Impact of Ag Nanoparticles on Seed Germination and Seedling Growth of Green Beans in Normal and Chill Temperatures


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Seed Treatment with AgNPs
2.2. Laboratory Experiment
2.3. Field Experiments
2.4. Statistical Analysis
3. Results and Discussion
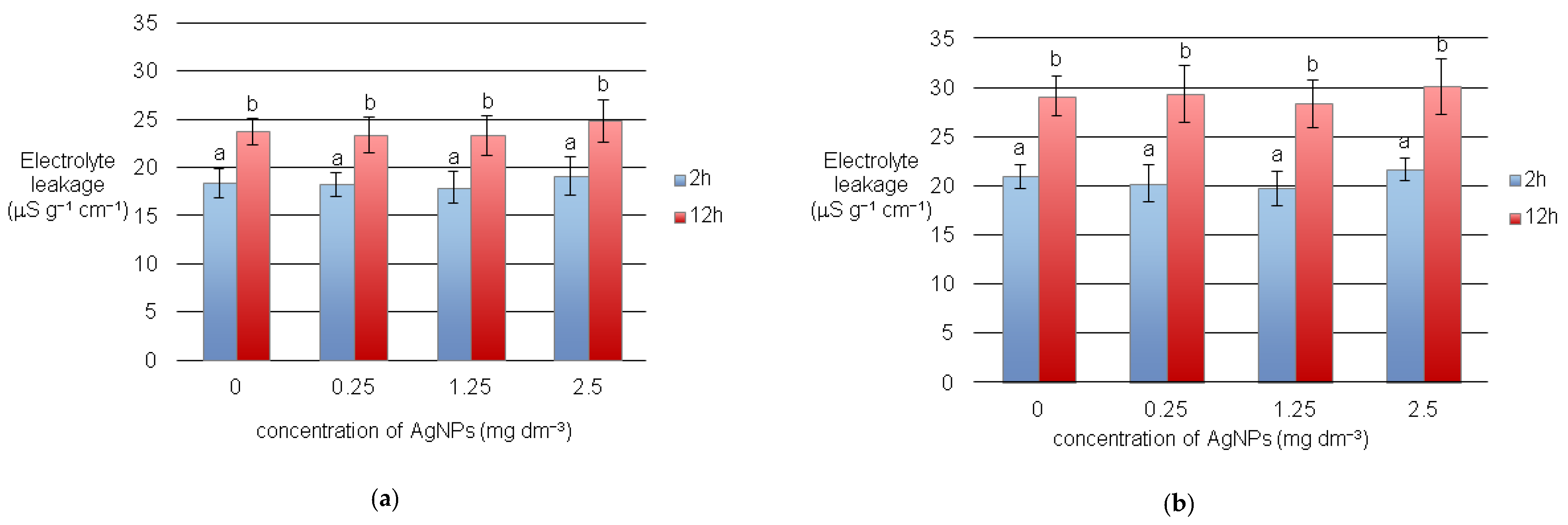
3.1. Electrolyte Leakages and Seed Germination
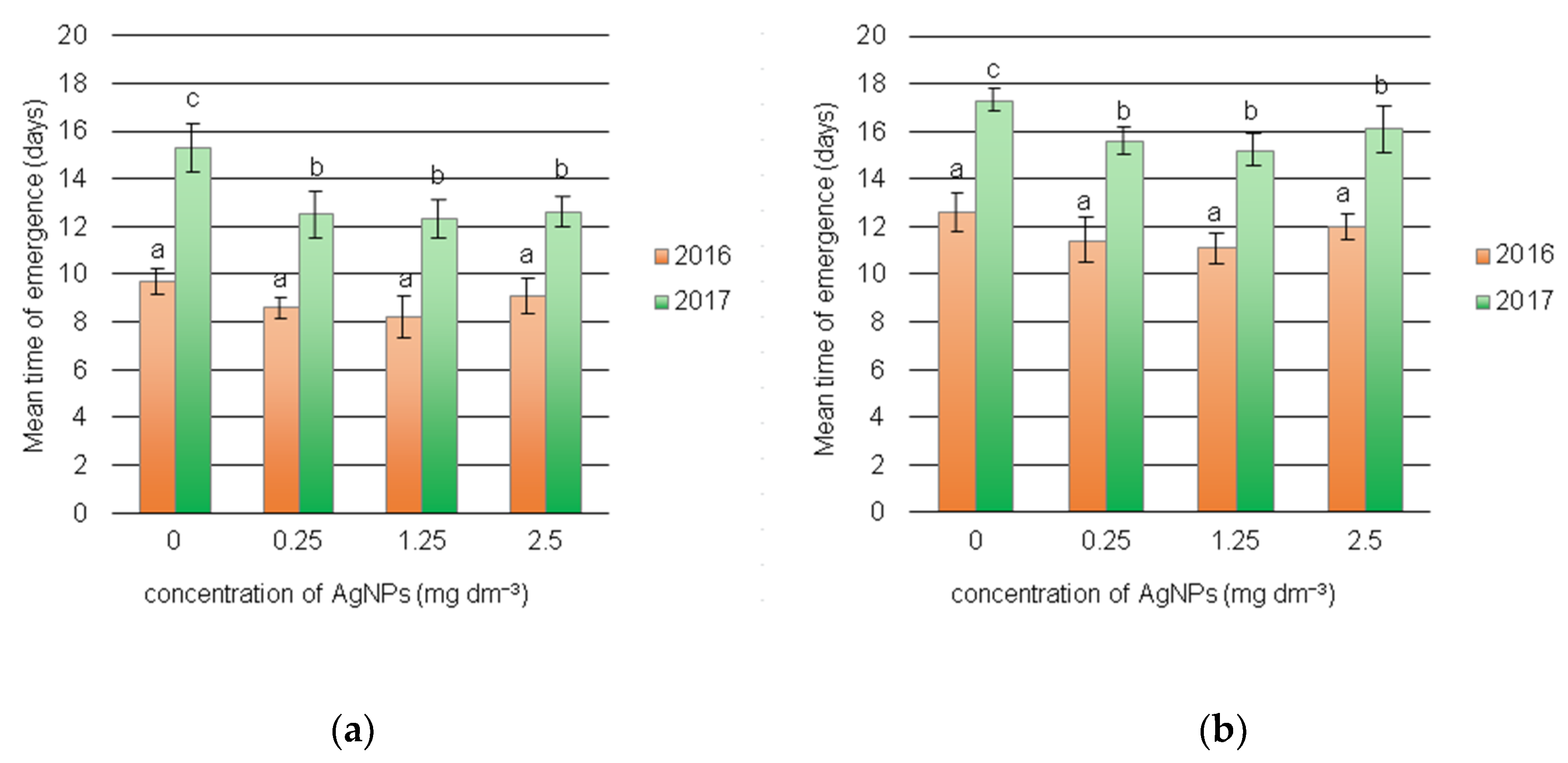
3.2. Dynamics of Field Emergence of Seedlings
3.3. Biometric and Physiological Parameters of Seedlings
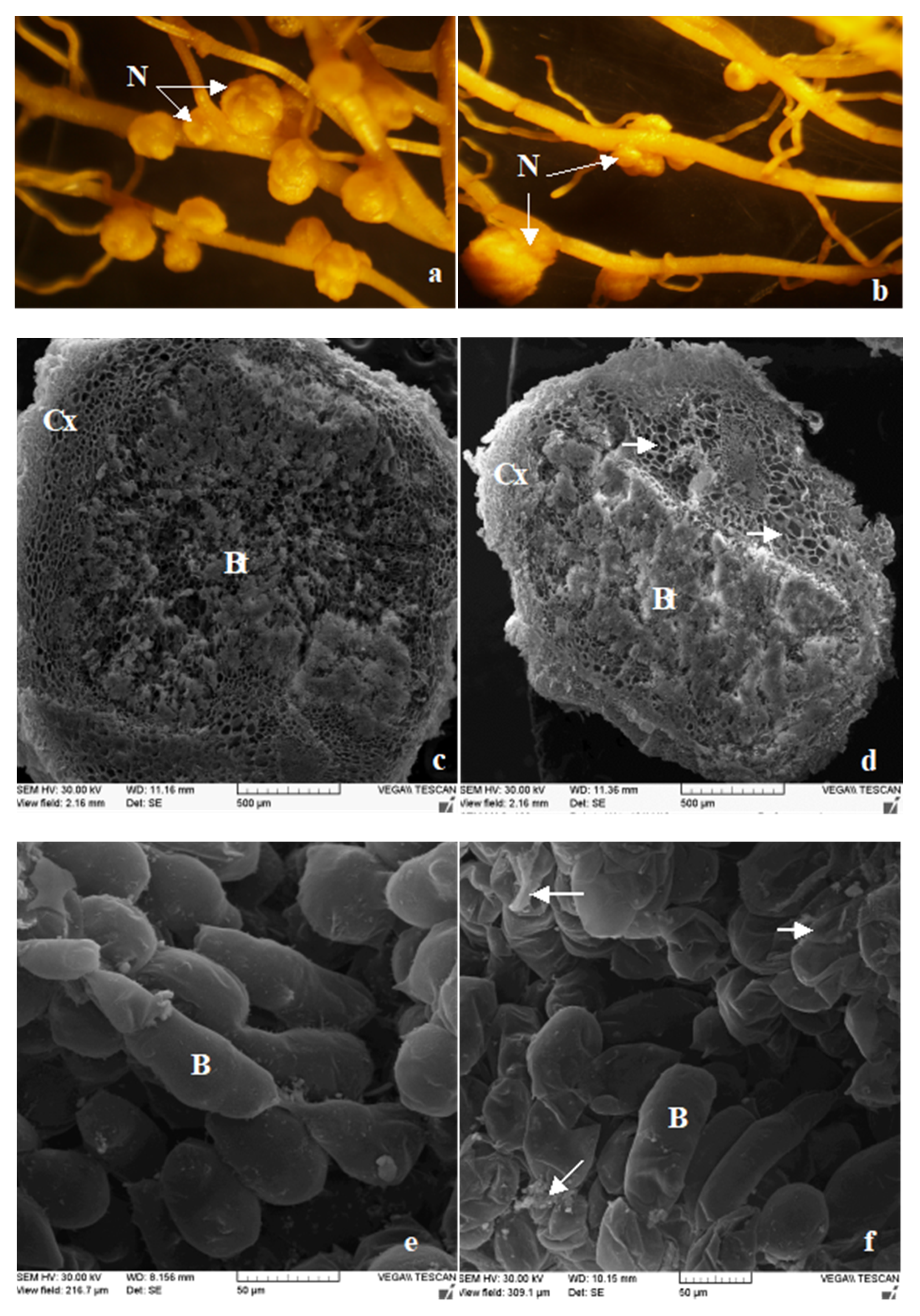
3.4. The Number and Structure of Root Nodules
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- El-Tohamy, W.A.; Singer, S.M.; El-Behairy, U.A.; Abou-Hadid, A.F. Effects of low tunnels, plastic much and mineral nutrient treatments on chilling tolerance of snap bean plants. Acta Hortic. 2001, 559, 127–134. [Google Scholar] [CrossRef]
- Głowacka, A.; Klikocka, H.; Onuch, J. Content of zinc and iron in common bean seeds (Phaseolus vulgaris L.) in different weed control methods. J. Elem. 2015, 20, 293–303. [Google Scholar]
- Abou-Zeid, H.M.; Moustafa, Y. Physiological and cytogenetic responses of wheat and barley to silver nanopriming treatment. Int. J. Appl. Biol. Pharm. Technol. 2014, 5, 265–278. [Google Scholar]
- Feregrino-Perez, A.A.; Magaña-López, E.; Guzmán, C.; Esquivel, K. A general overview of the benefits and possible negative effects of the nanotechnology in horticulture. Sci. Hort. 2018, 238, 126–137. [Google Scholar] [CrossRef]
- European Commission. Recommendation 2011/696/ EU of 18 October 2011 on the Definition of Nanomaterial; OJEU L 275/38; European Commission Recommendation: Brussel, Belgium, 2011; p. 3. [Google Scholar]
- Jo, D.H.; Lee, T.G.; Kim, J.H. Nanotechnology and nanotoxicology in retinopathy. Int. J. Mol. Sci. 2011, 12, 8288–8301. [Google Scholar] [CrossRef] [Green Version]
- Michałek, S.; Święciło, A.; Molas, J. Effect of silver nanoparticles and ions on seeds epiphytic microorganisms activity and early stages of sweetcorn development. Przem. Chem. 2018, 97, 1654–1658. [Google Scholar]
- Servin, A.; Elmer, W.; Mukherjee, A.; De La Torre Roche, R.; Hamdi, H.; White, J.; Bindraban, P.S.; Dimkpa, C. A review of the use of engineered nanomaterials to suppress plant disease and enhance crop yield. J. Nanopart. Res. 2015, 17, 92. [Google Scholar] [CrossRef]
- Zalewska, E.D.; Machowicz-Stefaniak, Z.; Król, E.D. Antifungal activity of nanoparticles against chosen species of the pathogenic fungi towards caraway. Acta Sci. Pol. Hortorum Cultus 2016, 15, 121–137. [Google Scholar]
- Almutairi, Z.; Alharbi, A. Effect of silver nanoparticles on seed germination of crop plants. J. Adv. Agric. 2015, 4, 280–285. [Google Scholar] [CrossRef]
- Sharma, P.; Bhatt, D.; Zaidi, M.G.; Saradhi, P.P.; Khanna, P.K.; Arora, S. Silver nanoparticle mediated enhancement in growth and antioxidant status of Brassica juncea. Appl. Biochem. Biotechnol. 2012, 167, 2225–2233. [Google Scholar] [CrossRef]
- Yin, L.; Colman, B.; McGill, B.; Wright, J.; Bernhardt, E. Effects of silver nanoparticle exposure on germination and early growth of eleven wetland plants. PLoS ONE 2012, 7, e47674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salama, H.M.H. Effects of silver nanoparticles in some crop plants, common bean (Phaseolus vulgaris L.) and corn (Zea mays L.). Int. Res. J. Biotech. 2012, 3, 190–197. [Google Scholar]
- Savithramma, N.; Ankanna, S.; Bhumi, G. Effect of nanoparticles on seed germination and seedling growth of Boswellia Ovalifoliolata—An endemic and endangered medicinal tree Taxon. Nano Vis. 2012, 2, 61–68. [Google Scholar]
- Parveen, A.; Rao, S.O. Effect of nanosilver on seed germination and seedling growth in Pennisetum glaucum. J. Clust. Sci. 2015, 26, 693–701. [Google Scholar] [CrossRef]
- Abou-Zeid, H.; Ismail, G. The role of priming with biosynthesized silver nanoparticles in the response of Triticum aestivum L. to salt stress. Egypt. J. Bot. 2018, 58, 73–85. [Google Scholar]
- Bhati-Kushwaha, H.; Kaur, A.; Malik, C.P. The synthesis and role of biogenic nanoparticles in overcoming chilling stresses. Ind. J. Plant Sci. 2013, 2, 54–62. [Google Scholar]
- Almutairi, Z.M. Influence of silver nano-particles on the salt resistance of tomato (Solanum lycopersicum) during germination. Int. J. Agric. Biol. 2016, 18, 449–457. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, M.; Raja, N.; Mashwani, Z.R.; Hussain, M.; Ejaz, M.; Yasmeen, F. Effect of silver nanoparticles on growth of wheat under heat stress. Iran. J. Sci. Technol. Trans. A Sci. 2019, 43, 287. [Google Scholar] [CrossRef]
- Kumari, M.; Mukherjee, A.; Chandrasekaran, N. Genotoxicity of silver nanoparticles in Alium cepa. Sci. Total Environ. 2009, 407, 5243–5246. [Google Scholar] [CrossRef]
- Kaveh, R.; Li, Y.S.; Ranjbar, S.; Tehrani, R.; Brueck, C.L.; Van Aken, B. Changes in Arabidopsis thaliana gene expression in response to silver nanoparticles and silver ions. Environ. Sci. Technol. 2013, 47, 10637–10644. [Google Scholar] [CrossRef]
- Wang, J.; Koo, Y.; Alexander, A.; Yang, Y.; Westerhof, S.; Zhang, Q.B.; Shnoor, J.L.; Colvin, V.L.; Brasm, J.; Alvarez, P.J.J. Phytostimulation of poplars and Arabidopsis exposed to silver nanoparticles and Ag+ at sublethal concentrations. Environ. Sci. Technol. 2013, 47, 5442–5449. [Google Scholar] [CrossRef] [PubMed]
- Vannini, C.; Domingo, G.; Onelli, E.; Prinsi, B.; Marsoni, M.; Espen, L.; Bracale, M. Morphological and protreomic responses of Eruca sativa exposed to silver nanparticles or silver nitrate. PLoS ONE 2013, 8, e68752. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Leu, Y.R.; Aitken, R.J.; Riediker, M. Inventory of engineered nanoparticle-containing consumer products available in the Singapore retail market and likelihood of release into the aquatic environment. Int. J. Environ. Res. Public Health 2015, 12, 8717–8743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurer-Jones, M.; Gunsolus, I.; Murphy, C.; Haynes, C. Toxicity of engineered nanoparticles in the environment. Anal. Chem. 2013, 85, 3036–3049. [Google Scholar] [CrossRef] [Green Version]
- Pyatenko, A.; Yamaguchi, M.; Suzuki, M. Synthesis of spherical silver nanoparticles with controllable sizes in aqueous solutions. J. Phys. Chem. 2007, 111, 7910–7917. [Google Scholar] [CrossRef]
- ISTA International Seed Testing Association. International Rules for Seed Testing; International Seed Testing Association: Bassersdorf, Switzerland, 2014. [Google Scholar]
- Maguire, J.D. Speed of germination—Aid in selection and evaluation for seedling emergence and vigor. Crop Sci. 1962, 2, 176–177. [Google Scholar] [CrossRef]
- Piekutowska, M.; Szparaga, A. Evaluation of germmination of Soy beans treated with natural environmental friendly extracts depending on the method of their appplication. Annu. Set Environ. Prot. 2018, 20, 375–391. [Google Scholar]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Kolasinska, K.; Szyrmer, J.; Dul, S. Relationship between laboratory seed quality tests and field emergence of common bean seed. Crop Sci. 2000, 40, 470–475. [Google Scholar] [CrossRef]
- Campos, P.S.; Quartin, V.; Ramalho, J.C.; Nunes, M.A. Electrolyte leakage and lipid degradation account for cold sensitivity in leaves of Coffea sp. plants. J. Plant Physiol. 2003, 160, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.S.; Qiu, X.N.; Li, G.B.; Li, W.; Yin, L.Y. Silver nanoparticles induced accumulation of reactive oxygen species and alteration of antioxidant systems in the aquatic plant Spirodela polyrhiza. Environ. Toxicol. Chem. 2014, 33, 1398–1405. [Google Scholar] [CrossRef] [PubMed]
- Mahakham, W.; Sarmah, A.K.; Maensiri, S.; Theerakulpisut, P. Nanopriming technology for enhancing germination and starch metabolism of aged rice seeds using phytosynthesized silver nanoparticles. Sci. Rep. 2017, 7, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Yago, J.I.; Roh, J.H.; Bae, S.D.; Yoon, Y.N.; Kim, H.J.; Nam, M.H. The effect of seed-borne mycoflora from sorghum and foxtail millet seeds on germination and disease transmission. Mycobiology 2011, 39, 206–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hojjat, S.S. Impact of silver nanoparticles on germinated fenugreek seed. Int. J. Agric. Crop. Sci. 2015, 8, 627–630. [Google Scholar]
- Rajjou, L.; Duval, M.; Gallardo, K.; Catusse, J.; Bally, J.; Job, C.; Job, D. Seed germination and vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Arora, R. Priming memory invokes seed stress-tolerance. Environ. Exp. Bot. 2013, 94, 33–45. [Google Scholar] [CrossRef]
- Panyuta, O.; Belava, V.; Fomaidi, S.; Kalinichenko, O.; Volkogon, M.; Taran, N. The effect of pre-sowing seed treatment with metal nanoparticles on the formation of the defensive reaction of wheat seedlings infected with the eyespot causal agent. Nanoscale Res. Lett. 2016, 11, 92. [Google Scholar] [CrossRef] [Green Version]
- Maurel, C.; Boursiac, Y.; Luu, D.T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef]
- Kibinza, S.; Bazin, J.; Bailly, C.; Farrant, J.M.; Corbineau, F.; El-Maarouf-Bouteau, H. Catalase is a key enzyme in seed recovery from ageing during priming. Plant Sci. 2011, 181, 309–315. [Google Scholar] [CrossRef]
- Shams, G.; Ranjbar, M.; Amiri, A. Effect of silver nanoparticles on concentration of silver heavy element and growth indexes in cucumber (Cucumis sativus L. negeen). J. Nanopart. Res. 2013, 15, 1630–1642. [Google Scholar] [CrossRef]
- Ashrafi, M.; Sarajuoghi, M.; Mohammadi, K.; Zarei, S. Effect of nanosilver application on agronomic traits of soybean in relation to different fertilizers and weed density in field conditions. Environ. Exper. Biol. 2013, 11, 53–58. [Google Scholar]
- Moghaddam, M.N.; Sabzevar, A.H.; Mortazaei, Z. Impact of ZnO and silver nanoparticles on legume-Sinorhizobium symbiosis. Adv. Stud. Biol. 2017, 9, 83–90. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Tripathi, A.; Singh, S.; Singh, Y.; Vishwakarma, K.; Yadav, G.; Sharma, S.; Singh, V.K.; Mishra, R.K.; Upadhyay, R.G.; et al. Uptake, accumulation and toxicity of silver nanoparticle in autotrophic plants, and heterotrophic microbes: A concentric review. Front. Microbiol. 2017, 8, 7. [Google Scholar] [CrossRef] [PubMed]
- Ekhtiyari, R.; Mohebbi, H.; Mansouri, M. The study of the effects of nano silver technology on salinity tolerance of (Foeniculum vulgare Mill.). Plant Ecosyst. 2011, 7, 55–62. [Google Scholar]
- Ekhtiyari, R.; Moraghebi, F. The study of the effects of nano silver technology on salinity tolerance of cumin seed (Cuminum cyminum L.). Plant Ecosyst. 2011, 25, 99–107. [Google Scholar]
- Shojaei, T.R.; Salleh, M.A.M.; Tabatabaei, M.; Mobli, H.; Aghbashlo, M.; Rashid, S.A.; Tan, T. Applications of nanotechnology and carbon nanoparticles in agriculture. In Synthesis, Technology and Applications of Carbon Nanomaterials; Suraya, A.R., Raja, N.I.R.O., Mohd, Z.H., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 247–277. [Google Scholar]
- Vance, C.P. Legume symbiotic nitrogen fixation: Agronomic aspects. In The Rhizobiaceae; Spaink, H.P., Kondorosi, A., Hooykaas, P.J.J., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998; pp. 509–530. [Google Scholar]
- Pérez Guerra, J.C.; Coussens, G.; De Keyser, A.; De Rycke, R.; De Bodt, S.; Van De Velde, W.; Goormachtig, S.; Holsters, M. Comparison of developmental and stress-induced nodule senescence in Medicago truncatula. Plant Physiol. 2010, 152, 1574–1584. [Google Scholar] [CrossRef] [Green Version]
- Liang, Z.H.; Das, A.; Hu, Z.Q. Bacterial response to a shock load of nanosilver in an activated sludge treatment system. Water Res. 2010, 44, 5432–5438. [Google Scholar]
- Chavan, S.; Nadanathangam, V. Effects of nanoparticles on plant growth-promoting bacteria in Indian agricultural soil. Agronomy 2019, 9, 140. [Google Scholar] [CrossRef] [Green Version]
- Abd-Alla, M.H.; Nafady, N.A.; Khalaf, D.M. Assessment of silver nanoparticles contamination on faba bean Rhizobium leguminosarum bv. Viciae—Glomus aggregatum symbiosis: Implications for induction of autophagy process in root nodule. Agric. Ecosyst. Environ. 2016, 218, 163–177. [Google Scholar] [CrossRef]
- Zhang, L.; Wu, L.; Si, Y.; Shu, K. Size-dependent cytotoxicity of silver nanoparticles to Azotobacter vinelandii: Growth inhibition, cell injury, oxidative stress and internalization. PLoS ONE 2018, 13, e0209020. [Google Scholar] [CrossRef] [Green Version]
- Michels, C.; Perazzoli, S.; Soares, H.M. Inhibition of an enriched culture of ammonia oxidizing bacteria by two different nanoparticles: Silver and magnetite. Sci. Total Environ. 2017, 586, 995–1002. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Fan, W.; Kishen, A.; Gutmann, J.L.; Fan, B. Evaluation of the antibacterial efficacy of silver nanoparticles against Enterococcus faecalis biofilm. J. Endod. 2014, 40, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.K.; Koo, H.C.; Kim, K.W.; Shin, S.; Kim, S.H.; Park, Y.H. Antibacterial activity and mechanism of action of the silver ion in Staphylococcus aureus and Escherichia coli. Appl. Environ. Microbiol. 2008, 74, 2171–2178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirzajani, F.; Ghassempour, A.; Aliahmadi, A.; Esmaeili, M.A. Antibacterial effect of silver nanoparticles on Staphylococcus aureus. Res. Microbiol. 2011, 162, 542–549. [Google Scholar] [CrossRef]
- Durán, N.; Durán, M.; de Jesus, M.B.; Seabra, A.B.; Fávaro, W.J.; Nakazato, G. Silver nanoparticles: A new view on mechanistic aspects on antimicrobial activity. Nanomedicine 2016, 12, 789–799. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Year | pH | C-org% | N-NH4 | N-NO3 | P | K | Ca | Mg |
|---|---|---|---|---|---|---|---|---|
| mg kg−1 Air Dry Mass | ||||||||
| 2016 | 6.8 | 2.4 | 15 | 9 | 17 | 71 | 647 | 25 |
| 2017 | 7.1 | 2.2 | 19 | 12 | 21 | 63 | 784 | 29 |
| Month | Year | Temperature (°C) | Rainfall (mm) | The Occurrence of Chill Period |
|---|---|---|---|---|
| May | 2016 | 15.6 | 32.7 | No |
| 2017 | 12.4 | 46.2 | Yes * | |
| June | 2016 | 17.6 | 57.3 | No |
| 2017 | 18.2 | 12.4 | No |
| Cultivar | Temperature | Treatment | |||
|---|---|---|---|---|---|
| Control | AgNPs (mg dm−3) | ||||
| 0.25 | 1.25 | 2.5 | |||
| Germination rate (%) | |||||
| Bali | Normal | 61.2 a | 68.7 b | 72.6 c | 69.4 b |
| Chill | 29.6 a | 40.1 b | 45.3 c | 38.6 b | |
| Delfina | Normal | 67.8 a | 75.3 b | 80.2 c | 72.6 b |
| Chill | 33.5 a | 49.5 b | 45.9 b | 40.3 c | |
| Germination capacity (%) | |||||
| Bali | Normal | 91.6 a | 90.9 a | 92.2 a | 90.5 a |
| Chill | 59.8 a | 69.1 b | 69.6 b | 62.4 a | |
| Delfina | Normal | 95.2 a | 94.2 a | 95,5 a | 93.4 b |
| Chill | 61.2 a | 71.4 b | 72.7 b | 64.6 a | |
| Percentage of fungus infections | |||||
| Bali | Normal | 10.4 a | 6.7 b | 6.1 b | 3.6 c |
| Chill | 3.6 a | 3.1 a | 2.5 b | 1.7 c | |
| Delfina | Normal | 7.4 a | 5.7 b | 3.6 c | 2.3 d |
| Chill | 2.2 a | 1.6 b | 1.5 b | 0.5 c | |
| Cultivar | Year | Treatments | |||
|---|---|---|---|---|---|
| Control | AgNPs (mg dm−3) | ||||
| 0.25 | 1.25 | 2.5 | |||
| Height of plants (cm) | |||||
| Bali | 2016 | 12.6 a | 13.6 b | 14.4 c | 13.3 b |
| 2017 | 9.7 a | 12.2 b | 11.8 b | 10.1 a | |
| Delfina | 2016 | 15.2 a | 17.4 b | 18.1 c | 15.3 a |
| 2017 | 13.6 a | 16.7 b | 16.0 b | 13.9 a | |
| Fresh mass of shoots (g plant−1) | |||||
| Bali | 2016 | 9.5 a | 9.9 a | 11.2 b | 9.35 a |
| 2017 | 7.9 a | 8.85 b | 9.9 c | 8.1 a | |
| Delfina | 2016 | 11.8 a | 13.5 b | 14.1 b | 9.2 d |
| 2017 | 8.6 a | 11.3 b | 11.5 b | 8.6 a | |
| Dry mass of shoots (g plant−1) | |||||
| Bali | 2016 | 0.77 a (8.1) * | 0.81 a (8.2) | 0.92 b (8.2) | 0.67 c (7.2) |
| 2017 | 0.73 a (9.2) | 0.79 b (8.9) | 0.85 c (8.6) | 0.57 d (7.0) | |
| Delfina | 2016 | 1.04 a (8.8) | 1.17 b (8.7) | 1.21 b (8.6) | 0.78 c (8.5) |
| 2017 | 0.81 a (9.4) | 0.95 b (8.4) | 0.98 b (8.5) | 0.66 c (7.7) | |
| Number of root nodules per plant | |||||
| Bali | 2016 | 20.4 a | 21.2 b | 21.8 b | 18.6 c |
| 2017 | 16.3 a | 18.2 b | 17.5 c | 12.3 d | |
| Delfina | 2016 | 24.7 a | 27.6 b | 28.2 b | 23.7 a |
| 2017 | 19.5 a | 22.8 b | 22.9 b | 15.7 c | |
| Cultivar | Year | Treatments | |||
|---|---|---|---|---|---|
| Control | AgNPs (mg dm−3) | ||||
| 0.25 | 1.25 | 2.5 | |||
| Leaf chlorophyll (a + b) content (mg g−1 f.w.) | |||||
| Bali | 2016 | 1.99 a | 2.04 a | 2.01 a | 1.97 a |
| 2017 | 2.02 a | 2.16 b | 2.19 b | 1.94 b | |
| Delfina | 2016 | 2.21 a | 2.22 a | 2.25 a | 2.09 b |
| 2017 | 2.07 a | 2.34 b | 2.39 b | 2.06 a | |
| Stomata conductance (mol H2O m−2 s−1) | |||||
| Bali | 2016 | 0.75 a | 0.76 a | 0.83 a | 0.84 a |
| 2017 | 0.62 a | 0.73 b | 0.77 b | 0.71 b | |
| Delfina | 2016 | 1.03 a | 1.02 a | 1.04 a | 0.98 a |
| 2017 | 0.74 a | 0.86b | 0.86 b | 0.80 b | |
| Net photosynthesis intensity (µmol CO2 m−2 s−1) | |||||
| Bali | 2016 | 12.45 a | 13.51 a | 14.85 b | 12.06 a |
| 2017 | 11.07 a | 12.96 b | 13.18 b | 11.22 a | |
| Delfina | 2016 | 13.68 a | 14.38 b | 15.02 c | 13.56 a |
| 2017 | 11.89 a | 13.14 b | 13.77 c | 12.17 a | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prażak, R.; Święciło, A.; Krzepiłko, A.; Michałek, S.; Arczewska, M. Impact of Ag Nanoparticles on Seed Germination and Seedling Growth of Green Beans in Normal and Chill Temperatures. Agriculture 2020, 10, 312. https://doi.org/10.3390/agriculture10080312
Prażak R, Święciło A, Krzepiłko A, Michałek S, Arczewska M. Impact of Ag Nanoparticles on Seed Germination and Seedling Growth of Green Beans in Normal and Chill Temperatures. Agriculture. 2020; 10(8):312. https://doi.org/10.3390/agriculture10080312
Chicago/Turabian StylePrażak, Roman, Agata Święciło, Anna Krzepiłko, Sławomir Michałek, and Marta Arczewska. 2020. "Impact of Ag Nanoparticles on Seed Germination and Seedling Growth of Green Beans in Normal and Chill Temperatures" Agriculture 10, no. 8: 312. https://doi.org/10.3390/agriculture10080312
APA StylePrażak, R., Święciło, A., Krzepiłko, A., Michałek, S., & Arczewska, M. (2020). Impact of Ag Nanoparticles on Seed Germination and Seedling Growth of Green Beans in Normal and Chill Temperatures. Agriculture, 10(8), 312. https://doi.org/10.3390/agriculture10080312