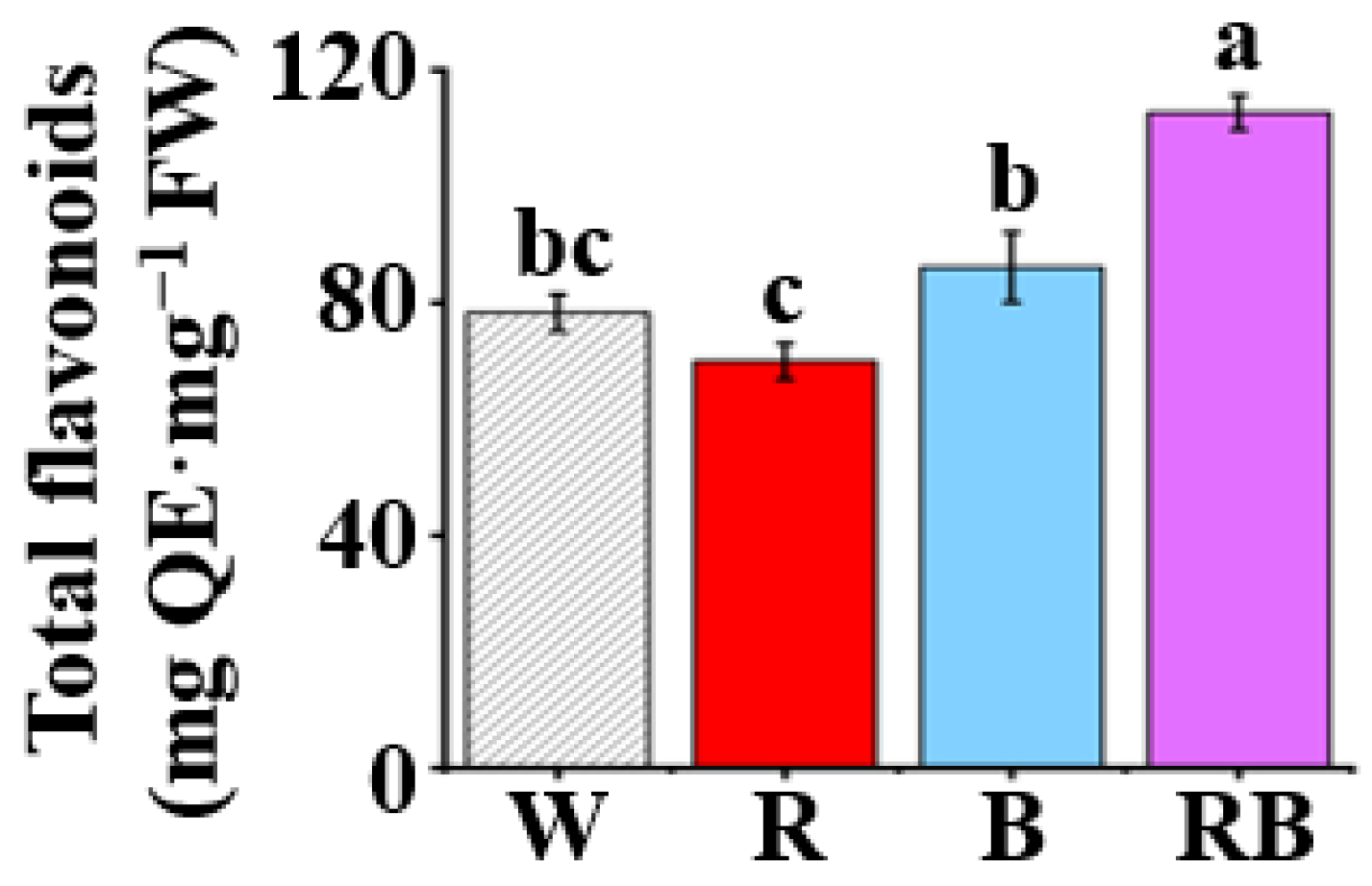
3.4. Effect of Light Quality on Content of Total Flavonoids
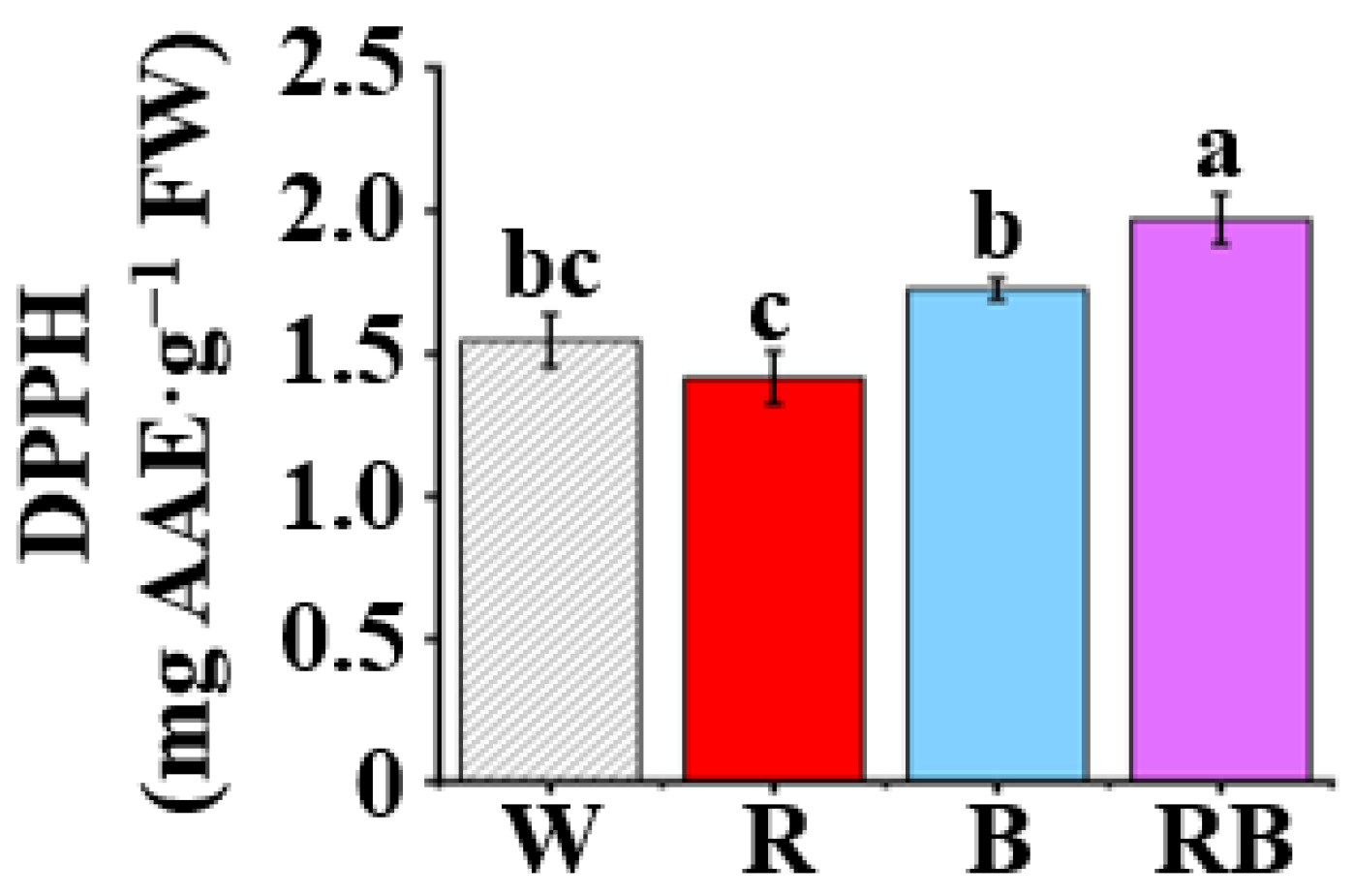
3.5. Effect of Light Quality on DPPH Radical Scavenging Capacity
3.6. Effect of Light Quality on Activities of Antioxidant Enzymes
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moon, H.I. Compounds with antiproliferative activity on five human cancer cell lines from South Korean Carpesium triste. Nat. Prod. Commun. 2012, 7, 1934578X1200700704. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.P.; Wang, G.W.; Tian, X.H.; Yang, Y.X.; Liu, Q.X.; Chen, L.P.; Li, H.L.; Zhang, W.D. The genus Carpesium: A review of its ethnopharmacology, phytochemistry and pharmacology. J. Ethnopharmacol. 2015, 163, 173–191. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Hwang, K.H.; Kim, G.H. In vitro evaluation of anti–inflammatory activity for salad–food material Carpesium Abrotanoides. J. Food Biochem. 2013, 37, 18–25. [Google Scholar] [CrossRef]
- Xia, Q.; Luo, J.; Mei, X.; Wang, Y.; Huang, W.; Wang, J.; Yang, R.; Ma, Z.; Lin, R. A developmental toxicity assay of Carpesii Fructus on zebrafish embryos/larvae. Toxicol. Res. 2017, 6, 460–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Jin, G.; Tian, L.; Ebrahim, W.; Höfert, S.P.; Janiak, C.; Chen, J.F.; Guo, Z.Y.; Schäberle, T.F.; Liu, Z.; et al. New eremophilane–type sesquiterpenes and maleimide–bearing compounds from Carpesium abrotanoides L. Fitoterapia 2019, 138, 104294. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.R.; Suh, B.R.; Kim, J.G.; Ho Kim, Y.; Kim, D.K.; Moon, D.C. Sesquiterpene lactones from Carpesium triste var. manshuricum. Phytochemistry 1999, 52, 113–115. [Google Scholar] [CrossRef]
- Kim, M.R.; Lee, S.K.; Kim, C.S.; Kim, K.S.; Moon, D.C. Phytochemical constituents of Carpesium macrocephalum FR– et SAV. Arch. Pharm. Res. 2004, 27, 1029–1033. [Google Scholar] [CrossRef]
- Kim, M.R.; Hwang, B.Y.; Jeong, E.S.; Lee, Y.M.; Yoo, H.S.; Chung, Y.B.; Hong, J.T.; Moon, D.C. Cytotoxic germacranolide sesquiterpene lactones from Carpesium triste var. manshuricum. Arch. Pharm. Res. 2007, 30, 556. [Google Scholar] [CrossRef]
- Zee, O.P.; Kim, D.K.; Choi, S.U.; Lee, C.O.; Lee, K.R. A new cytotoxic acyclic diterpene from Carpesium divaricatum. Arch. Pharm. Res. 1999, 22, 225–227. [Google Scholar] [CrossRef]
- Gao, X.; Lin, C.J.; Jia, Z.J. Cytotoxic germacranolides and acyclic diterpenoides from the seeds of Carpesium triste. J. Nat. Prod. 2007, 70, 830–834. [Google Scholar] [CrossRef]
- Zee, O.P.; Kim, D.K.; Lee, K.R. Thymol derivatives from Carpesium divaricatum. Arch. Pharm. Res. 1998, 21, 618–620. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Chen, G. Study on terpenoid constituents from aerial part of Carpesium minum Hemsl. Chem. Res. Appl. 2012, 24, 1762–1765. [Google Scholar]
- Wu, H.B.; Wu, H.B.; Wang, W.S.; Liu, T.T.; Qi, M.G.; Feng, J.C.; Li, X.Y.; Liu, Y. Insecticidal activity of sesquiterpene lactones and monoterpenoid from the fruits of Carpesium abrotanoides. Ind. Crop. Prod. 2016, 92, 77–83. [Google Scholar] [CrossRef]
- Kami, C.; Lorrain, S.; Hornitschek, P.; Fankhauser, C. Light–regulated plant growth and development. Curr. Top. Dev. Biol. 2010, 91, 29–66. [Google Scholar]
- De Wit, M.; Galvão, V.C.; Fankhauser, C. Light–mediated hormonal regulation of plant growth and development. Annu. Rev. Plant Biol. 2016, 67, 513–537. [Google Scholar] [CrossRef]
- De Capite, L. Action of light and temperature on growth of plant tissue cultures in vitro. Am. J. Bot. 1955, 42, 869–873. [Google Scholar] [CrossRef]
- Eckardt, N.A. From darkness into light: Factors controlling photomorphogenesis. Plant Cell 2001, 13, 219–221. [Google Scholar] [CrossRef]
- Hu, W.; Lagarias, J.C. A tightly regulated genetic selection system with signaling–active alleles of phytochrome B. Plant Physiol. 2017, 173, 366–375. [Google Scholar] [CrossRef] [Green Version]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; Ieperen, W.V.; Harbinson, J. Blue light dose–responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef]
- Sun, W.; Ubierna, N.; Ma, J.Y.; Walker, B.J.; Kramer, D.M.; Cousins, A.B. The coordination of C4 photosynthesis and the CO2–concentrating mechanism in maize and Miscanthus × giganteus in response to transient changes in light quality. Plant Physiol. 2014, 164, 1283–1292. [Google Scholar] [CrossRef] [Green Version]
- Bantis, F.; Ouzounis, T.; Radoglou, K. Artificial LED lighting enhances growth characteristics and total phenolic content of Ocimum basilicum, but variably affects transplant success. Sci. Hortic. 2016, 198, 277–283. [Google Scholar] [CrossRef] [Green Version]
- Baque, M.A.; Hahn, E.J.; Paek, K.Y. Induction mechanism of adventitious root from leaf explants of Morinda citrifolia as affected by auxin and light quality. In Vitro Cell. Dev. Biol. Plant 2010, 46, 71–80. [Google Scholar] [CrossRef]
- Lin, Y.; Li, J.; Li, B.; He, T.; Chun, Z. Effects of light quality on growth and development of protocorm–like bodies of Dendrobium officinale in vitro. Plant Cell Tiss. Organ Cult. 2011, 105, 329–335. [Google Scholar] [CrossRef]
- Fankhauser, C.; Batschauer, A. Shadow on the plant: A strategy to exit. Cell 2016, 164, 15–17. [Google Scholar] [CrossRef] [Green Version]
- Franklin, K.A.; Whitelam, G.C. Light-quality regulation of freezing tolerance in Arabidopsis thaliana. Nat. Genet. 2007, 39, 1410–1413. [Google Scholar] [CrossRef]
- Li, Q.; Kubota, C. Effects of supplemental light quality on growth and phytochemicals of baby leaf lettuce. Environ. Exp. Bot. 2009, 67, 59–64. [Google Scholar] [CrossRef]
- Gubler, F.; Hughes, T.; Waterhouse, P.; Jacobsen, J. Regulation of dormancy in barley by blue light and after-ripening: Effects on abscisic acid and gibberellin metabolism. Plant Physiol. 2008, 147, 886–896. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.H.; Wheeler, R.M.; Sager, J.C.; Yorio, N.C.; Goins, G.D. Light–emitting diodes as an illumination source for plants: A review of research at Kennedy Space Center. Habitation 2005, 10, 71–78. [Google Scholar] [CrossRef]
- Bourget, C.M. An introduction to light–emitting diodes. HortScience 2008, 43, 1944–1946. [Google Scholar] [CrossRef] [Green Version]
- Heo, J.W.; Lee, C.W.; Paek, K.Y. Influence of mixed LED radiation on the growth of annual plants. J. Plant Biol. 2006, 49, 286–290. [Google Scholar] [CrossRef]
- Pham, M.D.; Hwang, H.; Park, S.W.; Cui, M.; Lee, H.; Chun, C. Leaf chlorosis, epinasty, carbohydrate contents and growth of tomato show different responses to the red/blue wavelength ratio under continuous light. Plant Physiol. Bioch. 2019, 141, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.W.; Lee, Y.B.; Kim, D.E.; Chang, Y.S.; Chun, C. Effects of supplementary LED lighting on growth and biochemical parameters in Dieffenbachia amoena ‘Camella’and Ficus elastica ‘Melany’. Korea J. Hortic. Sci. Technol. 2010, 28, 51–58. [Google Scholar]
- Jee, G. Sixty–three years since Kautsky: Chlorophyll a fluorescence. Aust. J. Plant Physiol. 1995, 22, 131–160. [Google Scholar]
- Stirbet, A.; Lazár, D.; Kromdijk, J. Chlorophyll a fluorescence induction: Can just a one–second measurement be used to quantify abiotic stress responses? Photosynthetica 2018, 56, 86–104. [Google Scholar] [CrossRef]
- Strasser, R.J. The grouping model of plant photosynthesis: Heterogeneity of photosynthetic units in thylakoids. In Photosynthesis: Structure and Molecular Organization of the Photosynthetic Apparatus; Akoyunoglou, G., Ed.; Balaban International Science Services: Philadelphia, PA, USA, 1981; Volume III, pp. 727–737. [Google Scholar]
- Strasser, R.J. The F0 and the O–J–I–P fluorescence rise in higher plants and algae. In Regulation of Chloroplast Biogenesis; Argyroudi–Akoyunoglou, J.H., Ed.; Springer: Boston, MA, USA, 1992; pp. 423–426. [Google Scholar]
- Strasser, R. On the OJIP fluorescence transient in leaves and D1 mutants of Chlamydomonas reinhardtii. In Research in Photosynthesis: Proceedings of the IXth International Congress on Photosynthesis, Nagoya, Japan, 30 August–4 September 1992; Murata, N., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1992; Volume II, pp. 29–32. [Google Scholar]
- Strasser, R.J.; Srivastava, A. Polyphasic chlorophyll a fluorescent transient in plants and cyanobacteria. Photochem. Photobiol. 1995, 61, 32–42. [Google Scholar] [CrossRef]
- Stirbet, A. On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and photosystem II: Basics and applications of the OJIP fluorescence transient. J. Photochem. Photobiol. B 2011, 104, 236–257. [Google Scholar] [CrossRef]
- Schreiber, U.; Neubauer, C.; Klughammer, C. Devices and methods for room–temperature fluorescence analysis. Philos. Trans. R. Soc. B 1989, 323, 241–251. [Google Scholar]
- Schansker, G.; Strasser, R.J. Quantification of non–QB–reducing centers in leaves using a far–red pre–illumination. Photosynth. Res. 2005, 84, 145–151. [Google Scholar] [CrossRef]
- Schansker, G.; Tóth, S.Z.; Holzwarth, A.R.; Garab, G. Chlorophyll a fluorescence: Beyond the limits of the QA model. Photosynth. Res. 2014, 120, 43–58. [Google Scholar] [CrossRef]
- Roháček, K. Chlorophyll fluorescence parameters: The definitions, photosynthetic meaning, and mutual relationships. Photosynthetica 2002, 40, 13–29. [Google Scholar] [CrossRef]
- Waszczak, C.; Carmody, M.; Kangasjärvi, J. Reactive oxygen species in plant signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oukarroum, A.; Bussotti, F.; Goltsev, V.; Kalaji, H.M. Correlation between reactive oxygen species production and photochemistry of photosystems I and II in Lemna gibba L. plants under salt stress. Environ. Exp. Bot. 2015, 109, 80–88. [Google Scholar] [CrossRef]
- Triantaphylidès, C.; Havaux, M. Singlet oxygen in plants: Production, detoxification and signaling. Trends Plant Sci. 2009, 14, 219–228. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Eghbaliferiz, S.; Iranshahi, M. Prooxidant activity of polyphenols, flavonoids, anthocyanins and carotenoids: Updated review of mechanisms and catalyzing metals. Phytother. Res. 2016, 30, 1379–1391. [Google Scholar] [CrossRef]
- Baglieri, A.; Cadili, V.; Monterumici, C.M.; Gennari, M.; Tabasso, S.; Montoneri, E.; Nardi, S.; Negre, M. Fertilization of bean plants with tomato plants hydrolysates. Effect on biomass production, chlorophyll content and N assimilation. Sci. Hortic. 2014, 176, 194–199. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Strasser, R.J.; Srivastava, A.; Tsimilli–Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. In Probing Photosynthesis: Mechanism, Regulation and Adaptation; Yunus, M., Pathre, U., Mohanty, P., Eds.; Taylor and Francis: Abingdon, UK, 2000; Chapter 25; pp. 445–483. [Google Scholar]
- Strasser, R.J.; Tsimilli–Michael, M.; Srivastava, A. Analysis of the chlorophyll fluorescence transient. In Chlorophyll Fluorescence: A Signature of Photosynthesis, Advances in Photosynthesis and Respiration; Papageorgiou, G.C., Ed.; Springer: Dordrecht, The Netherlands, 2004; Volume 19, pp. 321–362. [Google Scholar]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Packer, L., Ed.; Academic Press: New York, NY, USA, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate–specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Pütter, J. Peroxidase. In Method of Enzymatic Method Analysis; Bergmeyer, H.U., Ed.; Academic Press: New York, NY, USA, 1974; Volume 2, pp. 685–690. [Google Scholar]
- Beauchamp, C.; Fridovich, I. Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Manivannan, A.; Ren, X.; Kang, D.I.; Jeong, B.R. Effect of explant type and plant growth regulators on callus induction, growth and secondary metabolites production in Cnidium officinale Makino. Mol. Biol. Rep. 2018, 45, 1919–1927. [Google Scholar]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar]
- Khan, T.; Abbasi, B.H.; Khan, M.A.; Shinwari, Z.K. Differential effects of thidiazuron on production of anticancer phenolic compounds in callus cultures of Fagonia indica. Appl. Biochem. Biotechnol. 2016, 179, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Tóth, S.Z.; Schansker, G.; Garab, G.; Strasser, R.J. Photosynthetic electron transport activity in heat–treated barley leaves: The role of internal alternative electron donors to photosystem II. Biochim. Biophys. Acta. 2007, 1767, 295–305. [Google Scholar] [CrossRef] [Green Version]
- Dewir, Y.H.; Chakrabarty, D.; Kim, S.J.; Hahn, E.J.; Paek, K.Y. Effect of light–emitting diode on growth and shoot proliferation of Euphorbia millii and Spathiphyllum cannifolium. J. Korea Soc. Hortic. Sci. 2005, 46, 375. [Google Scholar]
- Moon, H.K.; Park, S.K.; Kim, Y.W.; Kim, C.S. Growth of Tsuru–rindo (Tripterospermum japonicum) cultured in vitro under various sources of light–emitting diode (LED) irradiation. J. Plant Biol. 2006, 49, 174–179. [Google Scholar] [CrossRef]
- Chung, J.P.; Huang, C.Y.; Dai, T.E. Spectral effects on embryogenesis and plantlet growth of Oncidium ‘Gower Ramsey’. Sci. Hortic. 2010, 124, 511–516. [Google Scholar] [CrossRef]
- Maas, F.M.; Bakx, E.J.; Morris, D.A. Photocontrol of stem elongation and dry weight partitioning in Phaseolus vulgaris L. by the blue-light content of photosynthetic photon flux. J. Plant Physiol. 1995, 146, 665–671. [Google Scholar] [CrossRef]
- Appelgren, M. Effects of light quality on stem elongation of Pelargonium in vitro. Sci. Hortic. 1991, 45, 345–351. [Google Scholar] [CrossRef]
- You, T.; Barnett, S.M. Effect of light quality on production of extracellular polysaccharides and growth rate of Porphyridium cruentum. Biochem. Eng. J. 2004, 19, 251–258. [Google Scholar] [CrossRef]
- Bonnett, H.T. Phytochrome regulation of endogenous bud development on root cultures of Convolvulus arvensis. Planta 1972, 106, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Hahn, E.J.; Heo, J.W.; Paek, K.Y. Effects of LEDs on net photosynthetic rate, growth and leaf stomata of chrysanthemum plantlets in vitro. Sci. Hortic. 2004, 101, 143–151. [Google Scholar] [CrossRef]
- Heo, J.W.; Shin, K.S.; Kim, S.K.; Paek, K.Y. Light quality affects in vitro growth of grape ‘Teleki 5BB’. J. Plant Biol. 2006, 49, 276–280. [Google Scholar] [CrossRef]
- Nhut, D.T.; Nam, N.B. Light emitting diodes (LEDs): An artificial lighting source for biological studies. In Proceedings of the 3rd International Conference of the Development of BME in Vietnam, Ho Chi Minh, Vietnam, 11–14 January 2010. [Google Scholar]
- Goins, G.D.; Yorio, N.C.; Sanwo, M.M.; Brown, C.S. Photomorphogenesis, photosynthesis, and seed yield of wheat plants grown under red light-emitting diodes (LEDs) with and without supplemental blue lighting. J. Exp. Bot. 1997, 48, 1407–1413. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Takamura, T.; Watanabe, H.; Endo, M.; Yanagi, T.; Okamoto, K. In vitro growth of Cymbidium plantlets cultured under superbright red and blue light-emitting diodes (LEDs). J. Hortic. Sci. Biotech. 1998, 73, 39–44. [Google Scholar] [CrossRef]
- Lee, S.H.; Tewari, R.K.; Hahn, E.J.; Paek, K.Y. Photon flux density and light quality induce changes in growth, stomatal development, photosynthesis and transpiration of Withania somnifera (L.) Dunal. plantlets. Plant Cell Tiss. Organ Cult. 2007, 90, 141–151. [Google Scholar] [CrossRef]
- Bellamine, J.; Penel, C.; Greppin, H. Proton pump and IAA sensitivity changes in spinach leaves during the flowering induction. Plant Physiol. Biochem. 1993, 31, 197–203. [Google Scholar]
- Moe, R.; Morgan, L.; Grindal, G. Growth and plant morphology of Cucumis sativus and Fuchsia x hybrid are influenced by light quality during the photoperiod and by diurnal temperature alternations. Acta Hortic. 2002, 580, 229–234. [Google Scholar] [CrossRef]
- Poudel, P.R.; Kataoka, I.; Mochioka, R. Effect of red–and blue–light–emitting diodes on growth and morphogenesis of grapes. Plant Cell Tiss. Org. 2008, 92, 147–153. [Google Scholar] [CrossRef]
- Kurepin, L.V.; Emery, R.N.; Pharis, R.P.; Reid, D.M. Uncoupling light quality from light irradiance effects in Helianthus annuus shoots: Putative roles for plant hormones in leaf and internode growth. J. Exp. Bot. 2007, 58, 2145–2157. [Google Scholar] [CrossRef] [Green Version]
- Weatherwax, S.C.; Ong, M.S.; Degenhardt, J.; Bray, E.A.; Tobin, E.M. The interaction of light and abscisic acid in the regulation of plant gene expression. Plant Physiol. 1996, 111, 363–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathy, B.C.; Brown, C.S. Root–shoot interaction in the greening of wheat seedlings grown under red light. Plant Physiol. 1995, 107, 407–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venema, J.H.; Villerius, L.; van Hasselt, P.R. Effect of acclimation to suboptimal temperature on chilling-induced photodamage: Comparison between a domestic and a high-altitude wild Lycopersicon species. Plant Sci. 2000, 152, 153–163. [Google Scholar] [CrossRef]
- Leong, T.Y.; Anderson, J.M. Adaptation of the thylakoid membranes of pea chloroplasts to light intensities. I. Study on the distribution of chlorophyll–protein complexes. Photosynth. Res. 1984, 5, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Tholen, D.; Pons, T.L.; Voesenek, L.A.C.J.; Poorter, H. Ethylene insensitivity results in down–regulation of Rubisco expression and photosynthetic capacity in tobacco. Plant Physiol. 2007, 144, 1305–1315. [Google Scholar] [CrossRef] [Green Version]
- Kurilčik, A.; Miklušytė–Čanova, R.; Dapkūnienė, S.; Žilinskaitė, S.; Kurilčik, G.; Tamulaitis, G.; Duchovskis, P.; Žukauskas, A. In vitro culture of Chrysanthemum plantlets using light–emitting diodes. Cent. Eur. J. Biol. 2008, 3, 161–167. [Google Scholar] [CrossRef]
- Bukhov, N.G.; Drozdova, I.S.; Bondar, V.V.; Mokronosov, A.T. Blue, red and blue plus red light control of chlorophyll content and CO2 gas exchange in barley leaves: Quantitative description of the effects of light quality and fluence rate. Physiol. Plant. 1992, 85, 632–638. [Google Scholar] [CrossRef]
- Goodwin, T.W.; Britton, G. Distribution and analysis of carotenoids. In Plant Pigments; Goodwin, T.W., Ed.; Academic Press: London, UK, 1988; pp. 61–132. [Google Scholar]
- Haehnel, W. Photosynthetic electron transport in higher plants. Annu. Rev. Plant Physiol. 1984, 35, 659–693. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, T.; Fang, S.; Zhou, M.; Qin, J. Responses of morphology, gas exchange, photochemical activity of photosystem II, and antioxidant balance in Cyclocarya paliurus to light spectra. Front. Plant Sci. 2018, 9, 1704. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Guo, P.; Michael, B.; Stefania, G.; Salvatore, C. Evaluation of chlorophyll content and fluorescence parameters as indicators of drought tolerance in barley. Agric. Sci. China 2006, 5, 751–757. [Google Scholar] [CrossRef]
- Shangguan, Z.; Shao, M.; Dyckmans, J. Effects of nitrogen nutrition and water deficit on net photosynthetic rate and chlorophyll fluorescence in winter wheat. J. Plant Physiol. 2000, 156, 46–51. [Google Scholar] [CrossRef]
- Clark, A.J.; Landolt, W.; Bucher, J.B.; Strasser, R.J. Beech (Fagus sylvatica) response to ozone exposure assessed with a chlorophyll fluorescence performance index. Environ. Pollut. 2000, 109, 501–507. [Google Scholar] [CrossRef]
- Ke, B. Photosynthesis–photobiochemistry and photobiophysics. In Advances in Photosynthesis; Ke, B., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001; Volume 10, pp. 1–763. [Google Scholar]
- Strasser, R.J.; Tsimilli–Michael, M.; Qiang, S.; Goltsev, V. Simultaneous in vivo recording of prompt and delayed fluorescence and 820–nm reflection changes during drying and after rehydration of the resurrection plant Haberlea rhodopensis. Biochim. Biophys. Acta 2010, 1797, 1313–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estaji, A.; Kalaji, H.M.; Karimi, H.R.; Roosta, H.R.; Moosavi–Nezhad, S.M. How glycine betaine induces tolerance of cucumber plants to salinity stress? Photosynthetica 2019, 57, 753–761. [Google Scholar] [CrossRef] [Green Version]
- Yusuf, M.A.; Kumar, D.; Rajwanshi, R.; Strasser, R.J.; Tsimilli–Michael, M.; Sarin, N.B. Overexpression of γ–tocopherol methyl transferase gene in transgenic Brassica juncea plants alleviates abiotic stress: Physiological and chlorophyll a fluorescence measurement. Biochim. Biophys. Acta 2010, 1797, 1428–1438. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Zhou, X.; Xu, R.; Wang, J.; Lin, Y.; Pang, J.; Wu, S.; Zhong, F. Comprehensive analysis of photosynthetic characteristics and quality improvement of purple cabbage under different combinations of monochromatic light. Front. Plant Sci. 2016, 7, 1788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aliniaeifard, S.; Seif, M.; Arab, M.; Zare Mehrjerdi, M.; Li, T.; Lastochkina, O. Growth and photosynthetic performance of Calendula officinalis under monochromatic red light. Int. J. Hortic. Sci. Technol. 2018, 5, 123–132. [Google Scholar]
- Bayat, L.; Arab, M.; Aliniaeifard, S.; Seif, M.; Lastochkina, O.; Li, T. Effects of growth under different light spectra on the subsequent high light tolerance in rose plants. AoB Plants 2018, 10, 1–17. [Google Scholar] [CrossRef]
- Van Huylenbroeck, J.M.; Debergh, P.C. Impact of sugar concentration in vitro on photosynthesis and carbon metabolism during ex vitro acclimatization of Spathiphyllum plantlets. Physiol. Plant. 1996, 96, 298–304. [Google Scholar] [CrossRef]
- Rybczyński, J.J.; Borkowska, B.; Fiuk, A.; Gawrońska, H.; Śliwińska, E.; Mikuła, A. Effect of sucrose concentration on photosynthetic activity of in vitro cultures Gentiana kurroo (Royle) germlings. Acta Physiol. Plant. 2007, 29, 445–453. [Google Scholar] [CrossRef]
- Trojan, A.; Gabrys, H. Chloroplast distribution in Arabidopsis thaliana (L.) depends on light conditions during growth. Plant Physiol. 1996, 111, 419–425. [Google Scholar] [CrossRef] [Green Version]
- Kadleček, P.; Rank, B.; Tichá, I. Photosynthesis and photoprotection in Nicotiana tabacum L. in vitro-grown plantlets. J. Plant Physiol. 2003, 160, 1017–1024. [Google Scholar] [CrossRef] [PubMed]
- Lucchesini, M.; Monteforti, G.; Mensuali-Sodi, A.; Serra, G. Leaf ultrastructure, photosynthetic rate and growth of myrtle plantlets under different in vitro culture conditions. Biol. Plant. 2006, 50, 161–168. [Google Scholar] [CrossRef]
- Eckstein, A.; Zięba, P.; Gabryś, H. Sugar and light effects on the condition of the photosynthetic apparatus of Arabidopsis thaliana cultured in vitro. J. Plant Growth Regul. 2012, 31, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, S.; Raghuvanshi, R.; Bhardwaj, P.; Sood, H.; Saxena, S.; Chaurasia, O.P. Influence of light quality on growth, secondary metabolites production and antioxidant activity in callus culture of Rhodiola imbricata Edgew. J. Photoch. Photobio. B Biol. 2018, 183, 258–265. [Google Scholar] [CrossRef]
- Manivannan, A.; Soundararajan, P.; Halimah, N.; Ko, C.H.; Jeong, B.R. Blue LED light enhances growth, phytochemical contents, and antioxidant enzyme activities of Rehmannia glutinosa cultured in vitro. Hortic. Environ. Biotechnol. 2015, 56, 105–113. [Google Scholar] [CrossRef]
- Manivannan, A.; Ren, X.; Jeong, B.R. Light elicited growth, antioxidant enzymes activities and production of medicinal compounds in callus culture of Cnidium officinale Makino. J. Photoch. Photobio. B Biol. 2019, 196, 111509. [Google Scholar]
- Hoque, M.A.; Banu, M.N.A.; Okuma, E.; Amako, K.; Nakamura, Y.; Shimoishi, Y.; Murata, Y. Exogenous proline and glycinebetaine increase NaCl–induced ascorbate–glutathione cycle enzyme activities, and proline improves salt tolerance more than glycinebetaine in tobacco Bright Yellow–2 suspension–cultured cells. J. Plant Physiol. 2007, 164, 1457–1468. [Google Scholar] [CrossRef]
- Mittova, V.; Volokita, M.; Guy, M.; Tal, M. Activities of SOD and the ascorbate-glutathione cycle enzymes in subcellular compartments in leaves and roots of the cultivated tomato and its wild salt-tolerant relative Lycopersicon pennellii. Physiol. Plant. 2000, 110, 42–51. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defense antioxidants–superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defense grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Veitch, N.C. Structural determinants of plant peroxidase function. Phytochem. Rev. 2004, 3, 3–18. [Google Scholar] [CrossRef]
- Genkov, T.; Ivanova, I. Effect of cytokinin–active phenylurea derivatives on shoot multiplication, peroxidase and superoxide dismutase activities of in vitro cultured carnation. Bulg. J. Plant Physiol. 1995, 21, 73. [Google Scholar]
- Blazquez, S.; Olmos, E.; Hernández, J.A.; Fernández–García, N.; Fernández, J.A.; Piqueras, A. Somatic embryogenesis in saffron (Crocus sativus L.). Histological differentiation and implication of some components of the antioxidant enzymatic system. Plant Cell Tiss. Organ Cult. 2009, 97, 49–57. [Google Scholar] [CrossRef]
- Shafi, A.; Chauhan, R.; Gill, T.; Swarnkar, M.K.; Sreenivasulu, Y.; Kumar, S.; Kumar, N.; Shankar, R.; Ahuja, P.S.; Singh, A.K. Expression of SOD and APX genes positively regulates secondary cell wall biosynthesis and promotes plant growth and yield in Arabidopsis under salt stress. Plant Mol. Biol. 2015, 87, 615–631. [Google Scholar] [CrossRef]
- Shohael, A.M.; Ali, M.B.; Yu, K.W.; Hahn, E.J.; Islam, R.; Paek, K.Y. Effect of light on oxidative stress, secondary metabolites and induction of antioxidant enzymes in Eleutherococcus senticosus somatic embryos in bioreactor. Process. Biochem. 2006, 41, 1176–1185. [Google Scholar] [CrossRef]
- Prochazkova, D.; Wilhelmova, N. Leaf senescence and activities of the antioxidant enzymes. Biol. Plant. 2007, 51, 401–406. [Google Scholar] [CrossRef]
- Pastori, G.M.; del Río, L.A. Natural senescence of pea leaves (an activated oxygen-mediated function for peroxisomes). Plant Physiol. 1997, 113, 411–418. [Google Scholar] [CrossRef] [Green Version]
- Causin, H.F.; Jauregui, R.N.; Barneix, A.J. The effect of light spectral quality on leaf senescence and oxidative stress in wheat. Plant Sci. 2006, 171, 24–33. [Google Scholar] [CrossRef]
- Chen, J.; Li, F.; Li, Y.; Wang, Y.; Wang, C.; Yuan, D.; Jiang, Y. Exogenous procyanidin treatment delays senescence of harvested banana fruit by enhancing antioxidant responses and in vivo procyanidin content. Postharvest Biol. Tec. 2019, 158, 110999. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formula Abbreviation | Explanation |
|---|---|
| F0 | Minimum fluorescence in the dark-adapted state |
| FM | Maximum fluorescence at a saturating radiation pulse in the dark-adapted state |
| FV | Maximum variable fluorescence |
| FV/FM | Maximum quantum efficiency of PS II photochemistry |
| FM/F0 | Electron transfer activity |
| FV/F0 | Actual quantum yield of PS II |
| M0 = TR0/RC − ET0/RC = 4 (F300μs − F0)/(FM − F0) | Approximated initial slope of the fluorescence transient normalized on the FV |
| VJ = (FJ − F0)/FV | Relative variable fluorescence for J state |
| SM = Area/(FM − F0) | Normalized total complementary area |
| φP0 = (1 − F0)/F | Quantum yield of the QA reduction |
| φE0 = (1 − (F0/FM))·ψ0 | Quantum yield for electron transport |
| φD0 = 1 − φP0 | The quantum yield of energy dissipation |
| Ψ0 = 1 − VJ | Possibility of a trapped exciton moves an electron into the electron transport chain beyond QA |
| ABS/RC = M0·(1/VJ)·(FM/FV) | Absorption flux of antenna chlorophyll per reaction center |
| ET0/RC = M0·(1/VJ)·φE0 | Electron transport flux per reaction center |
| TR0/RC = M0·(1/VJ) | Trapped energy flux per reaction center |
| DI0/RC = ABS/RC − TR0/RC | Heat dissipation per reaction center |
| PIABS = (RC/ABS)·(φP0/(1 − φP0))·(ψ0/(1 − ψ0)) | Performance index per absorption flux |
| Light Quality | Shoot | Root | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Number | Length (cm) | Fresh Weight (g) | Dry Weight (g) | Diameter (mm) | Number | Length (cm) | Fresh Weight (g) | Dry Weight (g) | |
| W z | 5 ± 0.00 y a x | 10.19 ± 0.43 b | 0.66 ± 0.06 c | 0.09 ± 0.01 c | 5.72 ± 0.20 b | 6 ± 1.00 b | 4.78 ± 0.52 b | 1.18 ± 0.12 a | 0.16 ± 0.02 a |
| R | 3 ± 0.58 b | 14.30 ± 0.56 a | 0.97 ± 0.08 b | 0.13 ± 0.01 b | 5.22 ± 0.09 c | 8 ± 0.58 ab | 5.77 ± 0.30 a | 0.74 ± 0.04 b | 0.10 ± 0.01 b |
| B | 4 ± 0.58 b | 5.52 ± 0.22 d | 0.43 ± 0.04 d | 0.06 ± 0.01 d | 4.38 ± 0.10 d | 3 ± 0.58 c | 2.62 ± 0.22 c | 0.25 ± 0.05 c | 0.03 ± 0.01 c |
| RB | 6 ± 0.58 a | 7.00 ± 0.49 c | 1.25 ± 0.12 a | 0.17 ± 0.02 a | 6.11 ± 0.11 a | 9 ± 0.58 a | 5.55 ± 0.23 ab | 1.24 ± 0.10 a | 0.17 ± 0.01 a |
| Chlorophyll a Fluorescence Parameter | Light Quality | |||
|---|---|---|---|---|
| W z | R | B | RB | |
| F0 | 7954 ± 696 y ab x | 8399 ± 568 a | 7087 ± 326 ab | 6947 ± 443 b |
| FM | 46,827 ± 2489 a | 44,345 ± 1651 a | 43,532 ± 1297 a | 42,145 ± 2878 a |
| FV/FM | 0.83 ± 0.01 ab | 0.81 ± 0.02 b | 0.84 ± 0.00 a | 0.84 ± 0.00 a |
| FM/F0 | 5.90 ± 0.23 ab | 5.30 ± 0.49 b | 6.15 ± 0.12 a | 6.07 ± 0.03 a |
| FV/F0 | 4.90 ± 0.23 ab | 4.30 ± 0.49 b | 5.15 ± 0.12 a | 5.07 ± 0.03 a |
| M0 | 0.59 ± 0.05 ab | 0.76 ± 0.02 a | 0.57 ± 0.05 b | 0.53 ± 0.02 b |
| SM | 382 ± 14 c | 408 ± 19 c | 478 ± 19 b | 533 ± 14 a |
| φE0 | 0.52 ± 0.01 ab | 0.49 ± 0.02 b | 0.54 ± 0.00 a | 0.55 ± 0.02 a |
| φD0 | 0.17 ± 0.01 ab | 0.19 ± 0.02 a | 0.16 ± 0.00 b | 0.17 ± 0.00 ab |
| ψ0 | 0.63 ± 0.01 ab | 0.61 ± 0.02 b | 0.65 ± 0.00 ab | 0.66 ± 0.02 a |
| ABS/RC | 1.93 ± 0.12 ab | 2.29 ± 0.21 a | 1.94 ± 0.16 ab | 1.84 ± 0.15 b |
| ET0/RC | 1.02 ± 0.04 b | 1.13 ± 0.00 a | 1.05 ± 0.03 ab | 1.02 ± 0.06 b |
| TR0/RC | 1.61 ± 0.09 a | 1.92 ± 0.04 b | 1.62 ± 0.04 a | 1.51 ± 0.07 a |
| DI0/RC | 0.33 ± 0.03 b | 0.45 ± 0.03 a | 0.32 ± 0.03 b | 0.30 ± 0.02 b |
| PIABS | 4.44 ± 0.12 b | 2.98 ± 0.67 c | 4.89 ± 0.21 ab | 5.76 ± 0.08 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; Thi, L.T.; Park, Y.G.; Jeong, B.R. Light Quality Affects Growth and Physiology of Carpesium triste Maxim. Cultured In Vitro. Agriculture 2020, 10, 258. https://doi.org/10.3390/agriculture10070258
Zhao J, Thi LT, Park YG, Jeong BR. Light Quality Affects Growth and Physiology of Carpesium triste Maxim. Cultured In Vitro. Agriculture. 2020; 10(7):258. https://doi.org/10.3390/agriculture10070258
Chicago/Turabian StyleZhao, Jin, Luc The Thi, Yoo Gyeong Park, and Byoung Ryong Jeong. 2020. "Light Quality Affects Growth and Physiology of Carpesium triste Maxim. Cultured In Vitro" Agriculture 10, no. 7: 258. https://doi.org/10.3390/agriculture10070258
APA StyleZhao, J., Thi, L. T., Park, Y. G., & Jeong, B. R. (2020). Light Quality Affects Growth and Physiology of Carpesium triste Maxim. Cultured In Vitro. Agriculture, 10(7), 258. https://doi.org/10.3390/agriculture10070258