
Application of Novel Microorganism-Based Formulations as Alternative to the Use of Iron Chelates in Strawberry Cultivation

 ,
, 
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Soil Characterization
2.3. Treatments
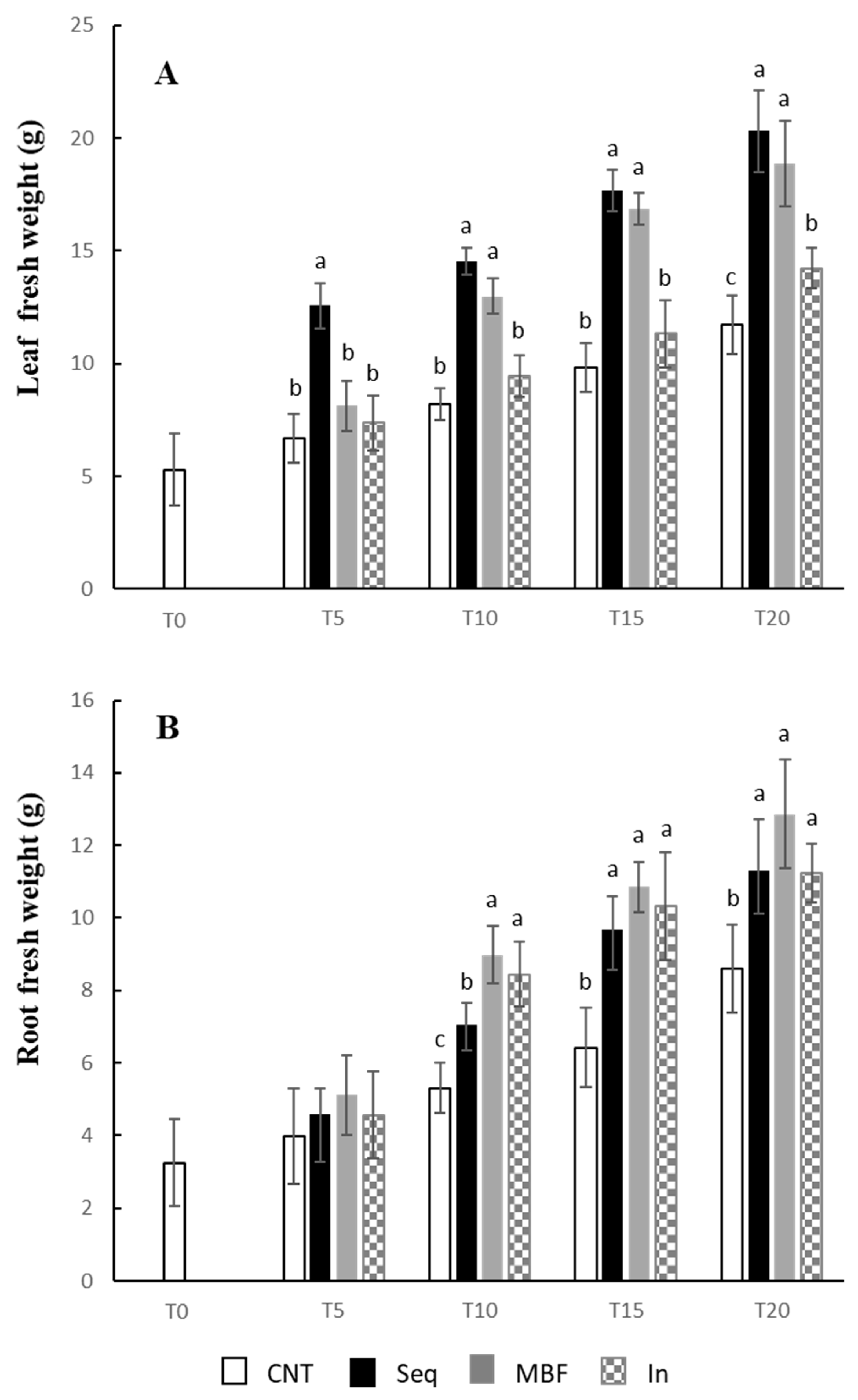
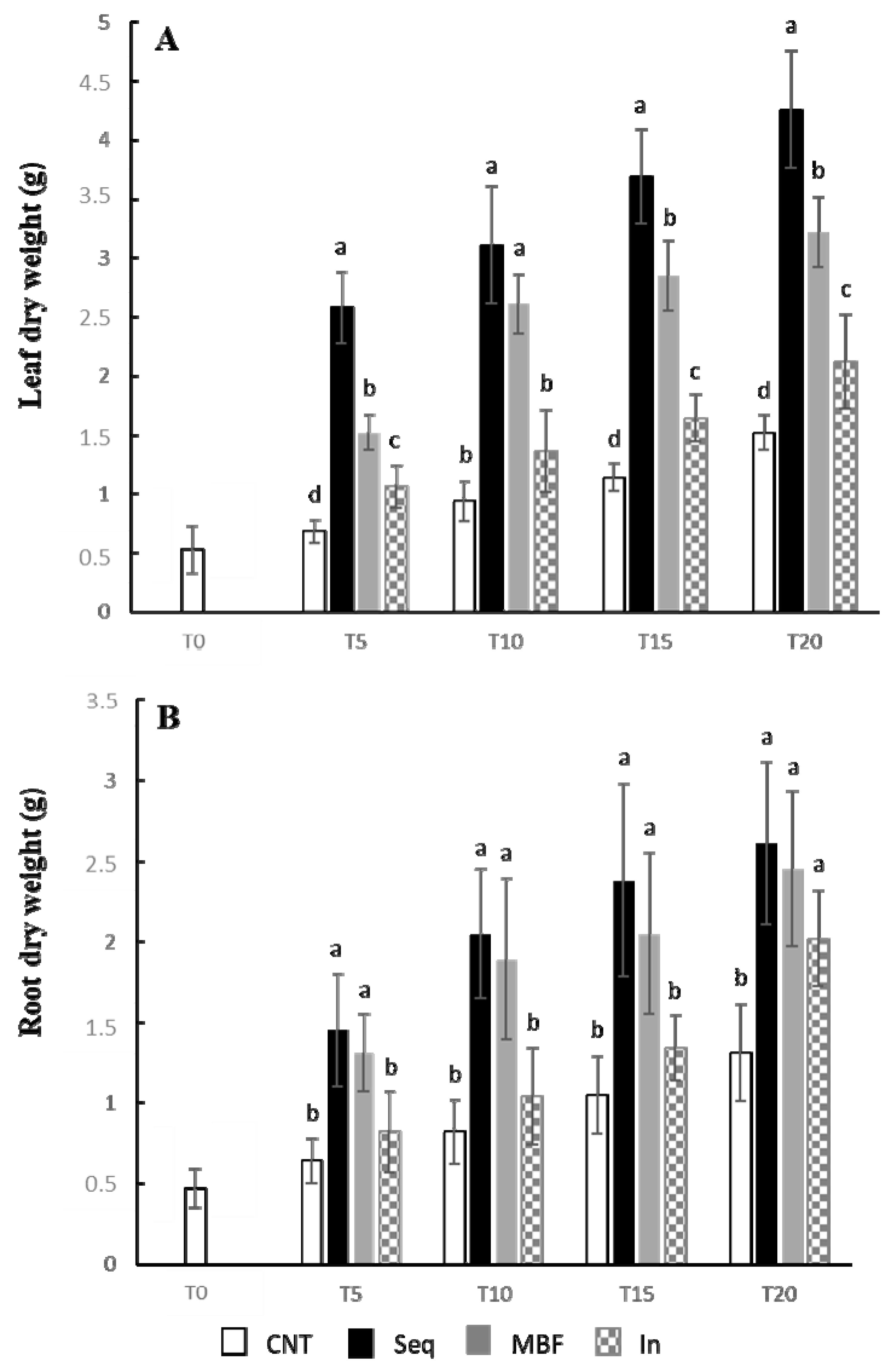
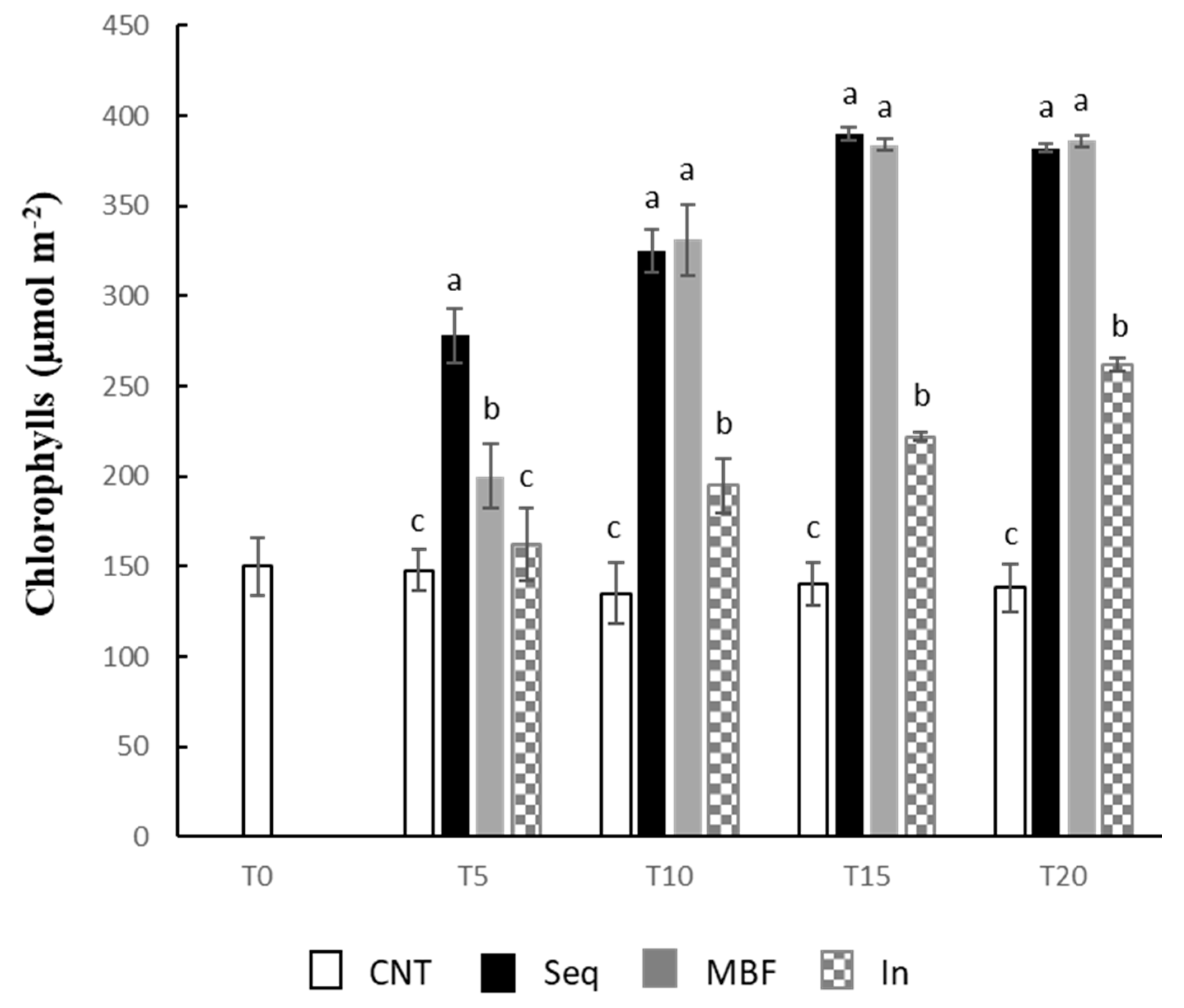
2.4. Determination of Fresh and Dry Weights, and Chlorophylls
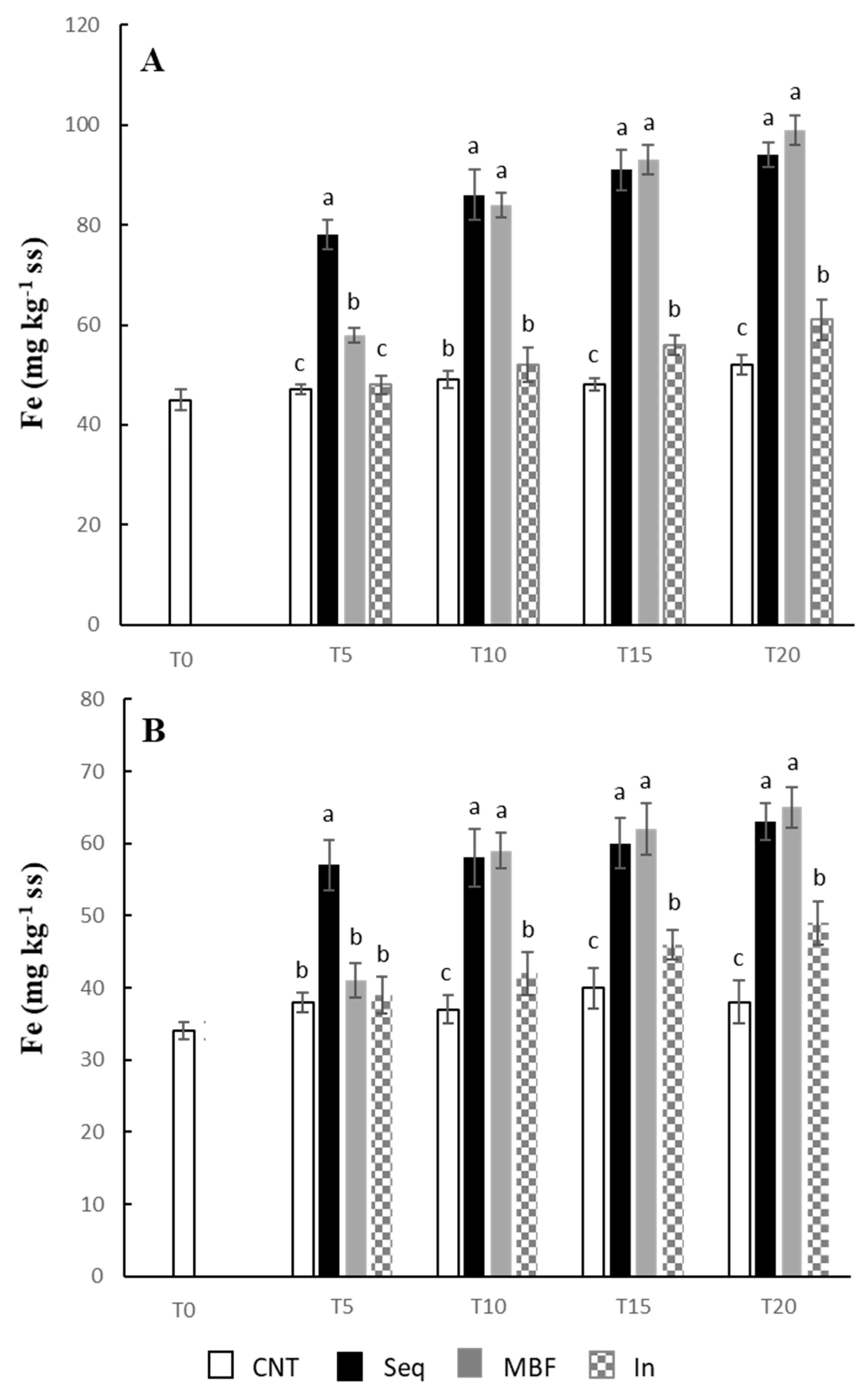
2.5. Iron Content in the Plants
2.6. Ferric Chelate Reductase Activity in Roots
2.7. Statistical Analysis
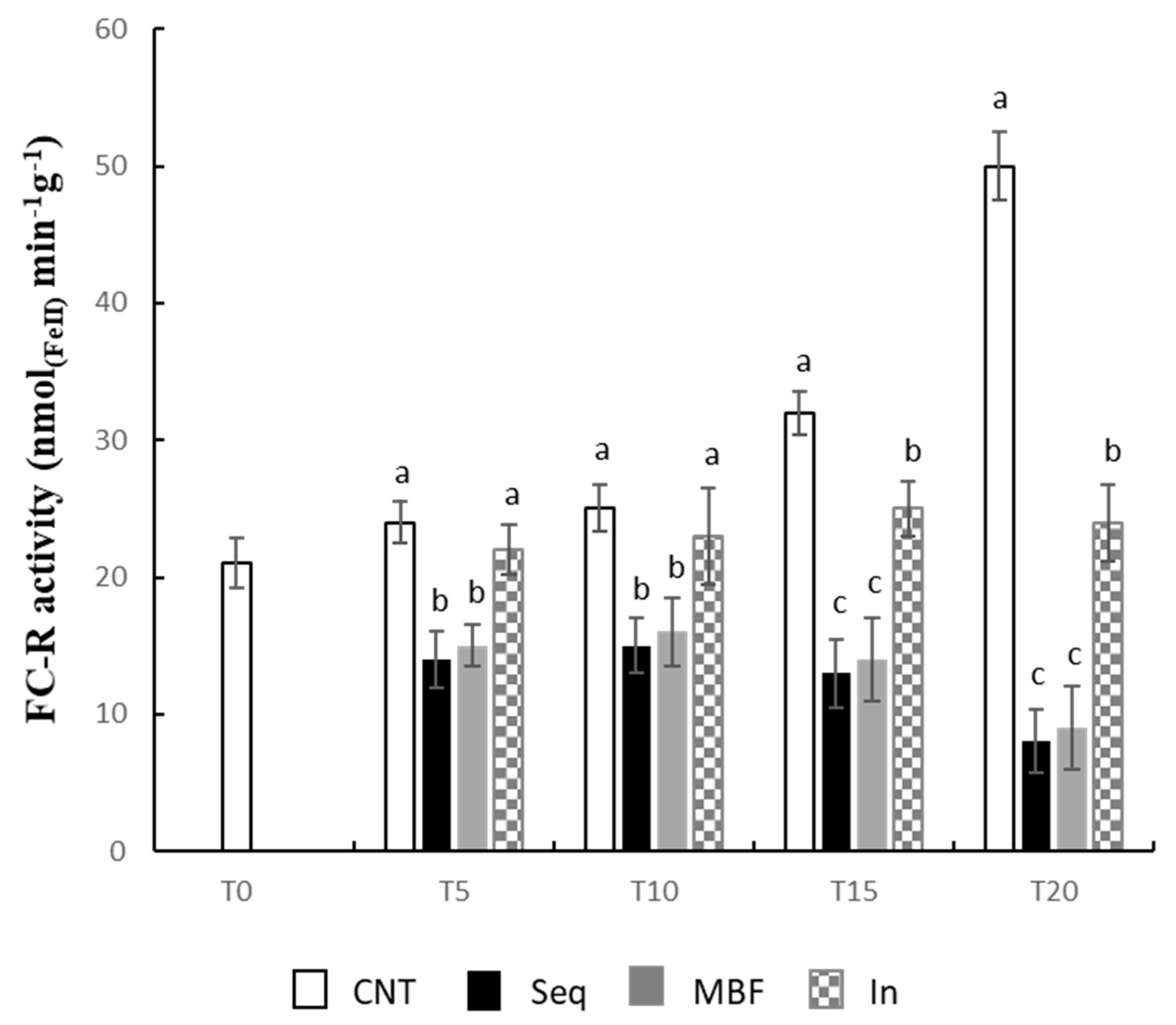
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Abadía, J.; Vázquez, S.; Rellán-Álvarez, R.; El-Jendoubi, H.; Abadía, A.; Álvarez-Fernández, A.; López-Millán, A.F. Towards a knowledge-based correction of iron chlorosis. Plant Physiol. Biochem. 2011, 49, 471–482. [Google Scholar] [CrossRef]
- Pushnik, J.C.; Miller, G.W. Iron regulation of chloroplast photosynthetic function: Mediation of PS-I development. J. Plant Nutr. 1989, 12, 407–421. [Google Scholar] [CrossRef]
- Terry, N.; Abadia, J. Function of iron in chloroplast. J. Plant Nutr. 1986, 9, 609–646. [Google Scholar] [CrossRef] [Green Version]
- Rutherford, A.W. Photosystem II, the water-splitting enzyme. Trends Biochem. Sci. 1989, 14, 227–232. [Google Scholar] [CrossRef]
- Abadia, J.; Abadia, A. Iron and plant pigments. In Iron Chelation in Plants and Soil Microorganisms; Barton, L.L., Hemming, B.C., Eds.; Academic Press: San Diego, CA, USA, 1993; pp. 327–343. [Google Scholar]
- Spiller, S.; Terry, N. Limiting factors in photosynthesis. 2. Iron stress diminished photochemical capacity by reducing the number of photosynthetic units. Plant Physiol. 1980, 65, 121–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabata-Pendias, A. Trace Elements in Soils and Plants; CRC: Boca Raton, FL, USA, 2001; pp. 76–78. [Google Scholar]
- Coulombe, B.A.; Chaney, R.L.; Wiebold, W.J. Bicarbonate directly induces iron chlorosis in susceptible soybean cultivars. Soil Sci. Soc. Amer. J. 1984, 48, 1297–1301. [Google Scholar] [CrossRef]
- Römheld, V.; Marschner, H. Evidence for a specific uptake system for iron phytosiderophores in roots of grasses. Plant Physiol. 1986, 80, 175–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephan, U.W. Intra- and intercellular iron trafficking and subcellular compartmentation within roots. Plant Soil 2002, 241, 19–25. [Google Scholar] [CrossRef]
- Imtiaz, M.; Rashid, A.; Khan, P.; Memon, M.Y.; Aslam, M. The role of micronutrients in crop production and human health. Pak. J. Bot. 2010, 42, 2565–2578. [Google Scholar]
- Taiz, L.; Zeiger, E. Plant Physiology, 4th ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2006. [Google Scholar]
- Cesco, S.; Nikolic, M.; Romheld, V.; Varanini, Z.; Pinton, R. Uptake of Fe-59 from soluble Fe-59-humate complexes by cucumber and barley plants. Plant Soil 2002, 241, 121–128. [Google Scholar] [CrossRef]
- Toulon, V.; Sentenac, H.; Thibaud, J.B.; Davidian, J.C.; Moulineaz, C.; Grignon, C. Role of apoplast acidification by H+ pump. Effect on the sensitivity to pH and CO2 of iron reduction by roots of Brassica napus L. Planta 1992, 186, 212–218. [Google Scholar] [PubMed]
- Marschner, H. Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: Hohenheim, Germany, 2011. [Google Scholar]
- Zanin, L.; Tomasi, N.; Cesco, S.; Varanini, Z.; Pinton, R. Humic substances contribute to plant iron nutrition acting as chelators and biostimulants. Front. Plant Sci. 2019, 675, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baglieri, A.; Borzí, D.; Abbate, C.; Ńegre, M.; Gennari, M. Removal of fenhexamid and pyrimethanil from aqueous solutions by clays and organoclays. J. Environ. Sci. Health Part B 2009, 44, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, Y.; Kong, S. Effects of Trichoderma asperellum and its siderophores on endogenous auxin in Arabidopsis thaliana under iron-deficiency stress. Int. Microbiol. 2020, 23, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Dimkpa, C.; Svatos, A.; Merten, D.; Buchel, G.; Kothe, E. Hydroxamate siderophores produced by Streptomyces acidiscabies E13 bind nickel and promote growth in cowpea (Vigna unguiculata L.) under nickel stress. Can. J. Microbiol. 2008, 54, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Siebner-Freibach, H.; Yariv, S.; Lapides, Y.; Hadar, Y.; Chen, Y. Thermo-FTIR spectroscopic study of the siderophore ferrioxamine B: Spectral analysis and stereochemical implications of iron chelation, pH, and temperature. J. Agric. Food Chem. 2005, 53, 3434–3443. [Google Scholar] [CrossRef]
- Colombo, C.; Palumbo, G.; He, J.Z.; Pinton, R.; Cesco, S. Review on iron availability in soil: Interaction of Fe minerals, plants, and microbes. J. Soils Sediments 2014, 14, 538–548. [Google Scholar] [CrossRef]
- Abbate, C.; Borzì, D.; Caboni, P.; Baglieri, A.; Gennari, M. Behavior of fenhexamid in soil and water. J. Environ. Sci. Health Part B 2007, 42, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, Y.Q. Effects of phosphate solubilization and phytohormone production of Trichoderma asperellum Q1 on promoting cucumber growth under salt stress. J. Integr. Agric. 2015, 14, 1588–1597. [Google Scholar] [CrossRef] [Green Version]
- Jin, C.W.; Ye, Y.Q.; Zheng, S.J. An underground tale: Contribution of microbial activity to plant iron acquisition via ecological processes. Ann. Bot. 2013, 113, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Schalk, I.J.; Hannauer, M.; Braud, A. New roles for bacterial siderophores in metal transport and tolerance. Environ. Microbiol. 2011, 13, 2844–2854. [Google Scholar] [CrossRef]
- Tagliavini, M.; Scudellari, D.; Marangoni, B.; Toselli, M. Nitrogen fertilization management in orchards to reconcile productivity and environmental aspects. Fertil. Res. 1996, 43, 93–102. [Google Scholar] [CrossRef]
- Tagliavini, M.; Baldi, E.; Lucchi, P.; Antonelli, M.; Sorrenti, G.; Baruzzi, G.; Faedi, W. Dynamics of nutrients uptake by strawberry plants (Fragaria × ananassa Dutch.) grown in soil and soilless culture. Eur. J. Agron. 2005, 23, 15–25. [Google Scholar] [CrossRef]
- Cojocaru, A.; Munteanu, N.; Petre, B.A.; Stan, T.; Teliban, G.C.; Vintu, C.; Stoleru, V. Biochemical and production of Rhubarb under growing technological factors. Rev. Chim. 2019, 70, 2000–2003. [Google Scholar] [CrossRef]
- Stoleru, V.; Munteanu, N.; Sellitto, V.M. New Approach of Organic Vegetable Systems; Aracne Editrice: Rome, Italy, 2014; pp. 56–71. [Google Scholar]
- Cojocaru, A.; Vlase, L.; Munteanu, N.; Stan, T.; Teliban, G.C.; Burducea, M.; Stoleru, V. Dynamic of phenolic compounds, antioxidant activity, and yield of Rhubarb under chemical, organic and biological fertilization. Plants 2020, 9, 355. [Google Scholar] [CrossRef]
- FAO. The Future of Food and Agriculture—Trends and Challenges; FAO: Rome, Italy, 2017. [Google Scholar]
- Barone, V.; Puglisi, I.; Fragalà, F.; Lo Piero, A.R.; Giuffrida, F.; Baglieri, A. Novel bioprocess for the cultivation of microalgae in hydroponic growing system of tomato plants. J. Appl. Phycol. 2019, 31, 465–470. [Google Scholar] [CrossRef]
- Violante, P. Metodi di analisi chimica del suolo. In Collana di Metodi Analitici per L’agricoltura; Sequi, P., Ed.; Francoangeli: Milano, Italy, 2000; pp. 46–51. [Google Scholar]
- Barone, V.; Puglisi, I.; Fragalà, F.; Stevanato, P.; Baglieri, A. Effect of living cells of microalgae or their extracts on soil enzyme activities. Arch. Agron. Soil Sci. 2019, 65, 712–726. [Google Scholar] [CrossRef]
- Puglisi, I.; Nicolosi, E.; Vanella, D.; Lo Piero, A.R.; Stagno, F.; Saitta, D.; Roccuzzo, G.; Consoli, S.; Baglieri, A. Physiological and biochemical responses of orange trees to different deficit irrigation regimes. Plants 2019, 8, 423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanni, A.; Anfossi, L.; Cignetti, A.; Baglieri, A.; Gennari, M. Degradation of pyrimethanil in soil: Influence of light, oxygen, and microbial activity. J. Environ. Sci. Health Part B 2006, 41, 67–80. [Google Scholar] [CrossRef]
- Puglisi, I.; La Bella, E.; Rovetto, E.I.; Lo Piero, A.R.; Baglieri, A. Biostimulant effect and biochemical response in lettuce seedlings treated with a Scenedesmus quadricauda extract. Plants 2020, 9, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pestana, M.; Correia, P.J.; Saavedra, T.; Gama, F.; Abadía, A.; de Varennes, A. Development and recovery of iron deficiency by iron resupply to roots or leaves of strawberry plants. Plant Physiol. Biochem. 2012, 53, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Puglisi, I.; Barone, V.; Sidella, S.; Coppa, M.; Broccanello, C.; Gennari, M.; Baglieri, A. Biostimulant activity of humic-like substances from agro-industrial waste on Chlorella vulgaris and Scenedesmus quadricauda. Eur. J. Phycol. 2018, 53, 433–442. [Google Scholar] [CrossRef]
- Barone, V.; Bertoldo, G.; Magro, F.; Broccanello, C.; Puglisi, I.; Baglieri, A.; Cagnin, M.; Concheri, G.; Squartini, A.; Pizzeghello, D.; et al. Molecular and morphological changes induced by Leonardite-based biostimulant in Beta vulgaris L. Plants 2019, 8, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bienfait, H.F.; Bino, R.J.; Vanderbliek, A.M.; Duivenvoorden, J.F.; Fontaine, J.M. Characterization of ferric reducing activity in roots of Fe-deficient Phaseolus vulgaris. Physiol. Plant. 1983, 59, 196–202. [Google Scholar] [CrossRef]
- Puglisi, I.; De Patrizio, A.; Schena, L.; Jung, T.; Evoli, M.; Pane, A.; Hoa, N.V.; Tri, M.V.; Wright, S.; Ramstedt, M.; et al. Two previously unknown Phytophthora species associated with brown rot of Pomelo (Citrus grandis) fruits in Vietnam. PLoS ONE 2017, 12, e0172085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erdal, I.; Kepenek, K.; Kizigöz, I. Effect of elemental sulphur and sulphur containing waste on the iron nutrition of strawberry plants grown in calcareous soil. Biol. Agric. Hortic. 2006, 23, 263–272. [Google Scholar] [CrossRef]
- Peris-Felipo, F.J.; Benavent-Gil, Y.; Hernandez-Apaolaza, L. Silicon beneficial effects on yield, fruit quality and shelf-life of strawberries grown in different culture substrates under different iron status. Plant Physiol. Biochem. 2020, 152, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Zaiter, H.Z.; Saad, I.; Nimah, M. Yield of iron-sprayed and non-sprayed strawberry cultivars grown on high calcareous soil. J. Plant Nutr. 1993, 16, 281–296. [Google Scholar] [CrossRef]
- Türemis, N.; Ozguven, A.L.; Paydas, S.; Idem, G. Effects of sequestrene Fe-138 as foliar and soil application on yield and earliness of some strawberry cultivars in the subtropics. Acta Hortic. 1997, 441, 369–374. [Google Scholar] [CrossRef]
- Spinelli, F.; Fiori, G.; Noferini, M.; Sprocatti, M.; Costa, G. A novel type of seaweed extract as a natural alternative to the use of iron chelates in strawberry production. Sci. Hortic. 2010, 125, 263–269. [Google Scholar] [CrossRef]
- Lucena, J.J.; Garate, A.; Ramon, A.M.; Manzanares, M. Iron nutrition of a hydroponic strawberry culture (Fragaria vesca L.) supplied with different Fe chelates. Plant Soil 1990, 123, 9–15. [Google Scholar] [CrossRef]
- Abadia, J.; Álvarez-Fernández, A.; Morales, F.; Sanz, M.; Abadia, A. Correction of iron chlorosis by foliar sprays. Acta Hortic. 2002, 594, 115–121. [Google Scholar] [CrossRef]
- Erdal, I.; Kepenek, K.; Kizigöz, I. Effect of foliar iron applications at different growth stages on iron and some nutrient concentrations in strawberry. Turk. J. Agric. For. 2004, 28, 421–427. [Google Scholar]
- Gilbert, E. A Guide to Soft Fruit Growing—The Strawberry; Codman Press: Boston, MA, USA, 2013. [Google Scholar]
- Timperio, A.M.; D’Amici, G.M.; Barta, C.; Loreto, G.M.; Zolla, L. Proteomics, pigment composition, and organization of thylakoid membranes in iron-deficient spinach leaves. J. Exp. Bot. 2006, 58, 3695–3710. [Google Scholar] [CrossRef] [Green Version]
- Larbi, A.; Abadía, A.; Morales, F.; Abadía, J. Fe resupply to Fe-deficient sugar beet plants leads to rapid changes in the violaxanthin cycle and other photosynthetic characteristics without significant de novo chlorophyll synthesis. Photosynth. Res. 2004, 79, 59–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajizadeh, H.S.; Heidari, B.; Bertoldo, G.; Della Lucia, M.C.; Magro, F.; Broccanello, C.; Baglieri, A.; Puglisi, I.; Squartini, A.; Campagna, G.; et al. Expression profiling of candidate genes in sugar beet leaves treated with Leonardite-based biostimulant. High-Throughput 2019, 8, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ipek, M.; Pirlak, L.; Esitken, A.; Donmez, M.F.; Turan, M.; Sahin, F. Plant Growth-Promoting Rhizobacteria (PGPR) increase yield, growth and nutrition of strawberry under high-calcareous soil conditions. J. Plant Nutr. 2014, 37, 990–1001. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Properties | Pre-Treatment |
|---|---|
| pH | 8.1 ± 0.2 |
| E.C (dS/m) | 0.179 ± 0.010 |
| CEC (meq/100 g) | 18 ± 1.1 |
| O.M. (%) | 2.0 * ± 0.8 |
| O.C. (%) | 1.14 * ± 0.6 |
| Active CaCO3 (%) | 19.5 ± 1.2 |
| Total CaCO3 (%) | 37.1 ± 3.2 |
| Total Fe (mg/kg) | 665 ± 21 |
| Extractable Fe (mg/kg) | 41.38 ± 5.3 |
| Available Fe (mg/kg) | 1.96 ± 0.2 |
| Readily available Fe (mg/kg) | 0.20 ± 0.02 |
| CPI | 113.9 ± 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puglisi, I.; Brida, S.; Stoleru, V.; Torino, V.; Sellitto, V.M.; Baglieri, A. Application of Novel Microorganism-Based Formulations as Alternative to the Use of Iron Chelates in Strawberry Cultivation. Agriculture 2021, 11, 217. https://doi.org/10.3390/agriculture11030217
Puglisi I, Brida S, Stoleru V, Torino V, Sellitto VM, Baglieri A. Application of Novel Microorganism-Based Formulations as Alternative to the Use of Iron Chelates in Strawberry Cultivation. Agriculture. 2021; 11(3):217. https://doi.org/10.3390/agriculture11030217
Chicago/Turabian StylePuglisi, Ivana, Sergio Brida, Vasile Stoleru, Valentina Torino, Vincenzo Michele Sellitto, and Andrea Baglieri. 2021. "Application of Novel Microorganism-Based Formulations as Alternative to the Use of Iron Chelates in Strawberry Cultivation" Agriculture 11, no. 3: 217. https://doi.org/10.3390/agriculture11030217
APA StylePuglisi, I., Brida, S., Stoleru, V., Torino, V., Sellitto, V. M., & Baglieri, A. (2021). Application of Novel Microorganism-Based Formulations as Alternative to the Use of Iron Chelates in Strawberry Cultivation. Agriculture, 11(3), 217. https://doi.org/10.3390/agriculture11030217