
Rhizosphere Microbiome Regulates the Growth of Mustard under Organic Greenhouse Cultivation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Place of Research and the Collection of Samples for Microbiota Profiling
2.2. PCR Amplification, Sequencing, and Statistical Analysis
2.3. Soil Physicochemical Properties
3. Results
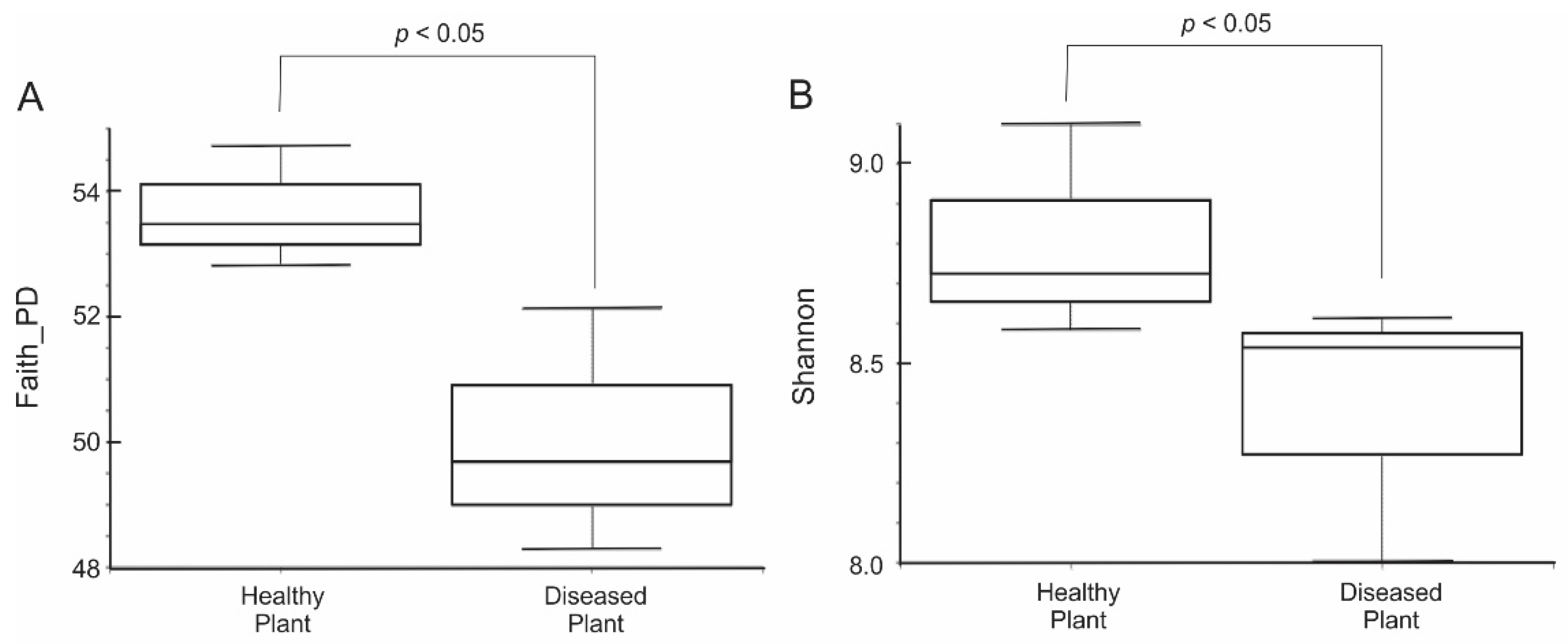
3.1. Microbial Biodiversity within Rhizosphere Soil Communities
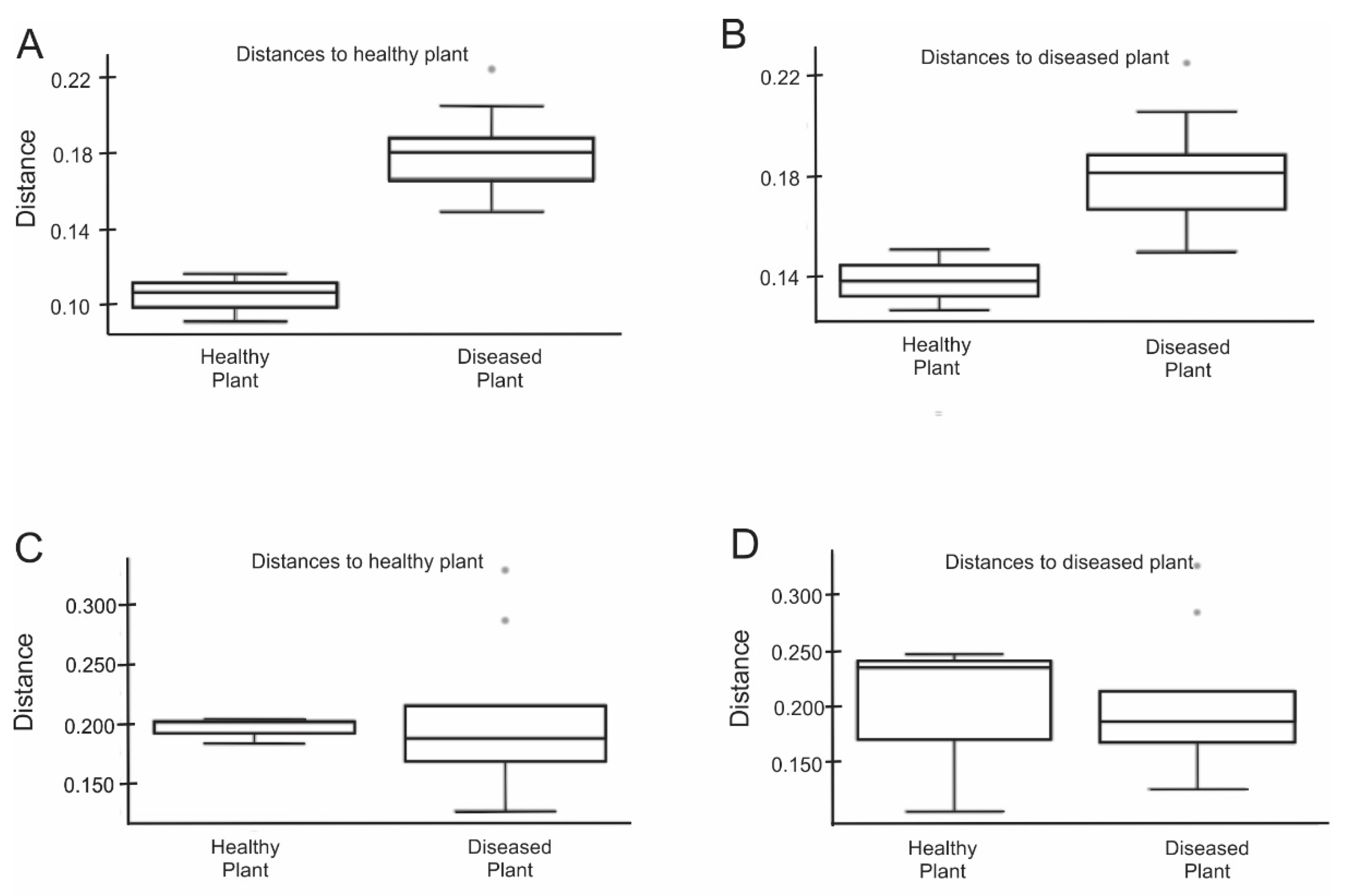
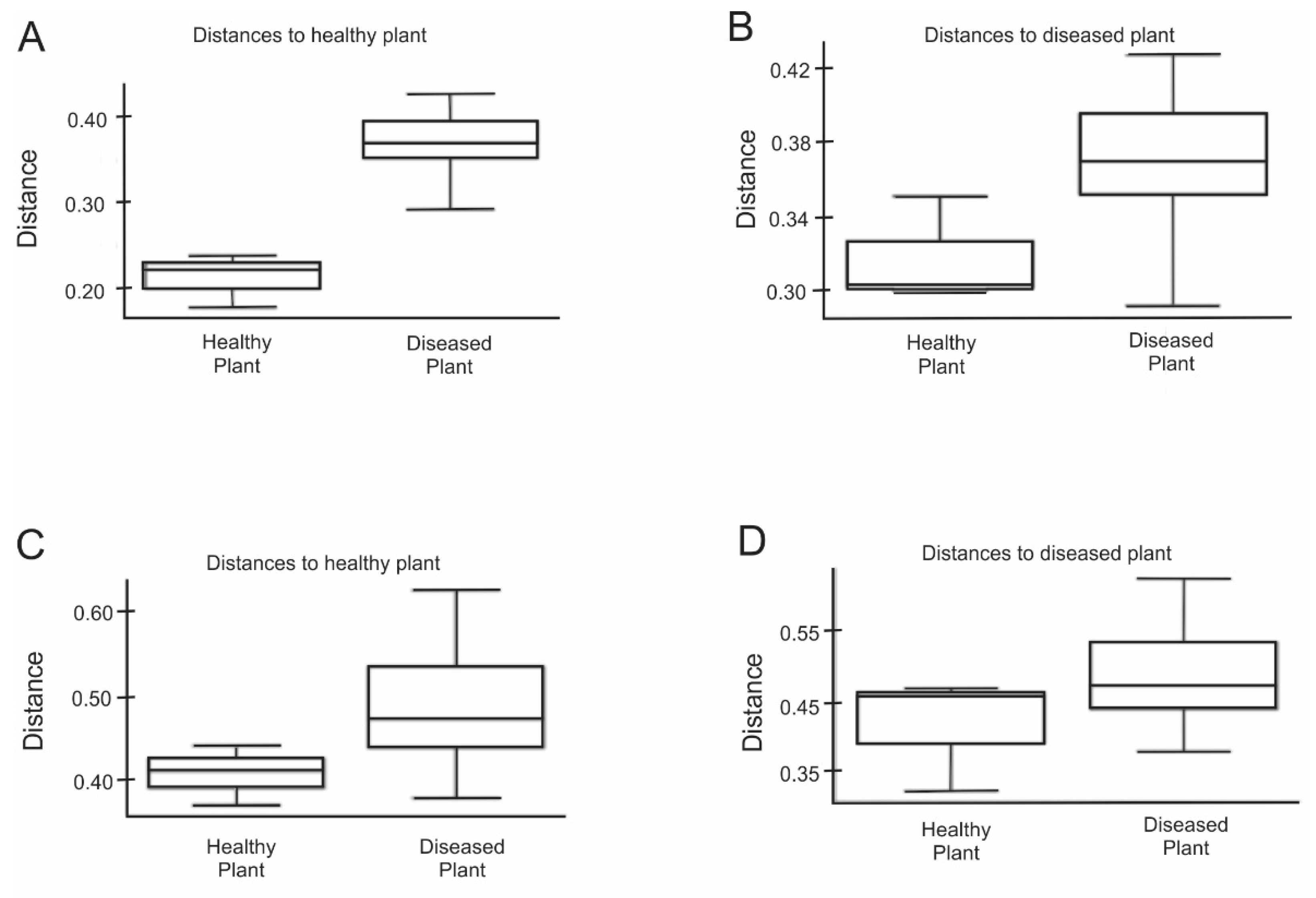
3.2. The Rhizosphere MMicrobiomes of Healthy and Diseased Mustard Plant Communities
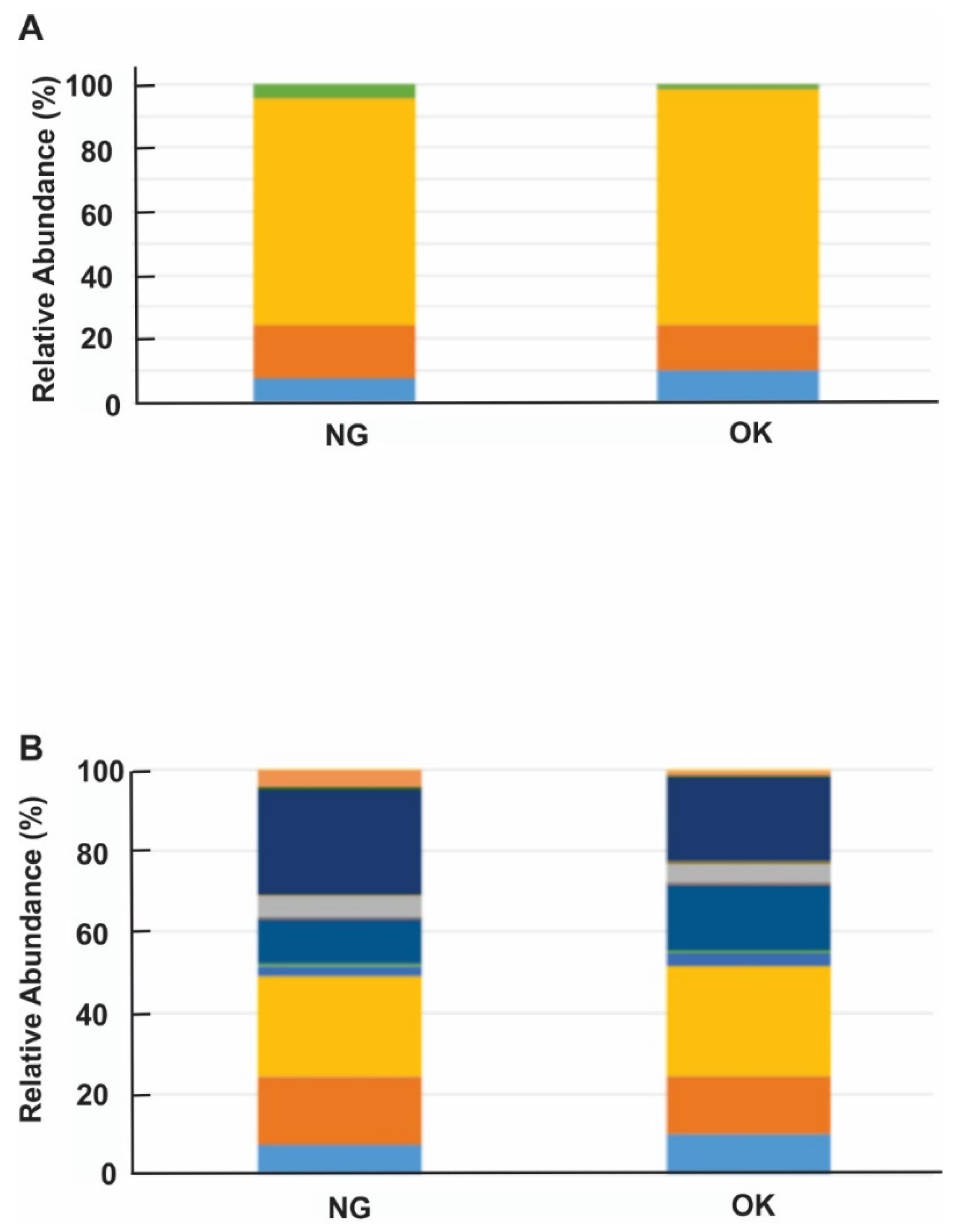
3.3. The Composition of Bacterial Taxa between the Healthy and Diseased Rhizosphere Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van der Heijden, M.G.A.; Bardgett, R.D.; Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Walters, K.E.; Martiny, J.B. Alpha-, beta-, and gamma-diversity of bacteria varies across habitats. PLoS ONE 2020, 15, e0233872. [Google Scholar] [CrossRef]
- Hartmann, M.; Frey, B.; Mayer, J.; Mäder, P.; Widmer, F. Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 2015, 9, 1177–1194. [Google Scholar] [CrossRef] [Green Version]
- Lupatini, M.; Korthals, G.W.; de Hollander, M.; Janssens, T.K.S.; Kuramae, E.E. Soil microbiome is more heterogeneous in organic than in conventional farming system. Front. Microbiol. 2017, 7, 2064. [Google Scholar] [CrossRef] [Green Version]
- Armalytë, J.; Skerniškytë, J.; Bakienë, E.; Krasauskas, R.; Šiugždinienë, R.; Kareivienë, V.; Kerzienė, S.; Klimienė, I.; Sužiedėlienė, E.; Ružauskas, M. Microbial diversity and antimicrobial resistance profile in microbiota from soils of conventional and organic farming systems. Front. Microbiol. 2019, 10, 892. [Google Scholar] [CrossRef]
- Hartman, K.; Tringe, S.G. Interactions between plants and soil shaping the root microbiome under abiotic stress. Biochem. J. 2019, 476, 2705–2724. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.C.; Yu, S.W. Occurrence and Management of Diseases and Pests of Cruciferous Vegetables (in Chinese); Taiwan Agricultural Chemicals and Toxic Substances Research Institute: Taichung, Taiwan, 2012; pp. 14–36.
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [Green Version]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Cheng, Z.; Meng, H.; Ahmad, I.; Zhao, H. Growth, yield and quality of spring tomato and physicochemical properties of medium in a tomato/garlic intercropping system under plastic tunnel organic medium cultivation. Sci. Hortic. 2014, 170, 159–168. [Google Scholar] [CrossRef]
- Kim, J.M.; Roh, A.S.; Choi, S.C.; Kim, E.J.; Choi, M.T.; Ahn, B.K.; Kim, S.; Lee, Y.; Joa, J.; Kang, S.; et al. Soil pH and electrical conductivity are key edaphic factors shaping bacterial communities of greenhouse soils in Korea. J. Microbiol. 2016, 54, 838–845. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Girard, R.A.; Martin, T.; Edwards, J.; Thrash, J.C. Status of the archaeal and bacterial census: An update. mBio 2016, 7, e00201–e00216. [Google Scholar] [CrossRef] [Green Version]
- He, Z.L.; Baligar, V.; Ritchey, K.; Martens, D. Determination of soluble phosphorus in the presence of organic ligands or fluoride. Soil Sci. Soc. Am. J. 1998, 62, 1538–1541. [Google Scholar] [CrossRef]
- Wei, L.L.; Chen, C.R.; Xu, Z.H. The effect of low-molecular-weight organic acids and inorganic phosphorus concentration on the determination of soil phosphorus by the molybdenum blue reaction. Biol. Fertil. Soils 2009, 45, 775–779. [Google Scholar] [CrossRef]
- Machado, R.M.A.; Serralheiro, R.P. Soil salinity: Effect on vegetable crop growth. Management practices to prevent and mitigate soil salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- Rawat, S.R.; Männistö, M.K.; Bromberg, Y.; Häggblom, M.M. Comparative genomic and physiological analysis provides insights into the role of Acidobacteria in organic carbon utilization in Arctic tundra soils. FEMS Microbiol. Ecol. 2012, 82, 341–355. [Google Scholar] [CrossRef] [Green Version]
- Ward, N.L.; Challacombe, J.F.; Janssen, P.H.; Henrissat, B.; Coutinho, P.M.; Wu, M.; Xie, G.; Haft, D.H.; Sait, M.; Badger, J.; et al. Three genomes from the phylum Acidobacteria provide insight into the lifestyles of these microorganisms in soils. Appl. Environ. Microbiol. 2009, 75, 2046–2056. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Gu, C.; Liu, X.; Yang, C.; Li, W.; Wang, S. Impact of soybean nodulation phenotypes and nitrogen fertilizer levels on the rhizosphere bacterial community. Front. Microbiol. 2020, 11, 750. [Google Scholar] [CrossRef] [PubMed]
- Palaniappan, P.; Chauhan, P.S.; Saravanan, V.S.; Anandham, R.; Sa, T. Isolation and characterization of plant growth promoting endophytic bacterial isolates from root nodule of Lespedeza sp. Biol. Fertil. Soils 2010, 46, 807–816. [Google Scholar] [CrossRef]
- Tapia-García, E.Y.; Hernández-Trejo, V.; Guevara-Luna, J.; Rojas-Rojas, F.U.; Arroyo-Herrera, I.; Meza-Radilla, G.; Vásquez-Murrieta, M.S.; Los Santos, P.E. Plant growth-promoting bacteria isolated from wild legume nodules and nodules of Phaseolus vulgaris L. trap plants in central and southern Mexico. Microbiol. Res. 2020, 239, 126522. [Google Scholar] [CrossRef] [PubMed]
- Dar, D.; Thomashow, L.S.; Weller, D.M.; Newman, D.K. Global landscape of phenazine biosynthesis and biodegradation reveals species-specific colonization patterns in agricultural soils and crop microbiomes. eLife 2020, 9, e59726. [Google Scholar] [CrossRef] [PubMed]
- Nanjundappa, A.; Bagyaraj, D.J.; Saxena, A.K.; Kumar, M.; Chakdar, H. Interaction between arbuscular mycorrhizal fungi and Bacillus spp. in soil enhancing growth of crop plants. Fungal Biol. Biotechnol. 2019, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Sanahuja, G.; Banakar, R.; Twyman, R.M.; Capella, T.; Christou, P. Bacillus thuringiensis: A century of research, development and commercial applications. Plant Biotechnol. J. 2011, 9, 283–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azizoglu, U. Bacillus thuringiensis as a biofertilizer and biostimulator: A mini-review of the little-known plant growth-promoting properties of Bt. Curr. Microbiol. 2019, 76, 1379–1385. [Google Scholar] [CrossRef] [PubMed]
- Nazari, M.; Smith, D.L. A PGPR-produced bacteriocin for sustainable agriculture: A review of thuricin 17 characteristics and applications. Front. Plant Sci. 2020, 11, 916. [Google Scholar] [CrossRef]
- Saad, M.M.G.; Kandil, M.; Mohammed, Y.M.M. Isolation and Identification of plant growth-promoting bacteria highly effective in suppressing root rot in fava beans. Curr. Microbiol. 2020, 77, 2155–2165. [Google Scholar] [CrossRef]
- Watsuji, T.; Yamada, S.; Yamabe, T.; Watanabe, Y.; Kato, T.; Saito, T.; Ueda, K.; Beppu, T. Identification of indole derivatives as self-growth inhibitors of Symbiobacterium thermophilum, a unique bacterium whose growth depends on coculture with a Bacillus sp. Appl. Environ. Microbiol. 2007, 73, 6159–6165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.J.; Yang, K.H.; Liu, Y.H.; Lin, Y.J.; Chen, C.Y. Suppression of southern corn leaf blight by a plant growth promoting rhizobacterium Bacillus cereus C1L. Ann. Appl. Biol. 2010, 157, 45–53. [Google Scholar] [CrossRef]
- Liu, J.; Wang, S.; Zheng, X.; Jin, N.; Lu, J.; Huang, Y.; Fan, B.; Wang, F. Antimicrobial activity against phytopathogens and inhibitory activity on solanine in potatoes of the endophytic bacteria isolated from potato tubers. Front. Microbiol. 2020, 11, 570926. [Google Scholar] [CrossRef]
- Liu, D.; Li, K.; Hu, J.; Wang, W.; Liu, X.; Gao, Z. Biocontrol and action mechanism of Bacillus amyloliquefaciens and Bacillus subtilis in soybean phytophthora blight. Int. J. Mol. Sci. 2019, 20, 2908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earl, A.M.; Losick, R.; Kolter, R. Ecology and genomics of Bacillus subtilis. Trends Microbiol. 2008, 16, 269–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, S.P.; Hartmann, A.; Gao, X.; Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42–a review. Front. Microbiol. 2015, 6, 780. [Google Scholar] [CrossRef] [Green Version]
- Kulmatiski, A.; Kardol, P. Getting Plant—Soil feedbacks out of the greenhouse: Experimental and conceptual approaches. In Progress in Botany; Lüttge, U., Beyschlag, W., Murata, J., Eds.; Springer: Berlin, Heidelberg, 2008; Volume 69, pp. 449–472. [Google Scholar] [CrossRef]
- Song, C.; Zhu, F.; Carrión, V.J.; Cordovez, V. Beyond plant microbiome composition: Exploiting microbial functions and plant traits via integrated approaches. Front. Bioeng. Biotechnol. 2020, 8, 896. [Google Scholar] [CrossRef]
- Wei, Z.; Gu, Y.; Friman, V.P.; Kowalchuk, G.A.; Xu, Y.; Shen, Q.; Jousset, A. Initial soil microbiome composition and functioning predetermine future plant health. Sci. Adv. 2019, 5, eaaw0759. [Google Scholar] [CrossRef] [Green Version]



 : Verrucomicrobia;
: Verrucomicrobia;  : Proteobacteria;
: Proteobacteria;  : Actinobacteria;
: Actinobacteria;  : SBR1093;
: SBR1093;  : Planctomycetes;
: Planctomycetes;  : unknown. (B) : Actinobacteria_Actinobacteria_Actinomycetales; : Planctomycetes_OM190_agg27; : Proteobacteria;
: unknown. (B) : Actinobacteria_Actinobacteria_Actinomycetales; : Planctomycetes_OM190_agg27; : Proteobacteria;  : Proteobacteria_Deltaproteobacteria;
: Proteobacteria_Deltaproteobacteria;  : Proteobacteria_Deltaproteobacteria_Desulfuromonadales; : Proteobacteria_Deltaproteobacteria_Desulfovibrionales;
: Proteobacteria_Deltaproteobacteria_Desulfuromonadales; : Proteobacteria_Deltaproteobacteria_Desulfovibrionales;  : Proteobacteria_Deltaproteobacteria_NB1-j;
: Proteobacteria_Deltaproteobacteria_NB1-j;  : Proteobacteria_Epsilonproteobacteria_Campylobacteriales;
: Proteobacteria_Epsilonproteobacteria_Campylobacteriales;  : Proteobacteria_Gammaproteobacteria;
: Proteobacteria_Gammaproteobacteria;  : Proteobacteria_Gammaproteobacteria_Chromatiales; : Proteobacteria_Gammaproteobacteria_Pseudomonadales;
: Proteobacteria_Gammaproteobacteria_Chromatiales; : Proteobacteria_Gammaproteobacteria_Pseudomonadales;  : Proteobacteria_Gammaproteobacteria_Thiotrichales;
: Proteobacteria_Gammaproteobacteria_Thiotrichales;  : SBR1093_VHS-BS-50;
: SBR1093_VHS-BS-50;  : Verrucomicrobia_Pedosphaerae_Pedosphaerales;
: Verrucomicrobia_Pedosphaerae_Pedosphaerales;  : Verrucomicrobia_Spartobacteria_Chthoniobacterales; : Unknown. Healthy plant: OK; diseased plant: NG.
: Verrucomicrobia; : Proteobacteria; : Actinobacteria; : SBR1093; : Planctomycetes; : unknown. (B) : Actinobacteria_Actinobacteria_Actinomycetales; : Planctomycetes_OM190_agg27; : Proteobacteria; : Proteobacteria_Deltaproteobacteria; : Proteobacteria_Deltaproteobacteria_Desulfuromonadales; : Proteobacteria_Deltaproteobacteria_Desulfovibrionales; : Proteobacteria_Deltaproteobacteria_NB1-j; : Proteobacteria_Epsilonproteobacteria_Campylobacteriales; : Proteobacteria_Gammaproteobacteria; : Proteobacteria_Gammaproteobacteria_Chromatiales; : Proteobacteria_Gammaproteobacteria_Pseudomonadales; : Proteobacteria_Gammaproteobacteria_Thiotrichales; : SBR1093_VHS-BS-50; : Verrucomicrobia_Pedosphaerae_Pedosphaerales; : Verrucomicrobia_Spartobacteria_Chthoniobacterales; : Unknown. Healthy plant: OK; diseased plant: NG.
: Verrucomicrobia_Spartobacteria_Chthoniobacterales; : Unknown. Healthy plant: OK; diseased plant: NG.
: Verrucomicrobia; : Proteobacteria; : Actinobacteria; : SBR1093; : Planctomycetes; : unknown. (B) : Actinobacteria_Actinobacteria_Actinomycetales; : Planctomycetes_OM190_agg27; : Proteobacteria; : Proteobacteria_Deltaproteobacteria; : Proteobacteria_Deltaproteobacteria_Desulfuromonadales; : Proteobacteria_Deltaproteobacteria_Desulfovibrionales; : Proteobacteria_Deltaproteobacteria_NB1-j; : Proteobacteria_Epsilonproteobacteria_Campylobacteriales; : Proteobacteria_Gammaproteobacteria; : Proteobacteria_Gammaproteobacteria_Chromatiales; : Proteobacteria_Gammaproteobacteria_Pseudomonadales; : Proteobacteria_Gammaproteobacteria_Thiotrichales; : SBR1093_VHS-BS-50; : Verrucomicrobia_Pedosphaerae_Pedosphaerales; : Verrucomicrobia_Spartobacteria_Chthoniobacterales; : Unknown. Healthy plant: OK; diseased plant: NG.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, T.-C.; Peng, H.-M.; Wollney, S.; Shen, C.-H. Rhizosphere Microbiome Regulates the Growth of Mustard under Organic Greenhouse Cultivation. Agriculture 2021, 11, 987. https://doi.org/10.3390/agriculture11100987
Liu T-C, Peng H-M, Wollney S, Shen C-H. Rhizosphere Microbiome Regulates the Growth of Mustard under Organic Greenhouse Cultivation. Agriculture. 2021; 11(10):987. https://doi.org/10.3390/agriculture11100987
Chicago/Turabian StyleLiu, Ting-Chung, Hui-Mei Peng, Seth Wollney, and Chang-Hui Shen. 2021. "Rhizosphere Microbiome Regulates the Growth of Mustard under Organic Greenhouse Cultivation" Agriculture 11, no. 10: 987. https://doi.org/10.3390/agriculture11100987
APA StyleLiu, T.-C., Peng, H.-M., Wollney, S., & Shen, C.-H. (2021). Rhizosphere Microbiome Regulates the Growth of Mustard under Organic Greenhouse Cultivation. Agriculture, 11(10), 987. https://doi.org/10.3390/agriculture11100987