The Screening of Resistance against Meloidogyne graminicola in Oats


Abstract
:1. Introduction
2. Materials and Methods
2.1. Nematode Inoculum
2.2. Plant Materials
2.3. Attraction and Penetration Study
2.4. Inoculation Study
2.5. Evaluation of Resistance and Susceptibility of Oat Lines
2.6. Data Analysis
3. Results
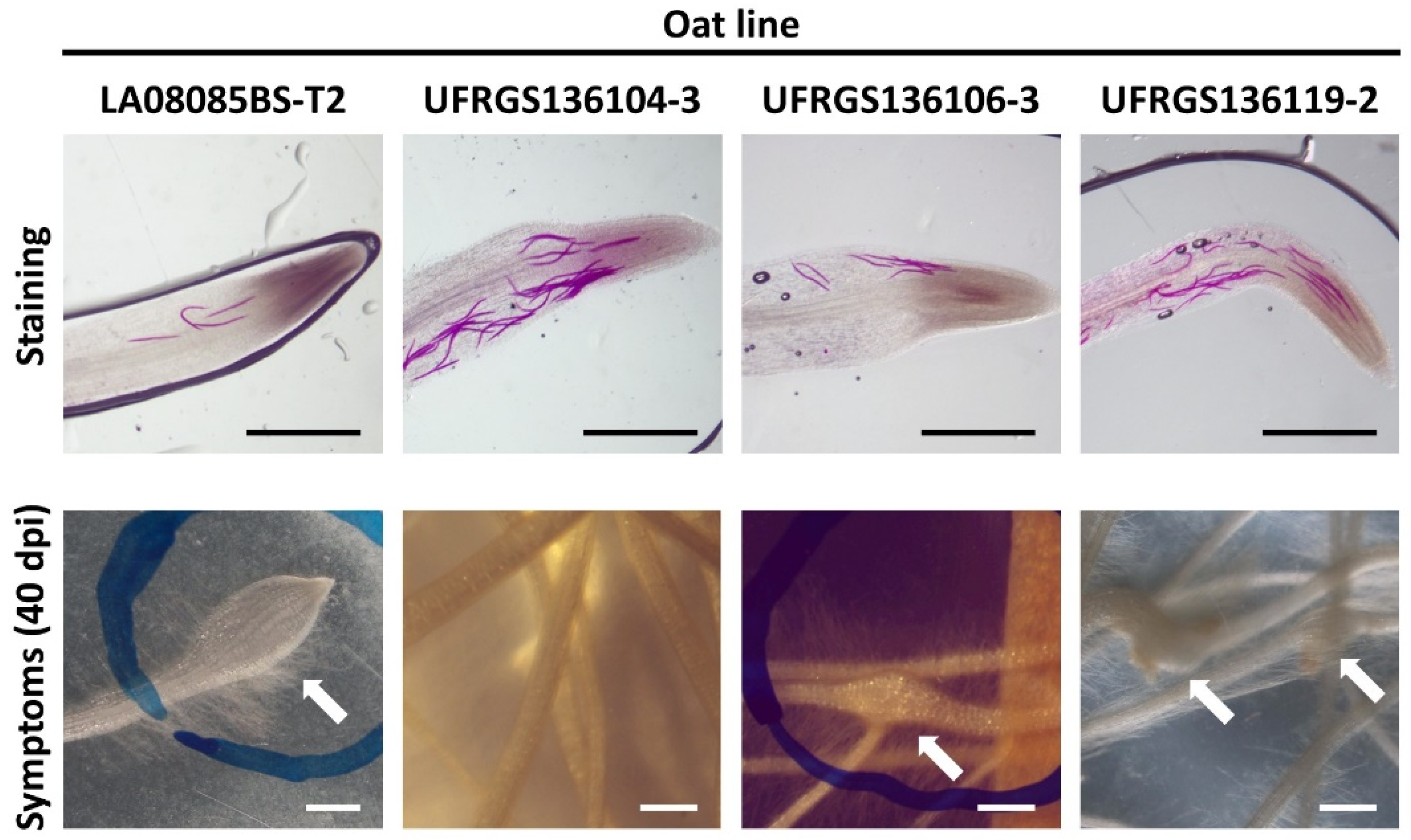
3.1. Attraction and Penetration Study
3.2. Inoculation Study
3.3. Resistance/Susceptibility of Oats Evaluation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vaish, S.; Pandey, S. Root-knot disease caused by Meloidogyne graminicola: A limiting factor for growth and yield of barley (Hordeum vulgare L.). Curr. Nematol. 2012, 23, 7–12. [Google Scholar]
- Plowright, R.A.; Bridge, J. Effect of Meloidogyne graminicola (Nematoda) on the establishment, growth and yield of rice cv. IR36. Nematologica 1990, 36, 81–89. [Google Scholar] [CrossRef]
- Soriano, I.R.S.; Prot, J.-C.; Matias, D.M. Expression of tolerance for Meloidogyne graminicola in rice cultivars as affected by soil type and flooding. J. Nematol. 2000, 32, 309–317. [Google Scholar]
- Duan, H.W. The Survey, Identification and Etiology Studies of Rice Root-Knot Nematode, Meloidogyne graminicola, in Taiwan. Master’s Thesis, National Chung Hsing University, Taichung, Taiwan, 2015. [Google Scholar]
- Mantelin, S.; Bellafiore, S.; Kyndt, T. Meloidogyne graminicola: A major threat to rice agriculture. Mol. Plant. Pathol. 2017, 18, 3–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bureau of Animal and Plant Health Inspection and Quarantine (BAPHIQ). Available online: pesticide.baphiq.gov.tw (accessed on 5 July 2020).
- Rao, Y.S.; Prasad, J.S.; Panwar, M.S. Nematode problems in rice: Crop losses, symptomatology and management. In Plant Parasitic Nematodes of India: Problems and Progress; Swarup, G., Dasgupta, D.R., Eds.; Indian Agricultural Research Institute: New Delhi, India, 1986; pp. 279–299. [Google Scholar]
- Yang, J.I.; Stadler, M.; Chuang, W.Y.; Wu, S.; Ariyawansa, H.A. In vitro inferred interactions of selected entomopathogenic fungi from Taiwan and eggs of Meloidogyne graminicola. Mycol. Prog. 2020, 19, 97–109. [Google Scholar] [CrossRef]
- Suttie, J.M.; Reynolds, S.G. Fodder Oats: A World Overview; Food and Agriculture Organization of the United Nations: Rome, Italy, 2004. [Google Scholar]
- Huang, C.W.; Liang, W.H.; Esvelt Klos, K.; Chen, C.S.; Huang, Y.F. Evaluation of agronomic performance and exploratory genome-wide association study of a diverse oat panel for forage use in Taiwan. Grassl. Sci. 2020. [Google Scholar] [CrossRef]
- Wang, S.H.; Shy, Y.M.; Chen, Y.H.; Chang, C.T.; Lee, K.H.; Jea, Y.S. Effect of oat silage substitute on lactation performance of Holstein lactating cows. J. Taiwan Livest. Res. 2018, 51, 109–115. [Google Scholar]
- Tateishi, Y.; Iwahori, H.; Uesugi, K.; Katsura, M. Invasion, development, and reproduction of 3 Meloidogyne species on oat cultivar Tachiibuki, a nematode-suppressive fall crop. Jpn. J. Nematol. 2012, 41, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Tateishi, Y.; Uesugi, K.; Iwahori, H. Host suitability of a common oat cultivar Sniper for five plant-parasitic nematode species. Jpn. J. Nematol. 2016, 46, 21–24. [Google Scholar] [CrossRef] [Green Version]
- Andersen, S.; Andersen, K. Suggestions for determination and terminology of pathotypes and genes for resistance in cyst-forming nematodes, especially Heterodera avenae 1. EPPO Bull. 1982, 12, 379–386. [Google Scholar] [CrossRef]
- Mai, W.; Mullin, P.G. Plant.-Parasitic Nematodes: A Pictorial Key to Genera, 5th ed.; Cornell University Press: Ithaca, NY, USA, 1996. [Google Scholar]
- Padgham, J.; Sikora, R. Biological control potential and modes of action of Bacillus megaterium against Meloidogyne graminicola on rice. Crop. Prot. 2007, 26, 971–977. [Google Scholar] [CrossRef]
- Bybd, D.W., Jr.; Kirkpatrick, T.; Barker, K.R. An improved technique for clearing and staining tissues for detection of nematodes. J. Nematol. 1983, 15, 142–143. [Google Scholar] [PubMed]
- Lasserre, F.; Rivoal, R.; Cook, R. Interactions between Heterodera avenae and Pratylenchus neglectus on wheat. J. Nematol. 1994, 26, 336. [Google Scholar] [PubMed]
- Dongre, M.; Simon, S. Infestation and host studies of Meloidogyne graminicola in rice: Nurseries of Allahabad. Int. J. Bot. Res. 2013, 3, 27–34. [Google Scholar]
- Taylor, A.L.; Sasser, J.N. Biology, Identification and Control. of Root-Knot Nematodes (Meloidogyne Species); Department of Plant Pathology, North Carolina State University Graphics and the USAID: Raleigh, NC, USA, 1978; p. 111. [Google Scholar]
- Sasser, J.C.; Carter, C.C.; Hartman, K.M. Standardization of Host Suitability Studies and Reporting of Resistance to Root-Knot Nematode; United States Agency for International Development (USAID): Raleigh, NC, USA, 1985; p. 7.
- Reynolds, A.M.; Dutta, T.K.; Curtis, R.H.C.; Powers, S.J.; Gaur, H.S.; Kerry, B.R. Chemotaxis can take plant-parasitic nematodes to the source of a chemoattractant via the shortest possible routes. J. R. Soc. Interface 2011, 8, 568–577. [Google Scholar] [CrossRef] [Green Version]
- Cabasan, M.T.N.; Kumar, A.; De Waele, D. Comparison of migration, penetration, development and reproduction of Meloidogyne graminicola on susceptible and resistant rice genotypes. Nematology 2012, 14, 405–415. [Google Scholar] [CrossRef]
- Pedrosa, E.M.R.; Hussey, R.S.; Boerma, H.R. Penetration and post-infectional development and reproduction of Meloidogyne arenaria races 1 and 2 on susceptible and resistant soybean genotypes. J. Nematol. 1996, 28, 343–351. [Google Scholar]
- Andwar, S.A.; McKenry, M.V. Penetration, development and reproduction of Meloidogyne arenaria on two new resistant Vitis spp. Nematropica 2000, 30, 9–17. [Google Scholar]
- Pegard, A.; Brizzard, G.; Fazari, A.; Soucaze, O.; Abad, P.; Djian-Caporalino, C. Histological characterization of resistance to different root-knot nematode species related to phenolics accumulation in Capsicum annuum. Phytopathology 2005, 95, 158–165. [Google Scholar] [CrossRef] [Green Version]
- Cabasan, M.T.N.; Kumar, A.; Bellafiore, S.; De Waele, D. Histopathology of the rice root-knot nematode, Meloidogyne graminicola, on Oryza sativa and O. glaberrima. Nematology 2014, 16, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Andersen, S. Resistens mod Havreål Heterodera avenae; I Kommission hos Dansk Videnskabs Forlag: Copenhagen, Denmark, 1961. [Google Scholar]
- Clamot, G.; Rivoal, R. Genetic resistance to cereal cyst nematode Heterodera avenae WOLL. in wild oat Avena sterilis I. 376. Euphytica 1984, 33, 27–32. [Google Scholar] [CrossRef]
- Ireholm, A. Characterization of pathotypes of cereal cyst nematodes, Heterodera spp., in Sweden. Nematologica 1994, 40, 399–411. [Google Scholar] [CrossRef]
- Barr, A.R.; Chalmers, K.J.; Karakousis, A.; Kretschmer, J.M.; Manning, S.; Lance, R.C.M.; Lewis, J.; Jeffries, S.P.; Langridge, P. RFLP mapping of a new cereal cyst nematode resistance locus in barley. Plant. Breed. 1998, 117, 185–187. [Google Scholar] [CrossRef]
- McIntosh, R.A.; Yamazaki, Y.; Dubcovsky, J.; Rogers, J.; Morris, C.; Somers, D.J.; Appels, R.; Devos, K.M. Catalogue of gene symbols for wheat. National BioResource Project, Komugi-Wheat Genetic Resources Database. Available online: https://shigen.nig.ac.jp/wheat/komugi/genes/download.jsp (accessed on 11 November 2019).
- Soriano, I.R.; Asenstorfer, R.E.; Schmidt, O.; Riley, I.T. Inducible flavone in oats (Avena sativa) is a novel defense against plant-parasitic nematodes. Phytopathology 2004, 94, 1207–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendy, H.; Dalmasso, A.; Cardin, M.C. Differences in resistant Capsicum annuum attacked by different Meloidogyne species. Nematologica 1985, 31, 72–78. [Google Scholar] [CrossRef]
- Kaloshian, I.; Yaghoobi, J.; Liharska, T.; Hontelez, J.; Hanson, D.; Hogan, P.; Jesse, T.; Wijbrandi, J.; Simons, G.; Vos, P.; et al. Genetic and physical localization of the root-knot nematode resistance locus Mi in tomato. Mol. Genet. Genom. 1998, 3, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Djian-Caporalino, C.; Fazari, A.; Arguel, M.J.; Vernie, T.; VandeCasteele, C.; Faure, I.; Brunoud, G.; Pijarowski, L.; Palloix, A. Root-knot nematode (Meloidogyne spp.) Me resistance genes in pepper (Capsicum annuum L.) are clustered on the P9 chromosome. Theor. Appl. Genet. 2007, 114, 473–486. [Google Scholar] [CrossRef]
- Hajihassani, A.; Rutter, W.B.; Luo, X. Resistant Pepper Carrying N, Me1, and Me3 have Different Effects on Penetration and Reproduction of Four Major Meloidogyne species. J. Nematol. 2019, 51, 1–9. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| GI | R | |
|---|---|---|
| 1 | <0.1 | Highly resistant |
| 2 | ≤1 | Resistant |
| 3 | >1 | Moderately susceptible |
| 4 | >1 | Susceptible |
| 5 | >10 | Highly susceptible |
| Oat Line | Root Gall Formation | Nematode Reproduction | |||
|---|---|---|---|---|---|
| Total Number of Root Galls | Numbers of Incompletely Developed Root Galls | Numbers of Completely Developed Root Galls | Total Number of Eggs | Numbers of Harvested J2 | |
| LA08085BS-T2 | 17 ± 6.26 a | 11.6 ± 4.5 a | 5.4 ± 2.2 a | 325.27 ± 131.96 a | 203.4 ± 160.55 a |
| UFRGS136104-3 | 0.00 ± 0.00 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.00 ± 0.00 b | 11 ± 4.65 b |
| UFRGS136106-3 | 3 ± 0.71 b | 1.6 ± 0.7 b | 1.4 ± 0.9 b | 2.2 ± 1.16 b | 0.00 ± 0.00 b |
| UFRGS136119-2 | 2.8 ± 0.86 b | 1.4 ± 0.9 b | 1.4 ± 0.6 b | 58.4 ± 25.96 b | 125 ± 64.02 a |
| Oat Line | Leaf Weight (g) | Root Weight (g) | ||||
|---|---|---|---|---|---|---|
| Inoculated | Control | Inoculated | Control | |||
| LA08085BS-T2 | 0.138 ± 0.006 | 0.132 ± 0.008 | NS | 0.431 ± 0.017 | 0.365 ± 0.035 | NS |
| UFRGS136104-3 | 0.141 ± 0.005 | 0.151 ± 0.006 | NS | 0.589 ± 0.039 | 0.704 ± 0.051 | NS |
| UFRGS136106-3 | 0.197 ± 0.004 | 0.188 ± 0.013 | NS | 0.723 ± 0.053 | 0.701 ± 0.049 | NS |
| UFRGS136119-2 | 0.144 ± 0.015 | 0.151 ± 0.008 | NS | 0.523 ± 0.062 | 0.475 ± 0.034 | NS |
| Oat Line | Root Galls Index 1 | Multiplication Factor 2 | Resistance/Susceptibility 3 |
|---|---|---|---|
| LA08085BS-T2 | 2.80 ± 0.37 a | 2.64 ± 1.36 a | MS |
| UFRGS136104-3 | 1.00 ± 0.00 b | 0.06 ± 0.02 b | HR |
| UFRGS136106-3 | 2.00 ± 0.00 b | 0.01 ± 0.01 b | R |
| UFRGS136119-2 | 1.80 ± 0.2 b | 0.92 ± 0.41 a | R |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, S.-M.; Huang, Y.-F.; Yang, J.-I. The Screening of Resistance against Meloidogyne graminicola in Oats. Agriculture 2020, 10, 352. https://doi.org/10.3390/agriculture10080352
Yao S-M, Huang Y-F, Yang J-I. The Screening of Resistance against Meloidogyne graminicola in Oats. Agriculture. 2020; 10(8):352. https://doi.org/10.3390/agriculture10080352
Chicago/Turabian StyleYao, Shun-Min, Yung-Fen Huang, and Jiue-In Yang. 2020. "The Screening of Resistance against Meloidogyne graminicola in Oats" Agriculture 10, no. 8: 352. https://doi.org/10.3390/agriculture10080352
APA StyleYao, S.-M., Huang, Y.-F., & Yang, J.-I. (2020). The Screening of Resistance against Meloidogyne graminicola in Oats. Agriculture, 10(8), 352. https://doi.org/10.3390/agriculture10080352