1. Introduction
Heat stress significantly restricts crop growth and compromises plant production around the world despite the several mechanisms evolved by the plants in order to alleviate adverse environmental conditions [
1,
2]. According to climate reports, average global temperature has been rising over the last century and at the current rates of greenhouse gas emissions, a further increase is projected by the end of the 21st century [
3]. Interestingly, due to expected increases in cloudiness and concomitant decreases in heat radiant loss, night temperatures are anticipated to rise faster and to a larger extent compared to daytime temperatures [
4]. Higher than optimum temperatures during the day and their negative effects on plant physiology, biochemistry, and yield have been extensively investigated [
1,
5,
6]. Substantial variability among species and genotypes has been recorded [
5,
7]; however, the response of heat tolerant varieties to high night temperatures has not been tested.
A negative correlation between high night temperatures and yield has been revealed by past research on several crops such as wheat (
Triticum aestivum L.) [
8], soybean (
Glycine max L.) [
9], sorghum (
Sorghum bicolor L.) [
10], cowpea (
Vigna unguiculata L.) [
11], and rice (
Oryza sativa L.) [
12]. Additionally, it has been reported that when high night temperatures coincide with the plant’s reproductive stage, their effect on yield becomes detrimental due to increases in rates of floral abscission, male sterility, and floral bud suppression [
8,
13,
14].
Cotton (
Gossypium hirsutum L.) may be considered a well-adapted crop to hot and semi-arid areas, since it originates from such areas; however, higher than optimum day temperatures have been shown to adversely affect its physiology and yield due to decreases in leaf photosynthetic rates and increases in respiration that result in limited carbohydrate availability to reproductive structures and consequently yield reductions through increased fruit abscission and reduced fruit set, as well as decreased boll size and seed number [
15,
16]. Similar to those observations, yield reductions due to lower boll setting [
17], higher fruit shedding rates [
18], and reduced number of seeds per locule and boll [
19] have been reported for cotton when increased night temperatures occurred during its reproductive phase. Further research additionally revealed that high night temperatures increased cotton leaf respiration rates, with concomitant decreases in leaf photosynthetic rates, when the stress was applied during their early reproductive period [
20].
In order to safeguard cotton production against the adverse effects of climate change, a concerted effort has been undertaken for breeding for heat tolerance, and several cotton cultivars, able to maintain their yield output under heat stress conditions, have been selected through the use of physiological, biochemical, and morphological parameters [
15,
21,
22,
23,
24]. As such, heat tolerance in cotton has been associated with maintenance of leaf photosynthesis [
22,
23], especially of the subtending to a flower/boll leaf [
25], and sufficient supply of assimilates to the reproductive structures, reduced leaf electrolyte leakage [
26], as well as increased pre-stress antioxidant capacity and maintenance of ATP levels [
23]. Nevertheless, those associations have been evaluated only under conditions of high day temperatures. Taking into consideration that night temperatures are expected to rise faster and to a larger extent than day temperatures, the efficiency of heat-tolerant varieties to heat stress occurring during the night is evidently important to be evaluated in order to provide breeders with a potentially new selection target. To that end, the objectives of this study were to monitor the effects of high night temperatures on the subtending leaf’s photosynthesis and respiration, electrolyte leakage, and ATP levels, as well as on the levels of the antioxidant glutathione reductase, and soluble carbohydrate content of the flower and its subtending leaf of two cotton cultivars differing in heat-tolerance. It was hypothesized that the contrast in their thermotolerance cultivars would differ in their physiological and biochemical responses to high night temperatures and that would ultimately result in significant differences in their number of reproductive structures and total reproductive dry weight.
2. Materials and Methods
Growth chamber studies were conducted and repeated at the Altheimer Laboratory, University of Arkansas. Cotton (
Gossypium hirsutum L.) cultivar ST5288B2F, a standard cultivar widely planted in the cotton belt, and VH260, a thermotolerant cultivar from Pakistan [
22], were planted in 2 L pots containing horticulture mix #2 (SunGro Distribution Inc., Bellevue, WA, USA). Pots were arranged in two growth chambers (Conviron PGW36, Conviron Inc., Winnipeg, Canada) that were equipped with incandescent and fluorescent lamps and set for a 14 h photoperiod with a photosynthetic flux density (PPFD) of 800–850 µmol/m
2s and a relative humidity of 60%. Half-strength Hoagland’s nutrient solution was applied daily to all pots in order to maintain adequate nutrients and water. Plants were grown until the flowering stage (approximately 8 weeks after planting) under normal day/night temperatures of 32/24 °C (maximum during the day, minimum during the night, respectively) simulating a normal diurnal variation. At flowering, the night temperatures on one of the two chambers were raised to 30 °C for 10 h at the start of the dark period (2000 h–0600 h) with a gradual decrease to 24 °C, for an overall duration of two weeks, while the control plants remained under normal conditions (32/24 °C). First position white flowers and their subtending leaves (LSWF) from main-stem nodes 8 and 10 were collected at the end of the first and second week, respectively, for determination of soluble carbohydrates, glutathione reductase, adenosine triphosphate (ATP), and electrolyte leakage. The study was repeated twice. In order to account for any possible differences between growth chambers, the growth chamber used for the high night temperature treatment (32/30 °C, day/night) in the first study was used for the control treatment (32/24 °C, day/night) in the second study. No significant effect on any of the parameters monitored was found between the two studies (
p > 0.05), and the data were combined from the two consecutive studies for statistical analysis.
2.1. Leaf Gas Exchange Measurements
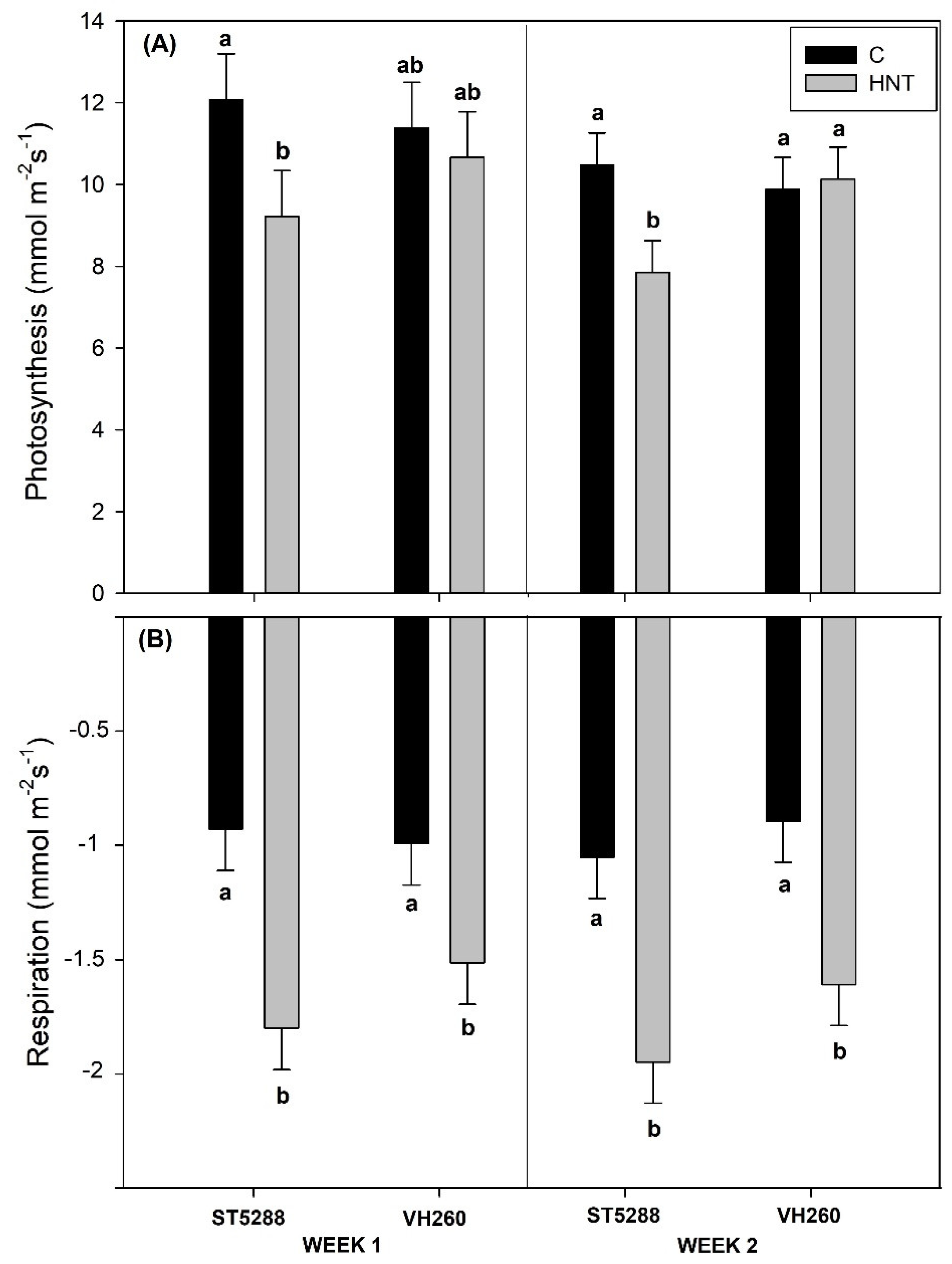
A CI-340 handheld photosynthesis system (CID Bioscience, Inc., Camas, WA, USA) was used to determine net photosynthetic and respiratory rates for the attached subtending to the first position flower leaf from the 8th and 10th node of each plant. Measurements of photosynthesis and respiration were taken at 1200 h and 2300 h, respectively, and the results were expressed as mmol m−2 s−1. Measurements of respiration were taken 3 h into the dark period in order to avoid the respiratory burst that follows the end of the light period.
2.2. Leaf Electrolyte Leakage Measurements
Electrolyte leakage was measured according to Martineau et al. [
27] with modifications. Five disks were punched from similar interveinal areas of the leaf subtending a flower and placed in closed vials filled with 10 mL of double deionized water and incubated at 25 °C for 48 h in the dark. After this period, the electrical conductivity of the solution (L1) of all samples was determined using a single probe conductivity meter (Thermo Orion Model 115, Beverly, MA, USA). Subsequently, the vials were placed in a boiling water bath for 10 min to obtain the maximum leakage of the leaf disks. Electrical conductivity was measured again (L2) after the samples equilibrated at 25 °C, and the electrolyte leakage was estimated as EL% = (L1/L2) × 100.
2.3. Leaf Adenosine Triphosphate (ATP) Measurements
Three leaf disks per each subtending to a first position white flower from each plant were excised using a cork borer (1 cm in diameter) at the end of the first and seventh night. The leaf disks were placed immediately in tubes containing 5 mL TRIS solution, and the tubes were put in a boiling water bath for 10 min for the extraction of ATP. The resulting aliquot was then stored in 1.5 mL microcentrifuge tubes at −80 °C. The substrate–enzyme complex of firefly luciferin–luciferase (ATP bioluminescent assay kit, Sigma Chemical Company, St. Louis, MO, USA) that converts the chemical energy associated with ATP into light was used, and the light produced (proportional to the ATP content of the sample) was determined with a 20/20n luminometer (Turner BioSystems Inc., Sunnyvale, CA, USA) and the help of a standard curve. The results were expressed as µg ATP m−2.
2.4. Glutathione Reductase (GR) Measurements
Glutathione reductase was extracted and measured from first position white flowers and their subtending leaves at the end of the first and second week after initiation of stress according to the methods described by Loka and Oosterhuis [
28]. Excised pistil and leaf tissues were homogenized using a mortar and pestle in an ice-cold extraction solution (10× tissue fresh weight, about 0.1 g for the pistil, about 0.5 g for the leaf) comprised of 50 mM PIPES (1,4-piperazinediethanesulfonic acid) buffer (pH 6.8), 6 mM cysteine hydrochloride, 10 mM d-isoascorbate, 1 mM ethylenediaminetetraacetic acid, 0.3% Triton X-100, and 1% (w/v) soluble polyvinylpyrrolidone (PVP). Solutions were further blended for 1 min in a tube containing 0.25 g insoluble PVP and 1 drop of antifoam A emulsion using a homogenizer (Model Polytron; Brinkman Instruments Inc., Palo Alto, CA, USA). Subsequently, samples were centrifuged at 12,000 rpm for 20 min (4 °C), and the supernatants were stored at −80 °C for further determination of glutathione reductase content. To each well of a 96-well microtitration plate, a 15.7 μL aliquot of enzyme extract from each sample was added to a 300 μL reaction solution containing 50 mM Tris-HCl buffer (pH = 7.5), 0.15 mM reduced nicotinamide adenine dinucleotide phosphate (NADPH), 0.5 mM oxidized glutathione, and 3 mM MgCl
2. Oxidation of NADPH was determined as the decrease in absorbance at 340 nm during a 1 min reaction time and was measured using an Ascent Multiscan microplate reader (Molecular Devices Corporation, Sunnyvale, CA, USA), and glutathione reductase activity was expressed as GR units gr
−1 FW (fresh weight).
2.5. Determination of Soluble Carbohydrates of Cotton Flowers and their Subtending Leaves
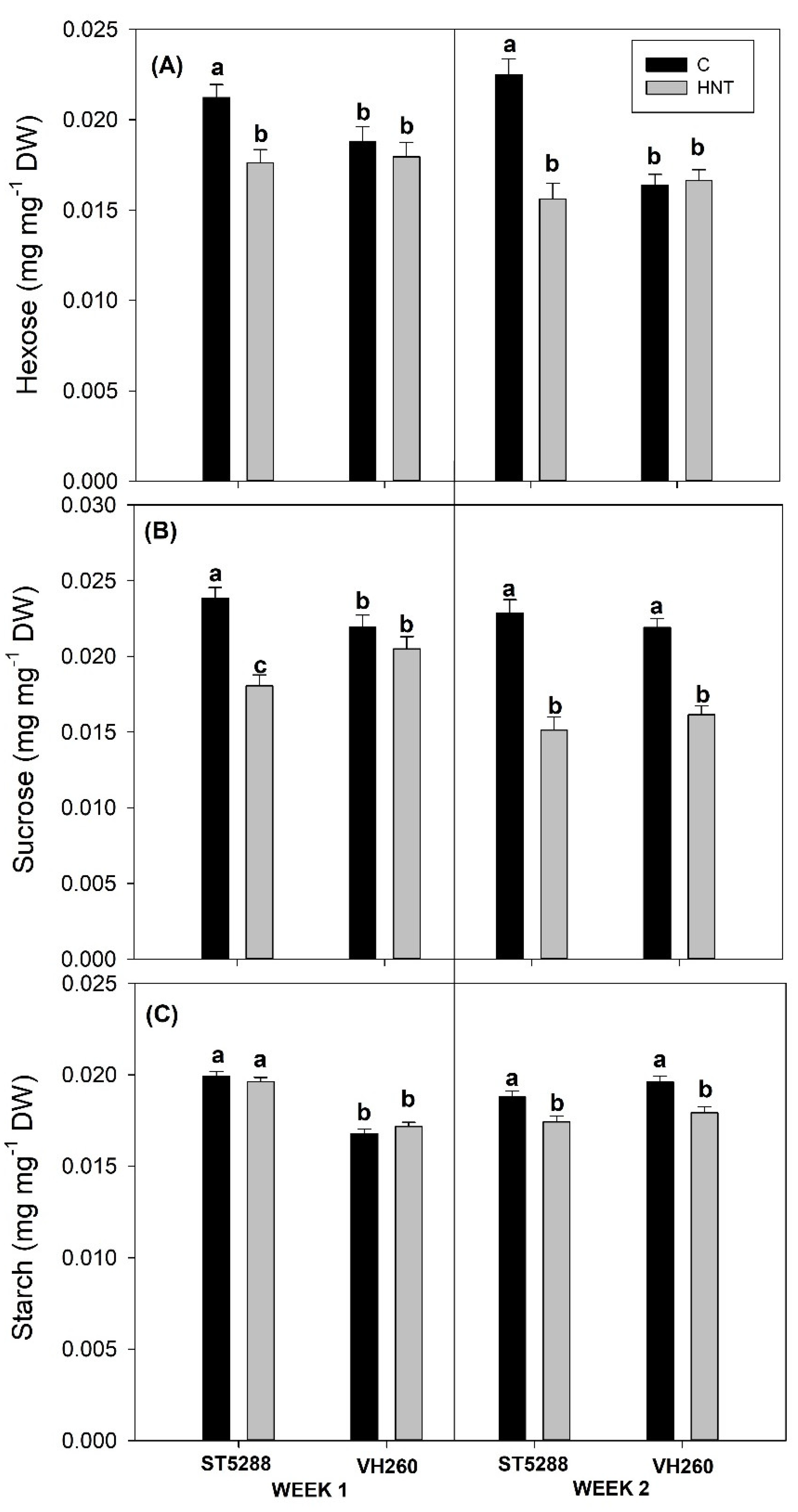
First fruiting position cotton flowers from the 8th and 10th nodes of each plant and their subtending leaves were sampled at the end of the first and second week. Soluble carbohydrate content of the pistils and their subtending leaves was measured according to the method described by Loka and Oosterhuis [
28]. The tissues (leaves and excised pistils) were oven dried for 3 days at 50 °C and then ground with a mortar and pestle. Then 40 mg of the ground tissue were extracted 3 times with 80 °C aqueous ethanol (800 mL ethanol/L), and the samples were centrifuged after each extraction at 5000 rpm. Finally the fractions were pooled, and the remaining tissue pellet was kept for extraction and determination of starch. Active charcoal (30 mg) was then added to the pooled fractions in order to remove substances that could interfere with the carbohydrate measurements, and the samples were centrifuged again at 3500 rpm for 15 min at 4 °C. The supernatant was immediately stored at −80 °C for further determination of sucrose, glucose, and fructose with a MultiScan Ascent microplate reader (Thermo Fisher Scientific Inc., Waltham, MA, USA). The glucose (HK) assay kit from Sigma (Sigma Chemical Company, St. Louis, MO, USA) was used. A 20 µL aliquot of each extract was pipetted into a well of a microtiter plate, and the plate was incubated at 50 °C for 40 min to evaporate ethanol. Ten microliters of water were then added to each well along with 100 µL of glucose assay reagent (included in the assay kit), and the plate was incubated again for 15 min at 30 °C. The absorbance was measured three times at 340 nm using a microplate reader. Ten microliters of phosphoglucose isomerase (0.25 EU) were added to the extracts in each well of the plate, and the absorbance was again measured at 340 nm. Subsequently, 10 µL of invertase (83 EU) was added to the extracts, and the microtiter plate was incubated at 30 °C for 60 min. The absorbance was measured three times at 340 nm, and the results were expressed in mg carbohydrate/mg dry weight with the help of a standard curve made of known glucose concentrations (0.4, 0.1, 0.01, 0.001, 0.0001 mL glucose mL
−1). The results were expressed as mg/mg DW (dry weight). The remaining tissue pellet was used for starch extraction, and 0.5 mL of 1M KOH was added into each sample. The samples were then placed in a boiling water bath for 60 min, and starch was hydrolyzed after additions of β-amylase and amyloglucosidase according to methods described by Zhao et al. [
29]. After centrifugation of the samples at 14,000 rpm for 10 min, the extracts were brought to a final volume of 1.5 mL with double deionized water. A 20 µL aliquot of each extract was pipetted into a well of a microtiter plate, and the glucose concentrations were determined according to the methods used to quantify glucose in the soluble carbohydrate extractions. Glucose content was multiplied by 0.9 to account for water loss after glucose units were connected to form starch [
29]. The results were expressed as mg mg
−1 DW (dry weight).
2.6. Dry Weight and Number of Reproductive Structures
The number of reproductive structures per plant, including flower buds, flowers, and young bolls, was recorded at the end of the experiment. Dry weight per reproductive structure and total dry weight of reproductive structures (total reproductive dry weight) were measured and expressed as g.
2.7. Statistical Analysis
The experimental design was a two-factor factorial, the two factors being cultivar and temperature, with thirty replications in each temperature treatment and fifteen replications for each cultivar in the same temperature treatment. The trends of the two growth chamber studies were similar, and the results were pooled and analyzed using a Student’s t-test. Means were considered significantly different at p ≤ 0.05.
4. Discussion
Higher than optimum temperatures have long been known to adversely affect several physiological and metabolic processes with detrimental effects on plant growth and yield. To that end, extensive efforts have been undertaken and heat-tolerant cultivars have been introduced [
21,
22,
23,
24]; however, their efficiency against high night temperatures has not been studied. Respiration, a key metabolic process closely related to crop growth, has long been known for its sensitivity to heat stress [
30] and increased respiration rates under higher than optimum temperatures have been reported in a variety of crops, including cotton [
8,
9,
10,
13]. Studies, however, have suggested that cultivars that can maintain low respiration rates under conditions of heat stress, either through more efficient energy use or reduced maintenance respiration, are more heat tolerant [
31,
32]. In contrast to those studies, high night temperatures in our experiments increased respiration rates of both cultivars, indicating that VH260 was unable to prevent increases in its respiration rates. In contrast, VH260 was able to maintain its net photosynthetic rates, while high night temperatures markedly decreased net photosynthetic rates of ST5288. Photosynthesis is one of the most heat-labile processes in plants due to the sensitivity of PS II, and several researchers have reported the adverse effect higher than optimum day temperatures has on photosynthesis [
1,
22]. In cotton, however, Schrader et al. [
33] observed that heating dark-adapted leaves resulted in decreased photosynthetic efficiency due to an increase in thylakoid permeability. In support of that observation, Prasad et al. [
8,
10], in experiments with wheat and sorghum, reported that high night temperatures decreased photosynthesis and that was attributed to compromises of cell membrane integrity due to over-production of reactive oxygen species (ROS) from increased respiration rates. Decreases in cell membrane integrity appear to be the reason for the decreased net photosynthetic rates of ST5288 observed in our study, since electrolyte leakage, a measure of cell membrane integrity, was substantially increased under high night temperatures stress, indicating significant cell membrane damage [
26,
27]. Contrastingly, electrolyte leakage of VH260 remained unaffected, suggesting that VH260 was able to maintain its cell membrane integrity and as a result managed to prevent decreases in its net photosynthetic rates. Similar to our results, Snider et al. [
22] observed that high day temperatures had no significant effect on net photosynthetic rates of VH260, while significant decreases were observed in their thermosensitive cultivar. According to the authors, that was attributed to the maintenance of high GR levels in VH260 under heat stress conditions that were able to protect the photosynthetic system from damage by reactive oxidative species (ROS). In accordance with their results, GR levels of VH260 in our study remained unaffected for the duration of heat stress. However, even though high night temperatures increased GR levels of ST5288 to similar to VH260 GR levels, they were unable to prevent reductions in cell membrane integrity and net photosynthesis, leading us to speculate that GR had a rather minimal antioxidative capacity under high night temperatures stress, and the increased cell membrane integrity of VH260 was the main reason for the maintenance of photosynthetic rates. Since membrane integrity and thermostability is largely determined by the membrane’s degree of saturation, and GR is not the only antioxidant controlling ROS accumulation. Further research into the cell membrane composition and antioxidant capacity of VH260 could provide a better insight into its increased cell membrane stability.
In addition to the formation of ROS and cell membrane damage, disruptions in photosynthesis and respiration, due to higher than optimum temperatures, have a significant effect on the plant’s energy balance, since both of them are involved in ATP synthesis [
32]. Increases in respiration, which is the main provider of ATP in plants, is a common response to heat stress, since energy costs of repair mechanisms for maintenance of cellular homeostasis are increased [
34,
35]. In our study, higher than optimum temperatures resulted in substantial decreases in LSWF ATP content in both cultivars, despite the significant increases in respiration rates. While the lower LSWF ATP content in ST5288 could be partially attributed to its decreased photosynthetic rates, the similar reductions in LSWF ATP content of VH260 lead us to speculate that the increased energy demands for synthesis of protective compounds [
36,
37], and maintenance of ATP-demanding processes, such as phloem loading [
32], were the main reason for the substantially reduced LSWF ATP levels of both cultivars.
Apart from energy supply, disruptions in photosynthesis and respiration can have a significant effect on leaf carbohydrate content, since they respectively control carbon fixation and utilization [
38,
39]. In our study, LSWF starch and sucrose levels of both cultivars were decreased under conditions of high night temperatures. Comparable decreases were also observed in soybean and were attributed to decreases in leaf net photosynthetic rates [
9], which is in accordance with our findings in ST5288 but not in VH260, whose net photosynthetic rates remained unaltered. Starch, as the main storage form of carbohydrates, is accumulated during the day in order to sustain carbohydrate demand during the night. However, its degradation during the day indicated that photosynthetic rates of VH260 were apparently insufficient to maintain growth under heat stress conditions [
40]. A similar explanation could also be provided for the decreases in LSWF sucrose content of VH260, but taking in consideration the high sensitivity of sucrose metabolizing enzymes to temperature changes [
41] we speculate that the observed reductions could also be due to decreased activities of sucrose synthesizing enzyme sucrose phosphate synthase (SPS) or increased activities of sucrose cleaving enzymes sucrose synthase (SS) and invertase (INV). In support of our speculation, Kaushal et al. [
42] reported that heat-stressed chickpea leaves had lower SPS activity compared to control, while increased INV activities were observed under conditions of heat stress [
43]. In contrast to LSWF sucrose content, decreases in LSWF hexose content were observed only in ST528 but not in VH260. Kaushal et al. [
42] reported similar decreases in hexose content of heat-sensitive chickpea genotypes, and they were attributed to lower photosynthetic rates as well as decreased INV activity, leading us to assume that VH260 was able to maintain its hexose content possibly due to its increased INV activity. However, since activities of sucrose metabolizing enzymes were not quantified in this study, further research is required in order to confirm these assumptions.
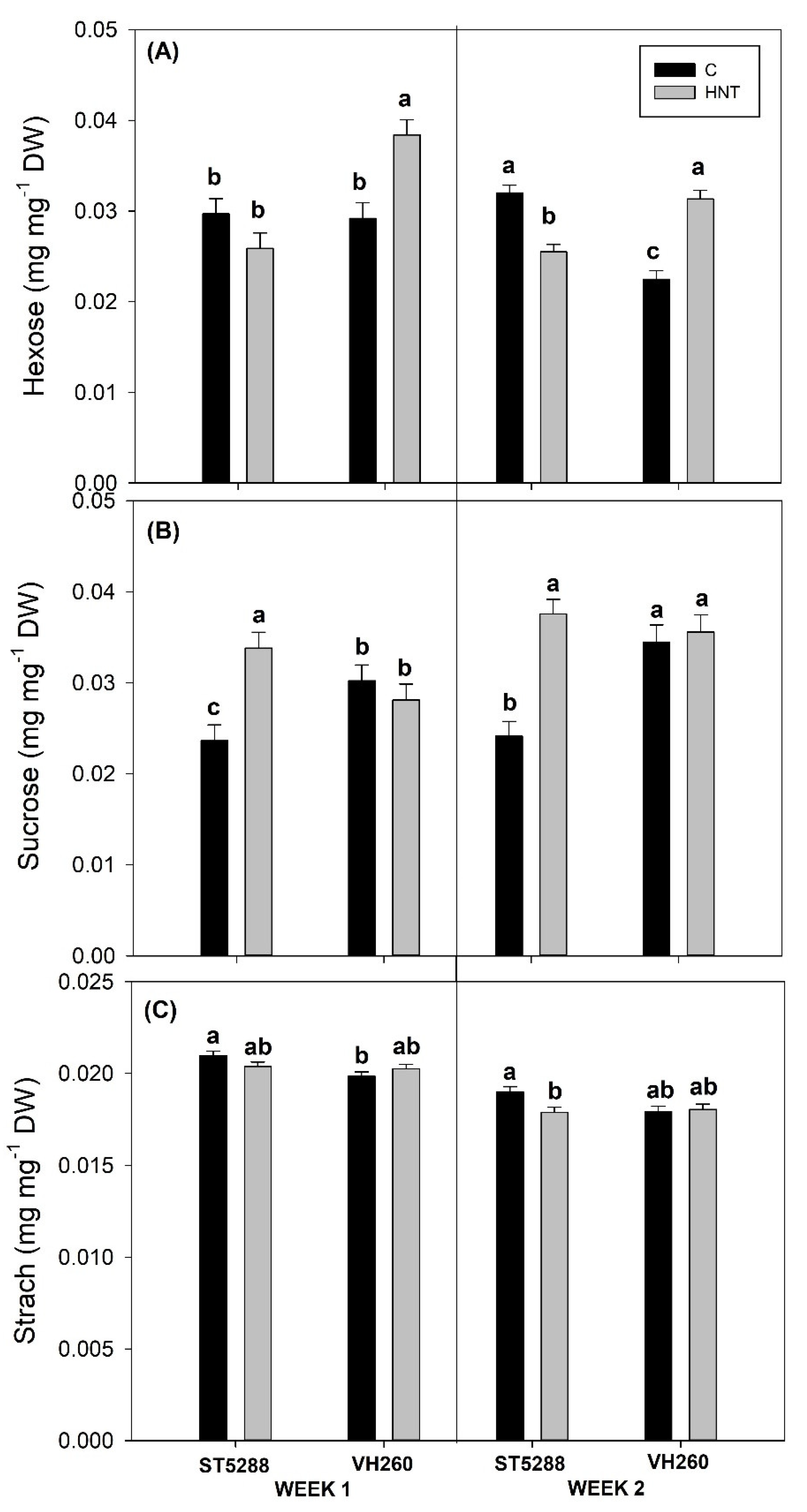
Reproductive success is greatly dependent on leaf carbohydrate supply [
10,
44], as well as on photoassimilate translocation to reproductive structures [
45] and redox homeostasis of them [
44,
45,
46]. According to our results, a marked increase was observed in the sucrose content of ST5288 reproductive structures, whereas hexose and starch contents were significantly decreased. Heat stress has been reported to inhibit assimilate translocation and result in substantial reductions in the carbohydrate content of reproductive structures [
47]. However, this over-accumulation of sucrose, despite the substantial decreases in its LSWF carbohydrate content, suggests that high night temperatures did not have an effect on sucrose translocation. Since assimilate translocation is determined by both sink strength and competition between sinks for assimilates [
48], the lower number of retained reproductive structures in ST5288 would have effectively reduced competition for substrates and hence allow the maintenance of sucrose translocation. Additionally, a decrease in sink strength, indicated by reduced activities of sucrose cleaving enzymes, should also be considered, especially since hexose contents of ST5288 reproductive structures were substantially reduced. Nevertheless, increases in sucrose content have also been associated with the antioxidant role that sucrose assumes under conditions of abiotic stress [
49]. Pistil GR levels of ST5288, in our study, remained unaffected under conditions of high night temperatures, suggesting that the antioxidant mechanism of pistils was rather unresponsive to stress, inducing as a result further starch degradation and over-accumulation of sucrose in the heat-stressed pistils of ST5288. In support of our suggestion, Snider et al. [
44] reported marked increases in GR of heat stressed pistils accompanied by concomitant decreases in pistil sucrose content, which according to the authors indicated a possible connection between those two responses. In contrast to ST5288, pistil sucrose and starch contents of VH260 remained unaffected by the high night temperature regime, whereas pistil hexose content was markedly increased. The lack of an effect of the decreased LSWF sucrose content on pistil sucrose content of VH260, despite the maintenance of the number of reproductive structures, leads us to suggest that VH260 managed to allocate its photoassimilates in a more efficient way compared to ST5288. Furthermore, the increases in pistil hexose content indicated that activities of sucrose-cleaving enzymes, and hence sink strength, was not inhibited under conditions of high night temperatures. Comparable to our results, Kaushal et al. [
42] reported increased hexose content in the heat-stress anthers of heat-tolerant chickpea cultivars, which were attributed to increased INV activities. In contrast to ST5288, pistil GR content of VH260 was markedly increased under conditions of high night temperatures This could be attributed to higher pistil GR observed in VH260; however, further research needs to be dedicated in the responses of antioxidant and carbohydrate metabolism of cotton under conditions of high night temperature stress in order to confirm those assumptions.
Despite the different physiological and biochemical responses observed between the two cultivars in our study, high night temperatures resulted in significant, yet similar, decreases compared to control on total reproductive dry weight of both cultivars. Further analysis, however, revealed that the reductions in total reproductive dry weight of ST5288 were associated with decreases in number of reproductive structures per plant, while in VH260, they were due to decreases in the dry weight per reproductive structure. Abscission of reproductive structures, with the exception of white flowers, in cotton, is a naturally occurring procedure, as the plant tries to balance the sink’s (reproductive structures) demand for photoassimilates with the source (leaf) availability of photoassimilates [
50]. However, abiotic stresses have been shown to exacerbate shedding of reproductive structures, mainly due to reductions in leaf photosynthetic rates that result in perturbations of carbohydrate metabolism and availability of leaves and reproductive structures [
51]. This appears to be the case in our study, where the reductions in LSWF photosynthetic rates of ST5288 were accompanied by decreases in LSWF carbohydrate contents, which led the plants to promptly decrease their number of reproductive structures. A different pattern was observed in VH260, where the maintenance of net photosynthetic rates allowed the plants to retain its LSWF carbohydrate content for longer and prevent abscission of reproductive structures. However, despite the increase in pistil GR levels and maintenance of pistil carbohydrate content, significant decreases were observed in the dry weight per reproductive structure in VH260, confirming the fact that plants that maintain a heavy load of reproductive structures often fail to achieve optimum reproductive structure weight, due to the increased competition between sinks for photoassimilates [
52]. In spite of the similar decreases compared to control on total reproductive dry weight of both cultivars, total reproductive dry weight of VH260 under conditions of high temperatures was significantly higher compared to that of ST5288, indicating that VH260 was more heat tolerant compared to ST5288. Comparable to our results, Xu et al. [
24], in their study with two cotton cultivars differing in thermotolerance and subjected to a short-term (5 days) heat stress during the day, reported that the heat tolerant cultivar had a higher yield compared to the sensitive cultivar. In contrast to our results, that was attributed to the increased boll weight of the heat tolerant cultivar despite the similar abscission rates of both cultivars. We speculate that the differences in stress imposition both in terms of duration and time (day vs. night), as well as the use of different cultivars, are the reasons for the differential responses observed between their study and ours.
In conclusion, high night temperatures resulted in significant decreases in total reproductive weight in the standard cultivar ST5288 but not in the heat-tolerant VH260, indicating that heat tolerant cultivars selected under high day temperatures are also efficient under increased night temperatures. However, taking into consideration the large variability that exists in terms of heat tolerance in cotton, further studies that include more genotypes are required in order to ascertain cotton’s heat tolerance under both high day and night temperature conditions. The results also indicated that net photosynthetic rates and cell membrane integrity could be useful indicators for heat tolerance evaluation under high night temperature stress; however, further research into carbohydrate and antioxidant metabolism of cotton’s reproductive structures would provide useful information into the mechanisms of heat tolerance in cotton.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}