Functional Diversity of Soil Microbial Community after Conversion of a Chestnut Forest to an Agricultural System




Abstract
:
1. Introduction
2. Materials and Methods
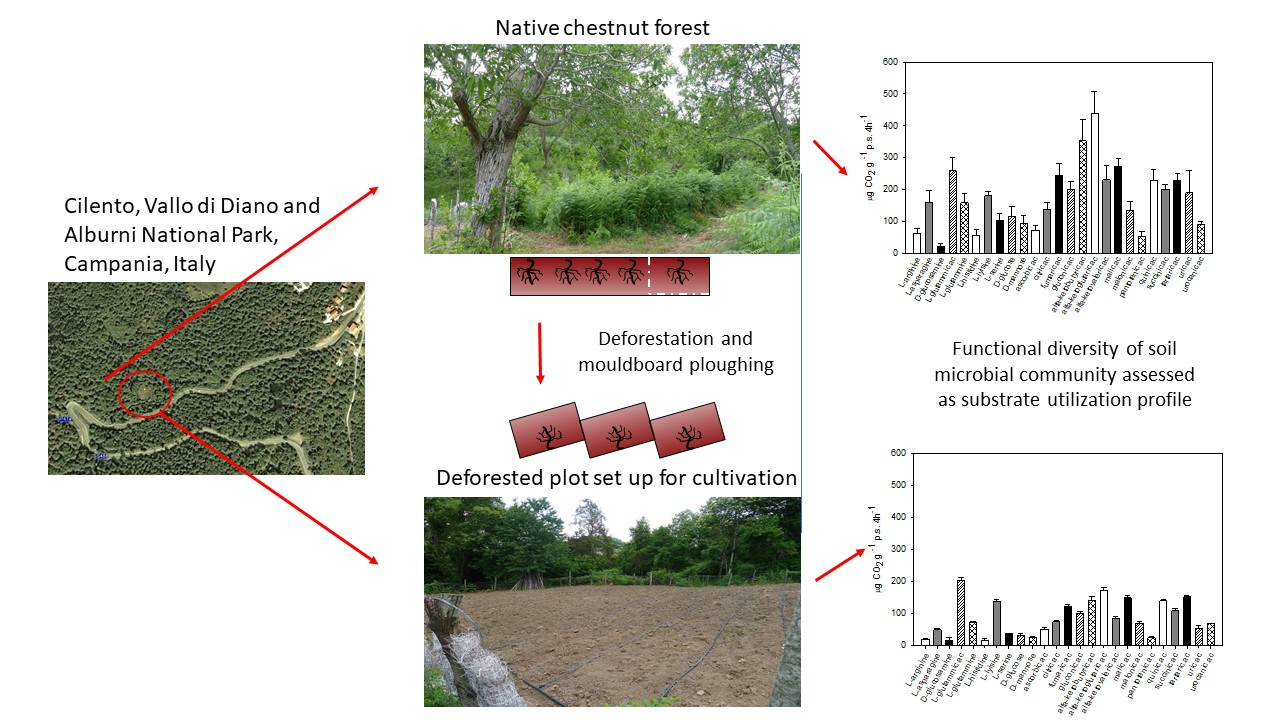
2.1. Site Description and Soil Sampling
2.2. Water Content and Organic Carbon
2.3. Soil Metabolic Fingerprint
2.4. Catabolic Response Profile (CRP)
2.5. Biolog Community Level Physiological Profile (CLPP)
2.6. Statystical Analysis
3. Results
3.1. Water Content and Soil Organic Carbon
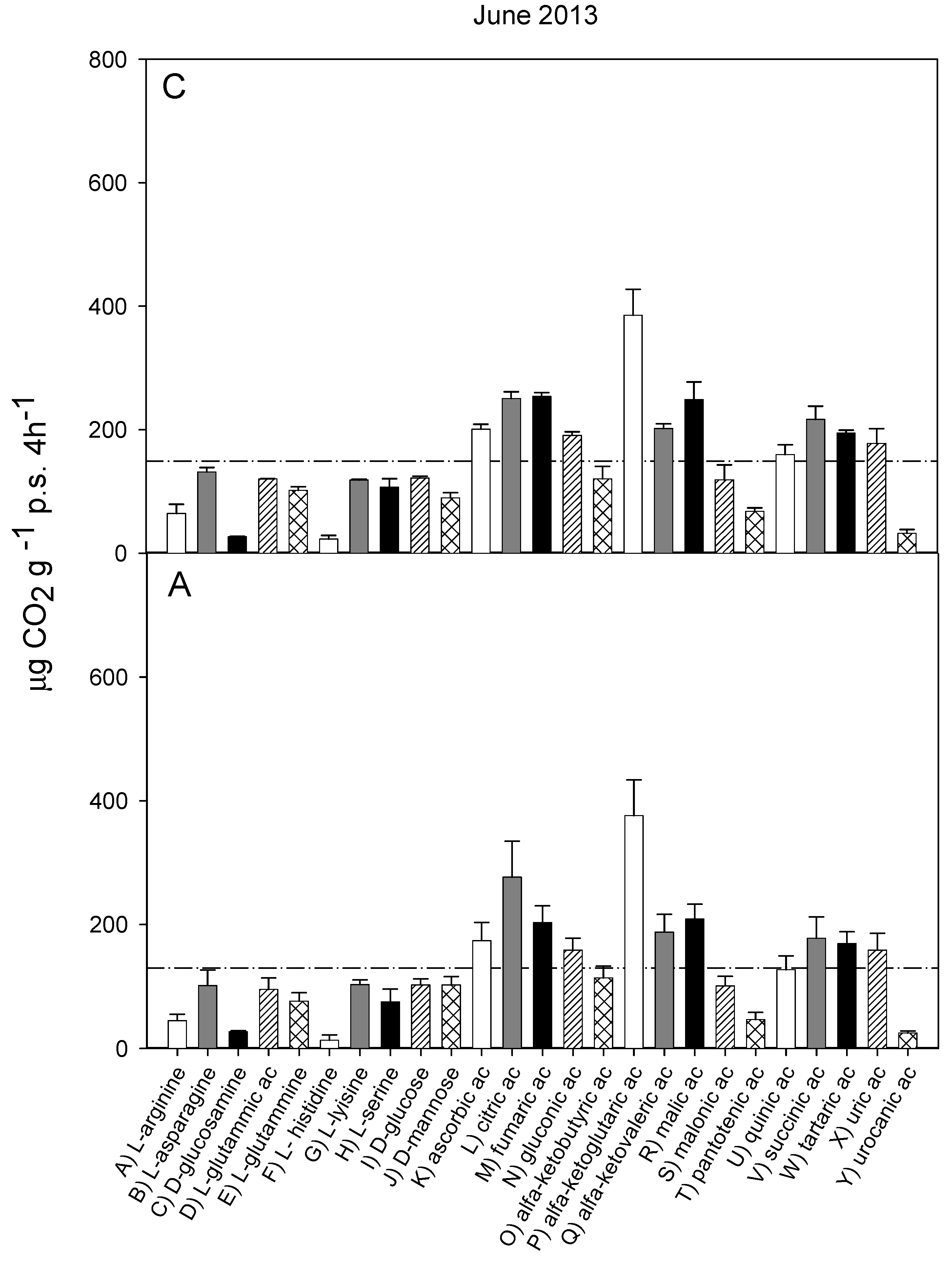
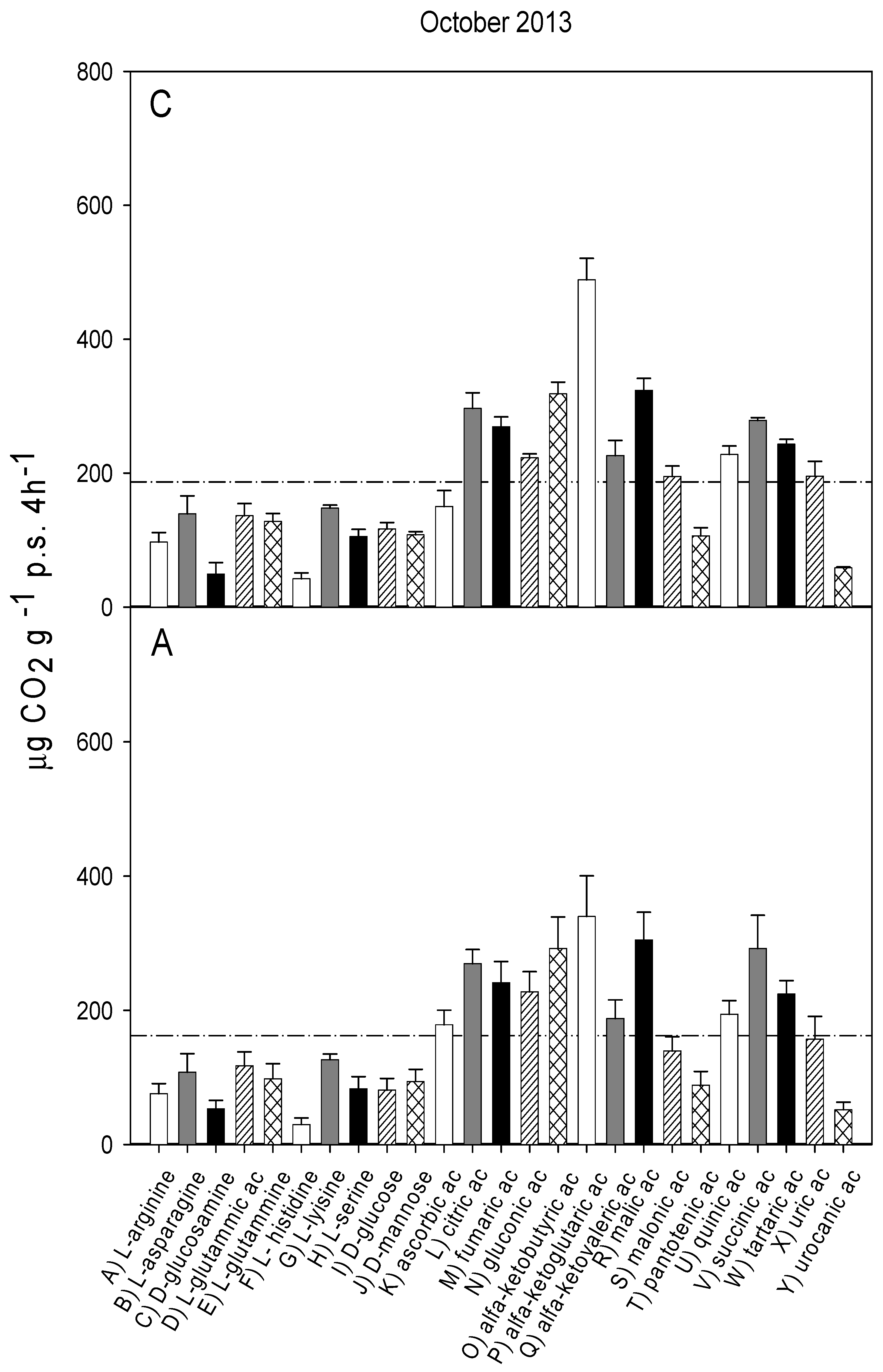
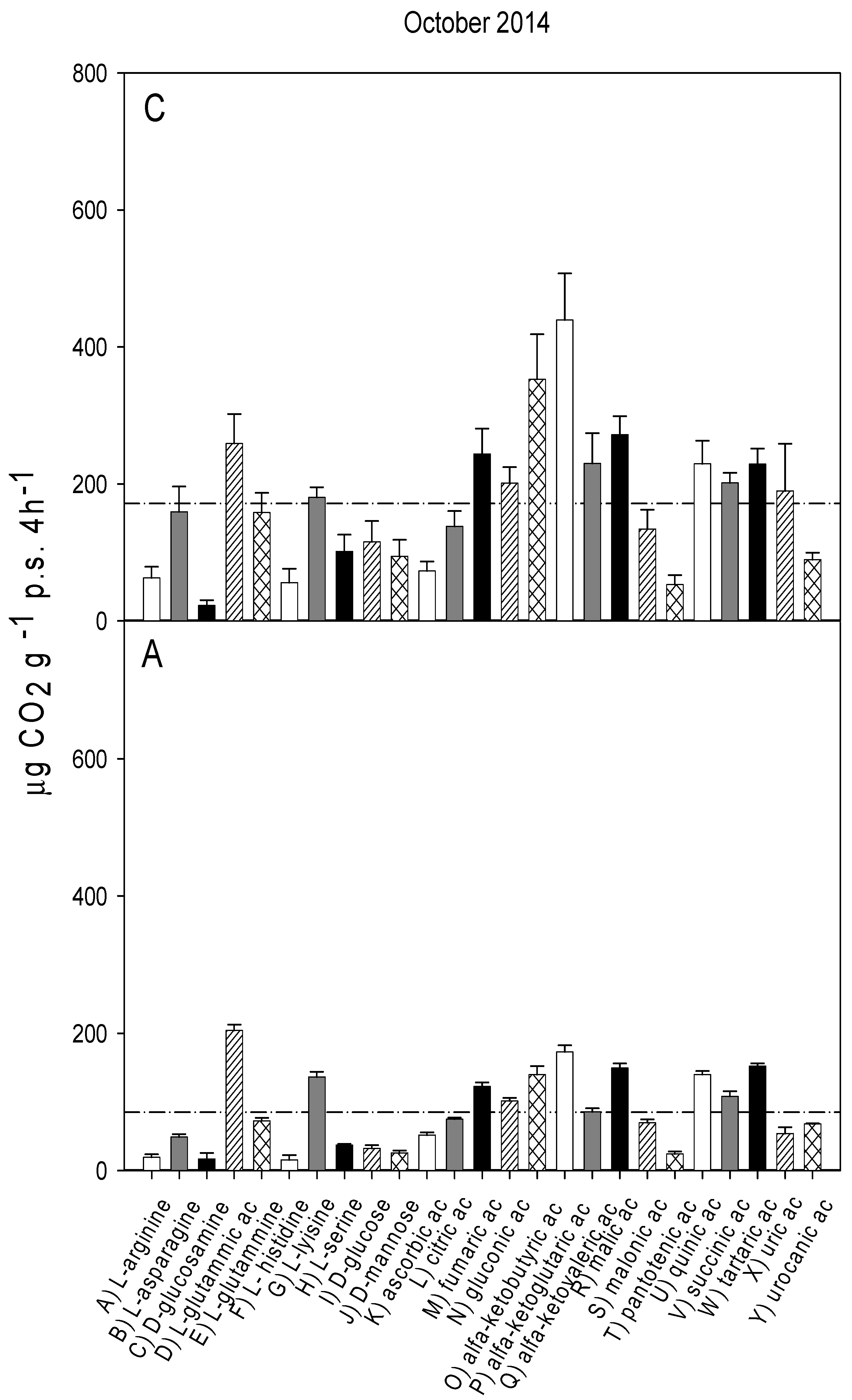
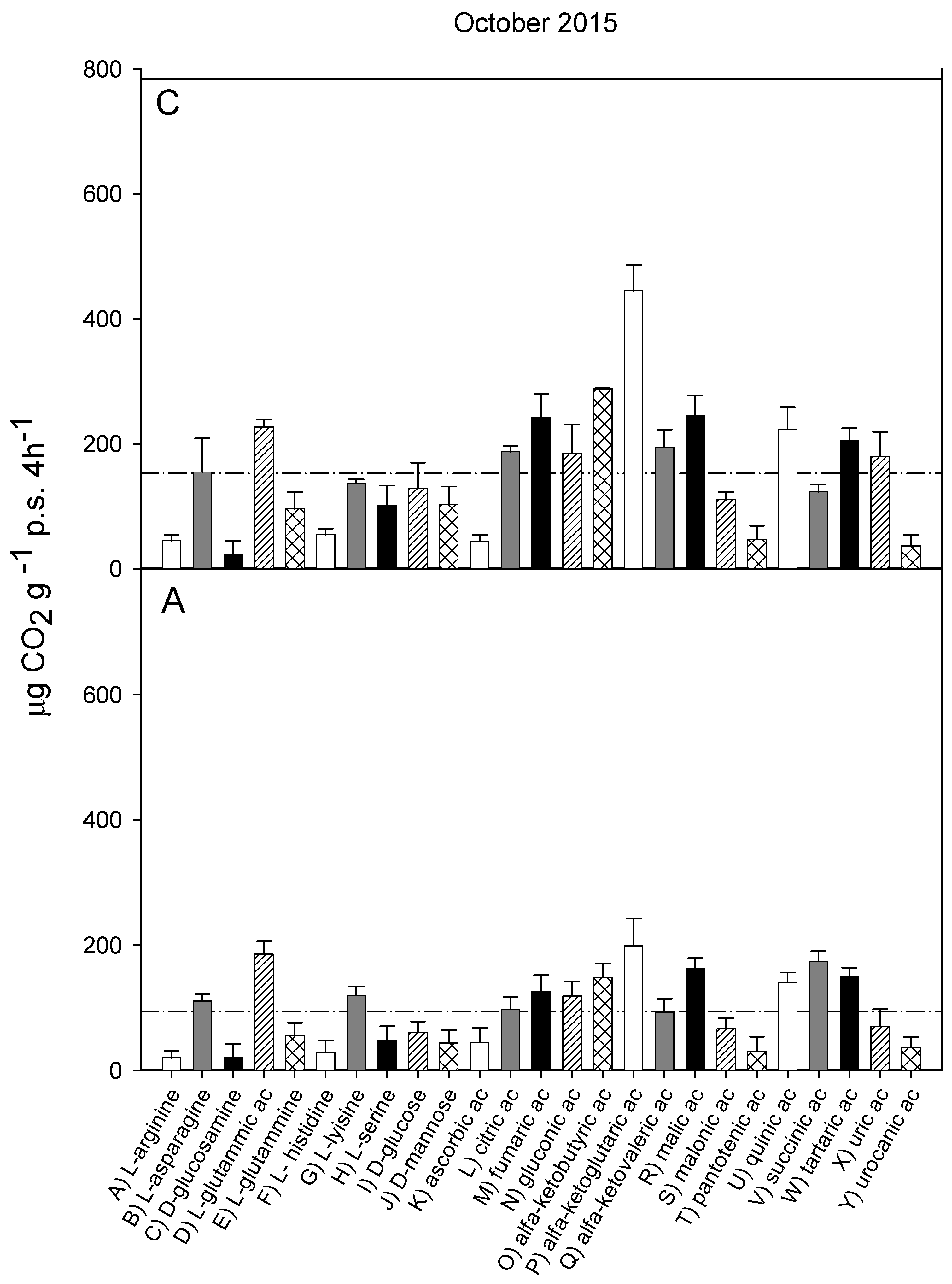
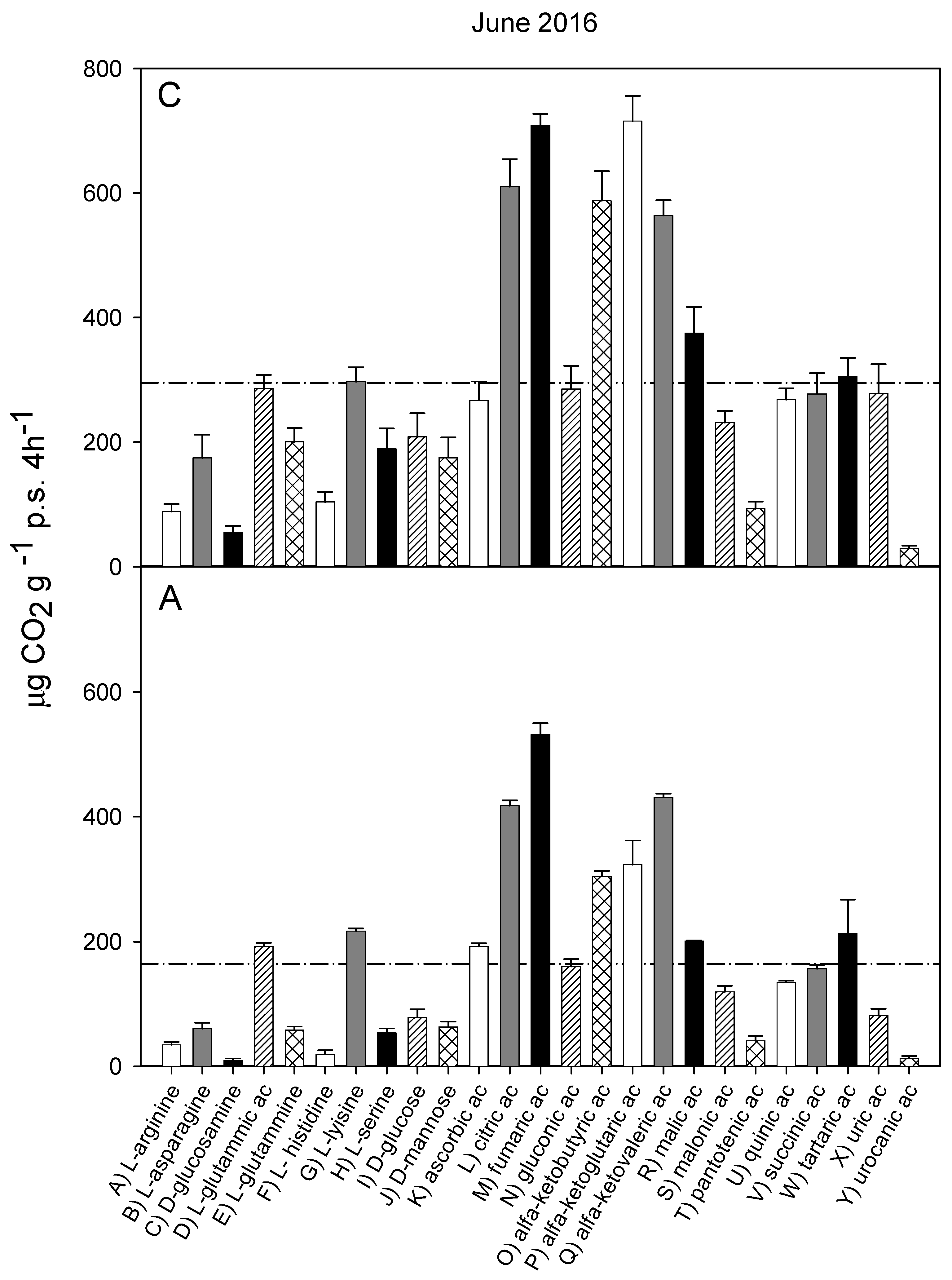
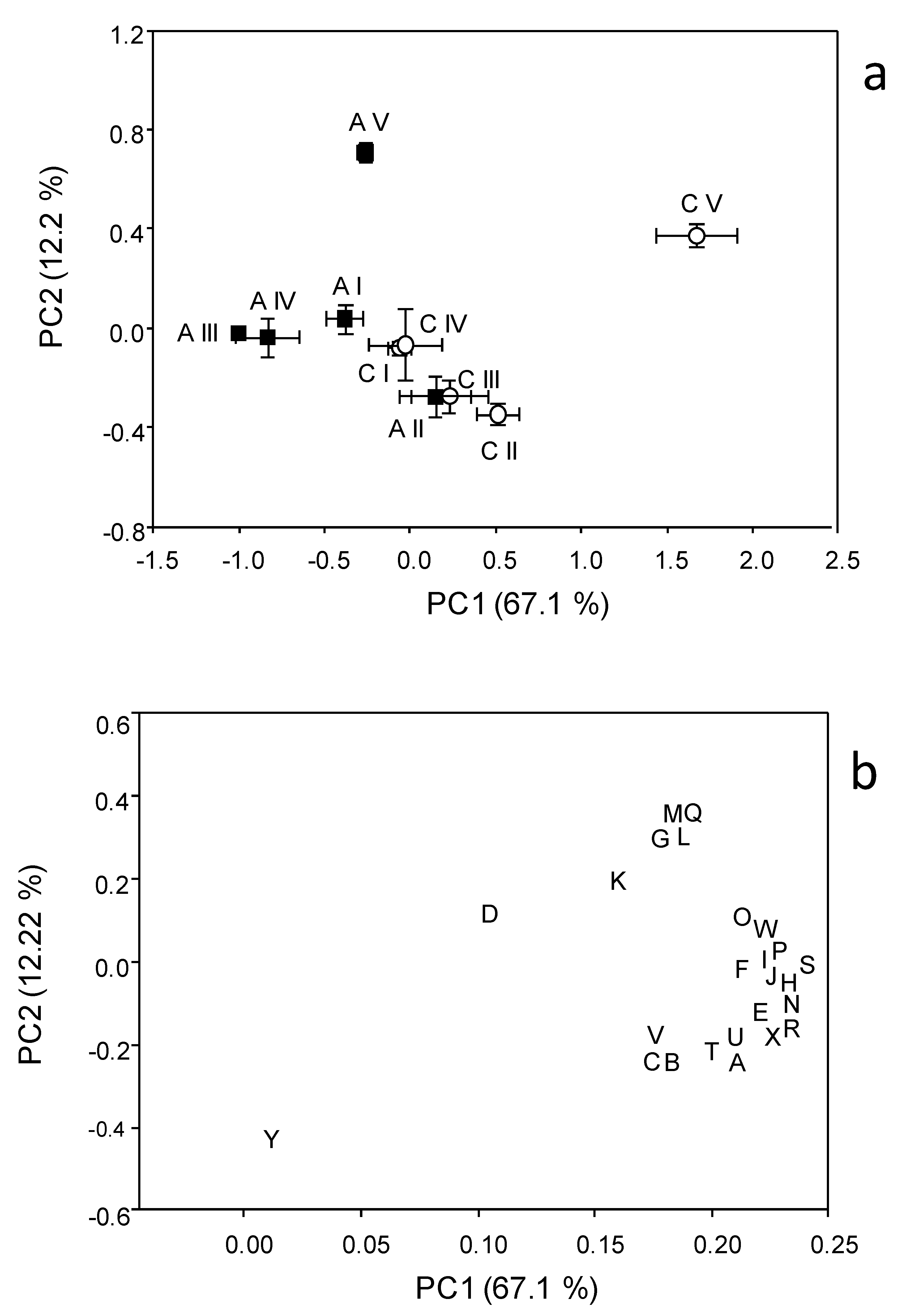
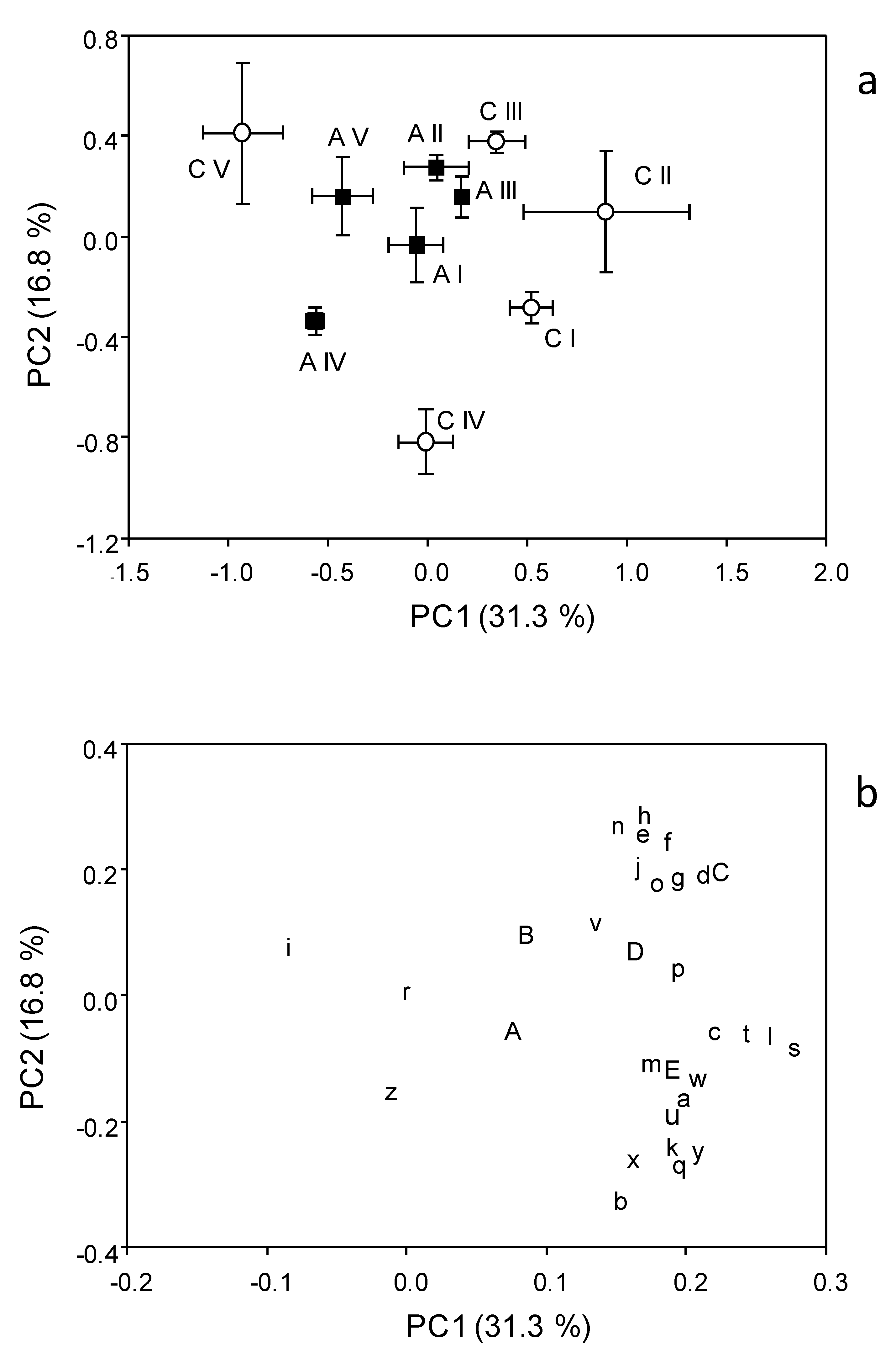
3.2. Catabolic Response Profile
3.3. Biolog Community Level Physiological Profile
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Köhl, M.; Lasco, R.; Cifuentes, M.; Jonsson, Ö.; Korhonen, K.T.; Mundhenk, P.; de Jesus Navar, J.; Stinson, G. Changes in forest production, biomass and carbon: Results from the UN FAO Global Forest Resource Assessment. For. Ecol. Manag. 2015, 352, 21–34. [Google Scholar] [CrossRef] [Green Version]
- Dixon, R.K.; Brown, S.; Houghton, R.A.; Solomon, A.M.; Trexler, M.C.; Wisniewski, J. Carbon pools and flux of global forest ecosystems. Sci. New Ser. 1994, 263, 185–190. [Google Scholar] [CrossRef]
- Schoene, D.; Killmann, W.; von Lüpke, H.; Loyche-Wilkie, M. Definitional Issues Related to Reducing Emissions from Deforestation in Developing Countries; Forests and Climate Change Working Paper. No. 5; FAO: Rome, Italy, 2007. [Google Scholar]
- Bradford, A. Deforestation: Facts, Causes and Effects. Available online: www.livescience.com/27692-deforestation.html (accessed on 15 October 2020).
- Kissinger, G.; Herold, M.; De Sy, V. Drivers of Deforestation and Forest Degradation: A Synthesis Report for REDD+ Policymakers; Lexeme Consulting: Vancouver, BC, Canada, 2012. [Google Scholar]
- FAO. State of the World’s Forests 2016. Forests and Agriculture: Land-Use Challenges and Opportunities. Available online: http://www.fao.org/3/a-i5588e.pdf (accessed on 15 October 2020).
- Chakravarty, S.; Ghosh, S.K.; Suresh, C.P.; Dey, A.N.; Shukla, G. Deforestation: Causes, Effects and Control Strategies. In Global Perspectives on Sustainable Forest Management; InTech: Rijeka, Croatia, 2012; Available online: http://www.intechopen.com/books/globalperspectives-on-sustainable-forest-management/deforestation-causes-effects-and-control-strategies (accessed on 15 October 2020).
- Gaertner, M.A.; Christensen, O.B.; Prego, J.A.; Polcher, J.; Gallardo, C.; Castro, M. The impact of deforestation on the hydrological cycle in the western Mediterranean: An ensemble study with two regional climate models. Clim. Dyn. 2001, 17, 857–873. [Google Scholar] [CrossRef]
- Mongabay. Deforestation Statistics for Italy. Available online: rainforests.mongabay.com (accessed on 15 October 2020).
- Hajabbasi, M.A.; Jalalian, A.; Karimzadeh, H.R. Deforestation effects on soil physical and chemical properties, Lordegan, Iran. Plant Soil 1997, 190, 301–308. [Google Scholar] [CrossRef]
- Attiwill, P.M.; Adams, M.A. Tansley review No. 50. Nutrient cycling in forests. New Phytol. 1993, 124, 561–582. [Google Scholar] [CrossRef]
- Kiikkilä, O.; Kitunen, V.; Smolander, A. Dissolved soil organic matter from surface organic horizons under birch and conifers: Degradation in relation to chemical characteristics. Soil Biol. Biochem. 2006, 38, 737–746. [Google Scholar] [CrossRef]
- Degens, B.P.; Harris, J.A. Development of a physiological approach to measuring the catabolic diversity of soil microbial communities. Soil Biol. Biochem. 1997, 29, 1309–1320. [Google Scholar] [CrossRef]
- Stefanowicz, A. The Biolog plates technique as a tool in ecological studies of microbial communities. Pol. J. Environ. Stud. 2006, 15, 669–676. [Google Scholar]
- Graham, M.H.; Haynes, R.J. Catabolic diversity of soil communities under sugarcane and other land uses estimated by Biolog and substrate-induced respiration methods. Appl. Soil Ecol. 2005, 29, 155–164. [Google Scholar] [CrossRef]
- Scotti, R.; Iovieno, P.; Zaccardelli, M. Comparative assessment of enzyme activities and soil microbial communities in paired chestnut forest and agricultural soils. Biol. Fertil. Soils 2015, 51, 1013–1019. [Google Scholar] [CrossRef]
- Walkley, A.J.; Black, I.A. Estimation of soil organic carbon by the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Garland, J.L.; Mills, A.L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Appl. Environ. Microbiol. 1991, 57, 2351–2359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degens, B.P.; Schipper, L.A.; Sparling, G.P.; Vojvodic-Vukovic, M. Decreases in organic C reserves in soils can reduce the catabolic diversity of soil microbial communities. Soil Biol. Biochem. 2000, 32, 189–196. [Google Scholar] [CrossRef]
- Bartelt-Ryser, J.; Joshi, J.; Schmid, B.; Brandl, H.; Balser, T. Soil feedbacks of plant-diversity on soil microbial communities and subsequent plant growth. Perspect. Plant Ecol. 2005, 7, 27–49. [Google Scholar] [CrossRef]
- Yao, H.; He, Z.; Wilson, M.J.; Campbell, C.D. Microbial Biomass and Community Structure in a Sequence of Soils with Increasing Fertility and Changing Land Use. Microb. Ecol. 2000, 40, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Fulthorpe, R.R.; Allen, D.G. Evaluation of biolog MT plates for aromatic and chloroaromatic substrate utilization tests. Can. J. Microbiol. 1994, 40, 1067–1071. [Google Scholar] [CrossRef]
- Staddon, W.J.; Duchensne, L.C.; Trevors, J.T. Microbial diversity and community structure of post-disturbance forest soils as determined by sole carbon-source utilization patterns. Microb. Ecol. 1997, 34, 125–130. [Google Scholar] [CrossRef]
- Guo, Y.J.; Ni, Y.; Han, J.G. The influence of land use change on chemical and biological properties of steppe soils in northern China. Arid Land Res. Manag. 2009, 23, 197–212. [Google Scholar] [CrossRef]
- Katsalirou, E.; Deng, S.; Nofziger, D.L.; Gerakis, A. Long-term management effects on organic C and N pools and activities of C-transforming enzymes in prairie soils. Eur. J. Soil Biol. 2010, 46, 335–341. [Google Scholar] [CrossRef]
- Qiu, L.; Wei, X.; Zhang, X.; Cheng, J.; Gale, W.; Guo, C.; Long, T. Soil organic carbon losses due to land use change in a semiarid grassland. Plant Soil 2012, 355, 299–309. [Google Scholar] [CrossRef]
- Ding, F.; Hu, Y.L.; Li, L.J.; Li, A.; Shi, S.; Lian, P.Y.; Zeng, D.H. Changes in soil organic carbon and total nitrogen stocks after conversion of meadow to cropland in Northeast China. Plant Soil 2013, 373, 659–672. [Google Scholar] [CrossRef]
- Lal, R. Land use and soil management effects on soil organic matter dynamics on Alfisols in Western Nigeria. In Soil Processes and the Carbon Cycle; CRC Press LLC: Boca Raton, FL, USA, 1997; pp. 109–126. [Google Scholar]
- Haque, S.M.S.; Das Gupta, S.; Miah, S. Deforestation effects on biological and other important soil properties in an upland watershed of Bangladesh. J. For. Res. 2014, 25, 877–885. [Google Scholar] [CrossRef]
- Bissett, A.; Richardson, A.E.; Baker, G.; Peter, H.; Thrall, P.H. Long-term land use effects on soil microbial community structure and function. Appl. Soil Ecol. 2011, 5, 66–78. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2017, 68, 12–26. [Google Scholar] [CrossRef]
- Chaer, G.; Fernandes, M.; Myrold, D.; Bottomley, P. Comparative resistance and resilience of soil microbial communities and enzyme activities in adjacent native forest and agricultural soils. Microb. Ecol. 2009, 58, 414–424. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| June 2013 (I) | October 2013 (II) | October 2014 (III) | October 2015 (IV) | June 2016 (V) | ||
|---|---|---|---|---|---|---|
| WC (% d.w.) | C | 23.9 ± 2.9 A, b | 44.2 ± 1.5 A, a | 36.4 ± 2.9 A, b | 37.2 ±6.6 A, a | 38.3 ± 3.8 A, b |
| A | 26.7 ± 2.9 A, b | 41.6 ± 1.1 A, a | 26.0 ±2.6 A, b | 38.8 ± 2.2 A, a | 26.6 ± 1.9 A, b | |
| C org (g kg−1 d.w.) | C | 43,9 ± 2.5 A, a | 54.7 ± 2.8 A, a | 49.2 ± 6.3 A, a | 47.8 ± 9.7 A, a | 53.7 ± 5.5 A, a |
| A | 32.7 ± 1.7 B, a | 30.9 ± 0.9 A, a | 31.1 ± 0.4 A, a | 34.6 ± 1.7 A, a | 29.4 ± 0.1 B, a | |
| CE | C | 18.5 ± 0.68 A, a | 19.2 ± 0.42 A, a | 18.8 ± 0.35 A, a | 17.7 ± 0.92 A, a | 18.1 ± 0.29 A, a |
| A | 13.6 ± 0.45 B, bc | 16.4 ± 0.48 A, ad | 17.8 ± 0.19 A, a | 18.3 ± 0.99 A, a | 14.5 ± 0.27 B, bd | |
| H’ | C | 3.10 ± 0.03 A, ab | 3.19 ± 0.05 A, ab | 3.20 ± 0.01 A, a | 2.80 ± 0.01 A, ab | 2.75 ± 0.1 A, b |
| A | 3.03 ± 0.07 A, ab | 2.99 ± 0.11 B, ab | 3.06 ± 0.04 A, a | 2.77 ± 0.02 A, b | 2.99 ± 0.09 B, ab | |
| CRP (n = 25) | rs | 0.908 *** | 0.986 *** | 0.906 *** | 0.879 *** | 0.967 *** |
| CLPP (n = 31) | rs | 0.856 *** | 0.823 *** | 0.759 *** | 0.899 *** | 0.804 *** |
| Sampling | I | II | III | IV | V | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Substrate | A | C | A | C | A | C | A | C | A | C |
| (a) Pyruvic acid methyl ester | 0.99 | 1.47 | 1.12 | 1.51 | 1.06 | 1.53 | 1.32 | 1.60 | 0.63 | 0.32 |
| (b) Tween 40 | 1.58 | 1.77 | 1.29 | 1.51 | 1.23 | 1.22 | 1.53 | 1.66 | 1.34 | 0.95 |
| (c) Tween 80 | 1.47 | 1.61 | 1.33 | 1.45 | 1.25 | 0.99 | 0.98 | 1.03 | 0.93 | 0.76 |
| (d) α-Cyclodextrine | 0.42 | 0.29 | 0.29 | 0.59 | 0.23 | 0.57 | 0.01 | 0.00 | 0.34 | 0.07 |
| (e) Glycogen | 0.15 | 0.20 | 0.50 | 1.12 | 0.61 | 1.32 | 0.22 | 0.04 | 0.30 | 0.35 |
| (f) D-Cellobiose | 1.07 | 0.95 | 1.23 | 1.65 | 0.88 | 1.38 | 0.31 | 0.25 | 1.13 | 0.50 |
| (g) D-Lactose | 0.57 | 0.97 | 0.89 | 1.38 | 0.24 | 1.02 | 0.06 | 0.05 | 0.47 | 0.41 |
| (h) b-Methyl-D-glucoside | 0.82 | 0.95 | 1.40 | 1.00 | 1.31 | 1.20 | 0.26 | 0.01 | 0.65 | 0.48 |
| (i) D-Xylose | 0.06 | 0.12 | 0.15 | 0.12 | 0.04 | 0.00 | 0.08 | 0.09 | 0.05 | 0.33 |
| (j) i-Erythritol | 0.35 | 0.39 | 0.56 | 0.51 | 0.49 | 0.61 | 0.05 | 0.00 | 0.27 | 0.17 |
| (k) D-Mannitol | 1.65 | 1.89 | 1.70 | 2.01 | 1.84 | 1.50 | 1.74 | 2.16 | 1.60 | 0.51 |
| (l) N-Acetil-D-glucosamine | 1.01 | 1.49 | 1.28 | 1.71 | 1.35 | 1.48 | 0.95 | 1.45 | 0.83 | 0.65 |
| (m) D-Glucosaminic acid | 1.07 | 1.14 | 1.09 | 1.34 | 1.68 | 1.39 | 0.94 | 1.52 | 0.86 | 0.77 |
| (n) Glucose-1-Phosphate | 0.87 | 0.71 | 0.87 | 0.77 | 1.41 | 1.16 | 0.01 | 0.00 | 0.54 | 0.38 |
| (o) D,L-α-Glycerol phosphate | 0.13 | 0.20 | 0.27 | 0.46 | 0.25 | 0.21 | 0.02 | 0.09 | 0.15 | 0.21 |
| (p) D-Galactonic acid γ-lactone | 0.62 | 0.81 | 0.81 | 1.12 | 0.90 | 0.90 | 0.57 | 0.91 | 0.54 | 0.78 |
| (q) D-Galacturonic acid | 1.41 | 1.90 | 1.22 | 1.77 | 1.62 | 1.45 | 1.46 | 2.01 | 1.49 | 0.86 |
| (r) 2-Hidroxy benzoic acid | 0.00 | 0.02 | 0.07 | 0.01 | 0.03 | 0.00 | 0.03 | 0.01 | 0.02 | 0.00 |
| (s) 4-Hydroxy benzoic acid | 0.87 | 1.31 | 0.93 | 1.45 | 0.76 | 0.92 | 0.77 | 0.85 | 0.72 | 0.28 |
| (t) γ-Hydroxybutyric acid | 0.41 | 0.72 | 0.25 | 0.63 | 0.25 | 0.18 | 0.10 | 0.25 | 0.14 | 0.03 |
| (u) Itaconic Acid | 0.43 | 1.31 | 0.23 | 1.45 | 0.16 | 1.08 | 0.22 | 1.41 | 0.43 | 0.22 |
| (v) γ-Ketobutyric acid | 0.22 | 0.10 | 0.07 | 0.49 | 0.40 | 0.22 | 0.14 | 0.02 | 0.27 | 0.02 |
| (w) D-Malic acid | 0.68 | 2.01 | 0.65 | 1.66 | 0.89 | 1.71 | 0.88 | 1.42 | 0.58 | 0.69 |
| (x) L-Arginine | 1.22 | 1.36 | 1.02 | 1.86 | 0.98 | 1.15 | 1.10 | 1.87 | 0.86 | 0.88 |
| (y) L-Asparagine | 1.78 | 1.95 | 1.94 | 1.88 | 1.96 | 1.68 | 1.61 | 2.22 | 1.48 | 1.03 |
| (z) L-Phenylalanine | 0.47 | 0.68 | 0.29 | 0.72 | 0.58 | 0.44 | 1.02 | 0.30 | 0.58 | 0.39 |
| (A) L-Serine | 1.05 | 1.44 | 3.08 | 1.53 | 1.69 | 1.28 | 0.96 | 1.93 | 1.13 | 0.63 |
| (B) L-Threonine | 0.23 | 0.32 | 0.18 | 0.47 | 0.36 | 0.65 | 0.42 | 0.20 | 0.37 | 0.33 |
| (C) Glycil-L-glutamic acid | 0.28 | 0.51 | 0.58 | 0.62 | 0.51 | 0.39 | 0.13 | 0.08 | 0.33 | 0.16 |
| (D) Phenyletyl-amine | 0.11 | 0.42 | 0.09 | 0.44 | 0.12 | 0.44 | 0.09 | 0.17 | 0.12 | 0.29 |
| (E) Putrescine | 1.02 | 0.90 | 0.73 | 0.95 | 1.32 | 0.73 | 0.46 | 1.03 | 0.47 | 0.52 |
| AWCD | 0.74 ± 0.07 A,a | 0.97 ± 0.1 B,ab | 0.84 ± 0.05 A,a | 1.10 ± 0.02 B,a | 0.85 ± 0.01 A,a | 0.93 ± 0.09 A,ad | 0.60 ± 0.01 A,a | 0.80 ± 0.01 B,bd | 0.63 ± 0.09 A,a | 0.45 ± 0.12 A,c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iovieno, P.; Scotti, R.; Zaccardelli, M. Functional Diversity of Soil Microbial Community after Conversion of a Chestnut Forest to an Agricultural System. Agriculture 2021, 11, 43. https://doi.org/10.3390/agriculture11010043
Iovieno P, Scotti R, Zaccardelli M. Functional Diversity of Soil Microbial Community after Conversion of a Chestnut Forest to an Agricultural System. Agriculture. 2021; 11(1):43. https://doi.org/10.3390/agriculture11010043
Chicago/Turabian StyleIovieno, Paola, Riccardo Scotti, and Massimo Zaccardelli. 2021. "Functional Diversity of Soil Microbial Community after Conversion of a Chestnut Forest to an Agricultural System" Agriculture 11, no. 1: 43. https://doi.org/10.3390/agriculture11010043
APA StyleIovieno, P., Scotti, R., & Zaccardelli, M. (2021). Functional Diversity of Soil Microbial Community after Conversion of a Chestnut Forest to an Agricultural System. Agriculture, 11(1), 43. https://doi.org/10.3390/agriculture11010043