
Comprehensive Analysis of the Histone Deacetylase Gene Family in Chinese Cabbage (Brassica rapa): From Evolution and Expression Pattern to Functional Analysis of BraHDA3

Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification and Characterization of HDAC Family in B. rapa
2.2. Gene Duplication and Calculating Duplication Times of BraHDAC Genes
2.3. Plant Growth and Treatment Conditions
2.4. The Analysis of BraHDAC Expression
2.5. Production of Recombinant BraHDA3 in E. coli, and the Analysis of HDAC Activity
2.6. Statistical Analyses
3. Results and Discussion
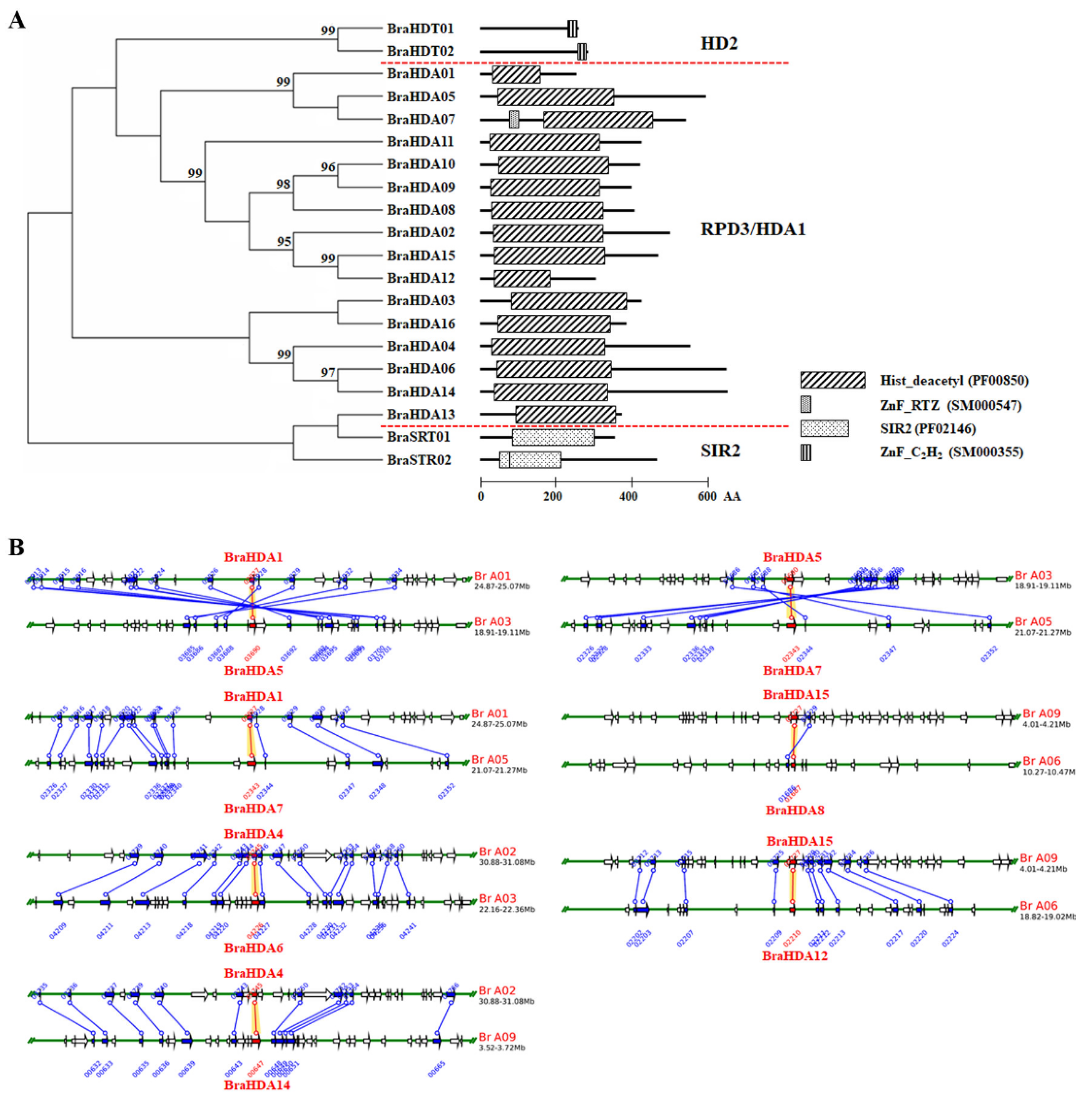
3.1. Identification of Histone Deacetylases in Chinese Cabbage
3.2. Gene Duplication Analysis of BraHDACs
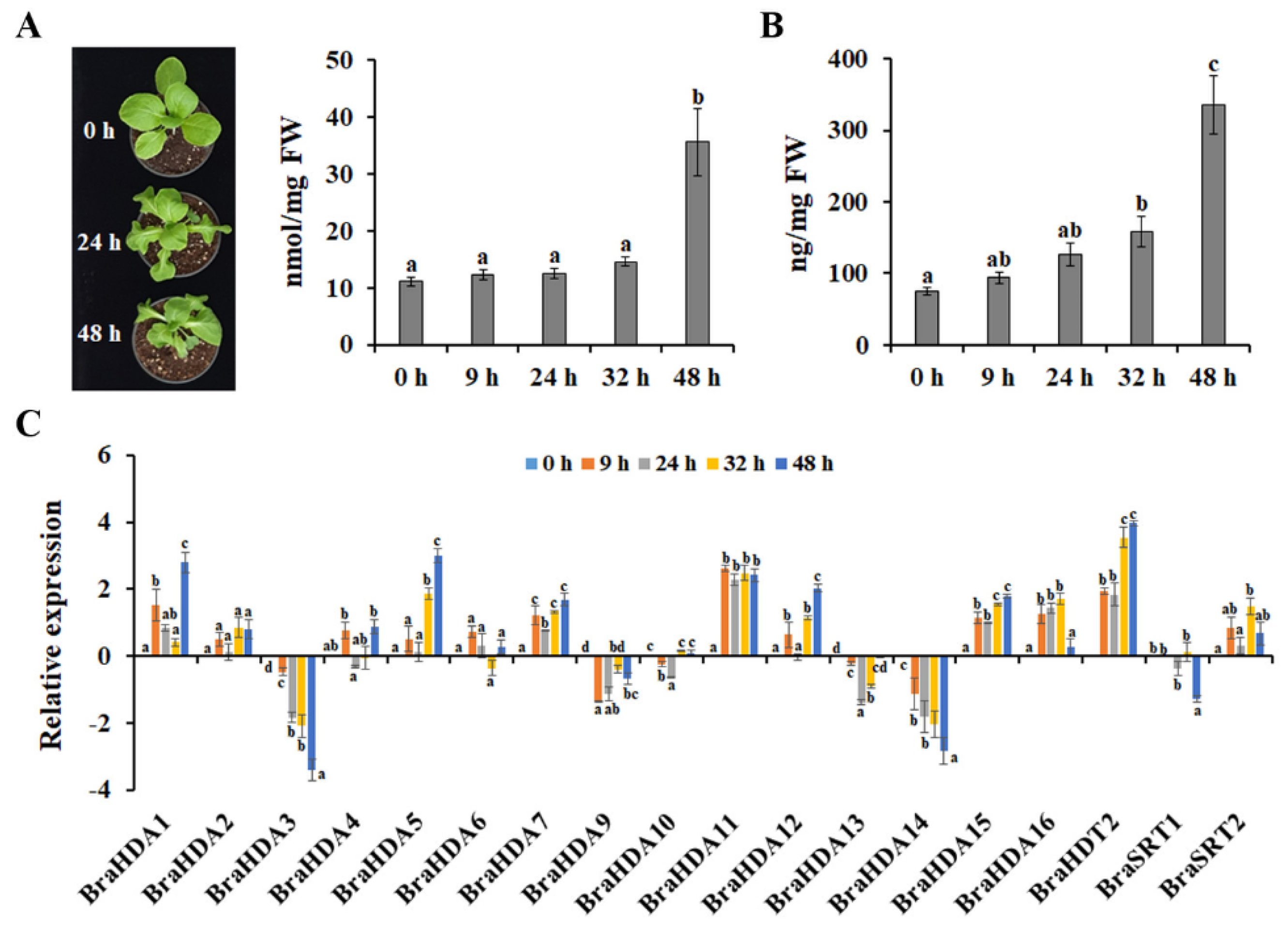
3.3. Effects of Heat Stress on Acetylation Status of Histone H3 in Chinese Cabbage
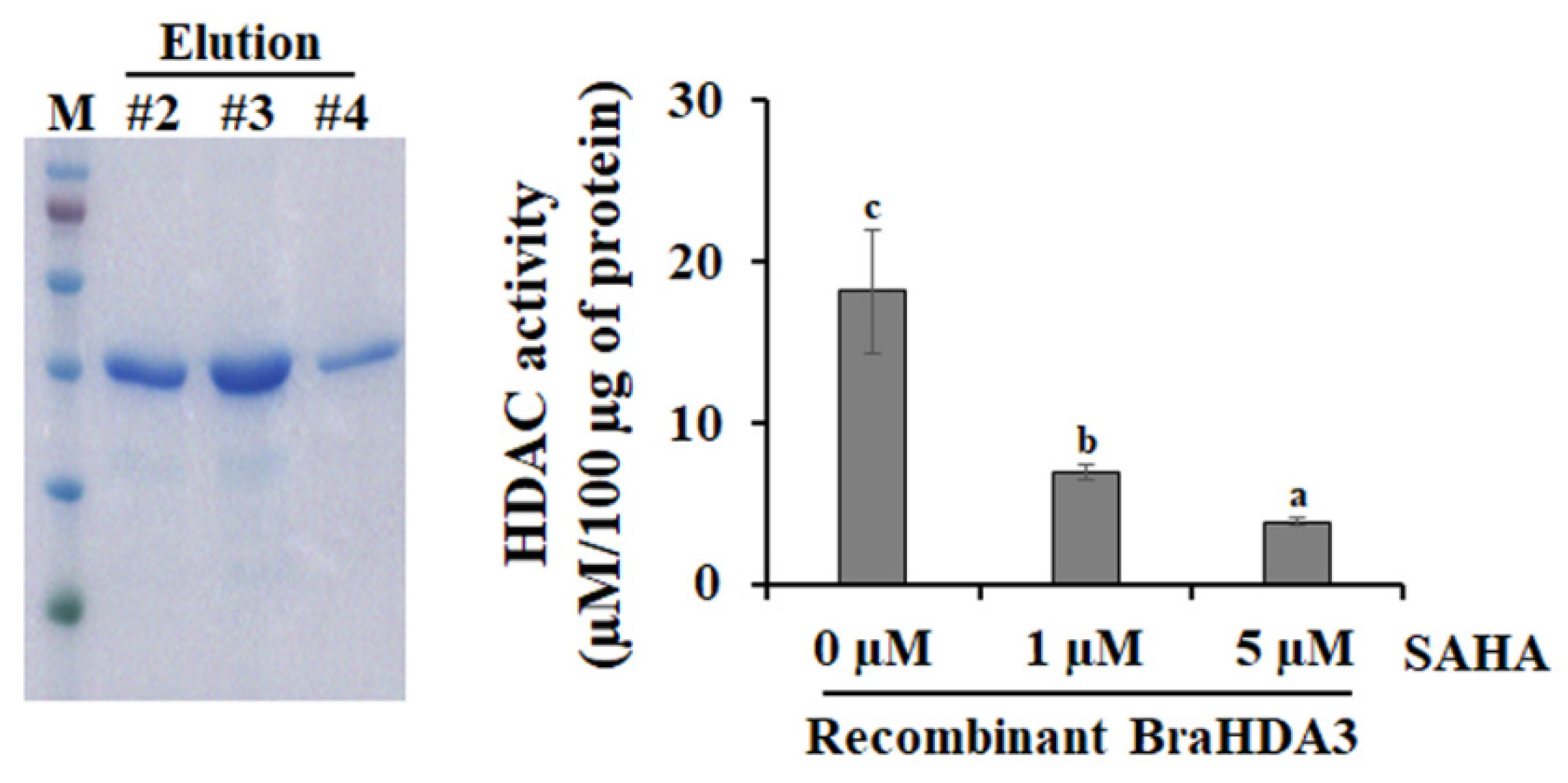
3.4. Histone Deacetylase Activity of BarHDA3
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lämke, J.; Bäurle, I. Epigenetic and chromatin-based mechanisms in environmental stress adaptation and stress memory in plants. Genome Biol. 2017, 18, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Berr, A.; Shafiq, S.; Shen, W.H. Histone modifications in transcriptional activation during plant development. Biochim. Biophys. Acta Gene Regul. Mech. 2011, 1809, 567–576. [Google Scholar] [CrossRef]
- Chen, X.; Ding, A.B.; Zhong, X. Functions and mechanisms of plant histone deacetylases. Sci. China Life Sci. 2020, 63, 206–216. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.M.; Pontes, O.; Searle, I.; Yelina, N.; Yousafzai, F.K.; Herr, A.J.; Pikaard, C.S.; Baulcombe, D.C. An SNF2 protein associated with nuclear RNA silencing and the spread of a silencing signal between cells in Arabidopsis. Plant Cell 2007, 19, 1507–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, R.; Müller, A.; Napoli, C.A.; Selinger, D.A.; Pikaard, C.S.; Richards, E.J.; Bender, J.; Mount, D.W.; Jorgensen, R.A. Analysis of histone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatin modification among multicellular eukaryotes. Nucleic Acids Res. 2002, 30, 5036–5055. [Google Scholar] [CrossRef] [Green Version]
- Hollender, C.; Liu, Z. Histone deacetylase genes in arabidopsis development. J. Integr. Plant Biol. 2008, 57, 875–885. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Qin, F.; Huang, L.; Sun, Q.; Li, C.; Zhao, Y.; Zhou, D.X. Rice histone deacetylase genes display specific expression patterns and developmental functions. Biochem. Biophys. Res. Commun. 2009, 388, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Liew, L.C.; Singh, M.B.; Bhalla, P.L. An RNA-Seq Transcriptome Analysis of Histone Modifiers and RNA Silencing Genes in Soybean during Floral Initiation Process. PLoS ONE 2013, 8, e77502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aquea, F.; Timmermann, T.; Arce-Johnson, P. Analysis of histone acetyltransferase and deacetylase families of Vitis vinifera. Plant Physiol. Biochem. 2010, 48, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.; Ying, P.; Liu, X.; Li, C.; Xia, R.; Li, J.; Zhao, M. Genome-wide identification of histone modifiers and their expression patterns during fruit abscission in litchi. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Zhang, K.; Yu, L.; Pang, X.; Cao, H.; Si, H.; Zang, J.; Xing, J.; Dong, J. In silico analysis of maize HDACs with an emphasis on their response to biotic and abiotic stresses. PeerJ 2020, 8, e8539. [Google Scholar] [CrossRef] [Green Version]
- Imran, M.; Shafiq, S.; Naeem, M.K.; Widemann, E.; Munir, M.Z.; Jensen, K.B.; Wang, R.R.C. Histone deacetylase (HDAC) gene family in allotetraploid cotton and its diploid progenitors: In silico identification, molecular characterization, and gene expression analysis under multiple abiotic stresses, DNA damage and phytohormone treatments. Int. J. Mol. Sci. 2020, 21, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Lu, Y.; Zhao, Y.; Zhou, D.X. Histone Acetylation Dynamics Integrates Metabolic Activity to Regulate Plant Response to Stress. Front. Plant Sci. 2019, 10, 1236. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; To, T.K.; Matsui, A.; Tanoi, K.; Kobayashi, N.I.; Matsuda, F.; Habu, Y.; Ogawa, D.; Sakamoto, T.; Matsunaga, S.; et al. Acetate-mediated novel survival strategy against drought in plants. Nat. Plants 2017, 3. [Google Scholar] [CrossRef]
- Zheng, Y.; Ding, Y.; Sun, X.; Xie, S.; Wang, D.; Liu, X.; Su, L.; Wei, W.; Pan, L.; Zhou, D.X. Histone deacetylase HDA9 negatively regulates salt and drought stress responsiveness in Arabidopsis. J. Exp. Bot. 2016, 67, 1703–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueda, M.; Matsui, A.; Nakamura, T.; Abe, T.; Sunaoshi, Y.; Shimada, H.; Seki, M. Versatility of HDA19-deficiency in increasing the tolerance of Arabidopsis to different environmental stresses. Plant Signal. Behav. 2018, 13, e1475808. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhang, L.; Duan, J.; Miki, B.; Wu, K. Histone Deacetylase19 is involved in jasmonic acid and ethylene signaling of pathogen response in Arabidopsis. Plant Cell 2005, 17, 1196–1204. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Li, M.; Gu, D.; Liu, X.; Zhang, J.; Wu, K.; Zhang, X.; Teixeira Da Silva, J.A.; Duan, J. Involvement of rice histone deacetylase HDA705 in seed germination and in response to ABA and abiotic stresses. Biochem. Biophys. Res. Commun. 2016, 470, 439–444. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, J.; Zhang, W.; Wu, K.; Zheng, F.; Tian, L.; Liu, X.; Duan, J. Expression and functional analysis of the plant-specific histone deacetylase HDT701 in rice. Front. Plant Sci. 2015, 5. [Google Scholar] [CrossRef] [Green Version]
- Latrasse, D.; Jégu, T.; Li, H.; de Zelicourt, A.; Raynaud, C.; Legras, S.; Gust, A.; Samajova, O.; Veluchamy, A.; Rayapuram, N.; et al. MAPK-triggered chromatin reprogramming by histone deacetylase in plant innate immunity. Genome Biol. 2017, 18, 131. [Google Scholar] [CrossRef]
- Eom, S.H.; Hyun, T.K. Histone acetyltransferases (HATs) in chinese cabbage: Insights from histone H3 acetylation and expression profiling of HATs in response to abiotic stresses. J. Am. Soc. Hortic. Sci. 2018, 143, 296–303. [Google Scholar] [CrossRef] [Green Version]
- Koch, M.A.; Haubold, B.; Mitchell-Olds, T. Comparative evolutionary analysis of chalcone synthase and alcohol dehydrogenase loci in Arabidopsis, Arabis, and related genera (Brassicaceae). Mol. Biol. Evol. 2000, 17, 1483–1498. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Labbe, H.; Sridha, S.; Wang, L.; Tian, L.; Latoszek-Green, M.; Yang, Z.; Brown, D.; Miki, B.; Wu, K. Expression and function of HD2-type histone deacetylases in Arabidopsis development. Plant J. 2004, 38, 715–724. [Google Scholar] [CrossRef]
- Yang, C.; Shen, W.; Chen, H.; Chu, L.; Xu, Y.; Zhou, X.; Liu, C.; Chen, C.; Zeng, J.; Liu, J.; et al. Characterization and subcellular localization of histone deacetylases and their roles in response to abiotic stresses in soybean. BMC Plant Biol. 2018, 18, 226. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Sun, Q.; Qin, F.; Li, C.; Zhao, Y.; Zhou, D.X. Down-regulation of a Silent Information Regulator2-related histone deacetylase gene, OsSRT1, induces DNA fragmentation and cell death in rice. Plant Physiol. 2007, 144, 1508–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giesecke, A.V.; Fang, R.; Joung, J.K. Synthetic protein-protein interaction domains created by shuffling Cys 2His2 zinc-fingers. Mol. Syst. Biol. 2006, 2, 0011. [Google Scholar] [CrossRef] [Green Version]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.H. Evolution of gene duplication in plants. Plant Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef] [Green Version]
- Freeling, M. Bias in plant gene content following different sorts of duplication: Tandem, whole-genome, segmental, or by transposition. Annu. Rev. Plant Biol. 2009, 60, 433–453. [Google Scholar] [CrossRef]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Blanc, G.; Wolfe, K.H. Widespread paleopolyploidy in model plant species inferred from age distributions of duplicate genes. Plant Cell 2004, 16, 1667–1678. [Google Scholar] [CrossRef] [Green Version]
- Lysak, M.A.; Koch, M.A.; Pecinka, A.; Schubert, I. Chromosome triplication found across the tribe Brassiceae. Genome Res. 2005, 15, 516–525. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Hu, S.; Yu, Q.; Wang, C.; Yang, Y.; Sun, H.; Yang, Y.; Sun, X. Genome-wide identification and characterization of BrrTCP transcription factors in Brassica rapa ssp. rapa. Front. Plant Sci. 2017, 8, 1588. [Google Scholar] [CrossRef] [Green Version]
- Cheng, F.; Wu, J.; Fang, L.; Sun, S.; Liu, B.; Lin, K.; Bonnema, G.; Wang, X. Biased gene fractionation and dominant gene expression among the subgenomes of Brassica rapa. PLoS ONE 2012, 7, e36442. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef]
- Morales, M.; Munné-Bosch, S. Malondialdehyde: Facts and artifacts. Plant Physiol. 2019, 180, 1246–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, W.T.; Lin, B.; Zhang, M.; Hua, X.J. Proline accumulation is inhibitory to arabidopsis seedlings during heat stress. Plant Physiol. 2011, 156, 1921–1933. [Google Scholar] [CrossRef] [Green Version]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [Green Version]
- Alinsug, M.V.; Yu, C.W.; Wu, K. Phylogenetic analysis, subcellular localization, and expression patterns of RPD3/HDA1 family histone deacetylases in plants. BMC Plant Biol. 2009, 9, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, C.; Wang, X.; Yu, J.; Wu, J.; Li, W.; Huang, J.; Dong, C.; Hua, W.; Liu, S. Comprehensive analysis of RNA-seq data reveals the complexity of the transcriptome in Brassica rapa. BMC Genom. 2013, 14, 689. [Google Scholar] [CrossRef] [Green Version]
- Ueda, M.; Matsui, A.; Watanabe, S.; Kobayashi, M.; Saito, K.; Tanaka, M.; Ishida, J.; Kusano, M.; Seo, M.; Seki, M. Transcriptome analysis of the hierarchical response of histone deacetylase proteins that respond in an antagonistic manner to salinity stress. Front. Plant. Sci. 2019, 10, 1323. [Google Scholar] [CrossRef]
- Hartl, M.; Füßl, M.; Boersema, P.J.; Jost, J.O.; Kramer, K.; Bakirbas, A.; Sindlinger, J.; Plöchinger, M.; Leister, D.; Uhrig, G.; et al. Lysine acetylome profiling uncovers novel histone deacetylase substrate proteins in Arabidopsis. Mol. Syst. Biol. 2017, 13, 949. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Lee, H.Y.; Back, K. Rice histone deacetylase 10 and Arabidopsis histone deacetylase 14 genes encode N-acetylserotonin deacetylase, which catalyzes conversion of N-acetylserotonin into serotonin, a reverse reaction for melatonin biosynthesis in plants. J. Pineal Res. 2018, 64, e12460. [Google Scholar] [CrossRef] [PubMed]
- Buttar, Z.A.; Wu, S.N.; Arnao, M.B.; Wang, C.; Ullah, I.; Wang, C. Melatonin suppressed the heat stress-induced damage in wheat seedlings by modulating the antioxidant machinery. Plants 2020, 9, 809. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Name | Gene ID | Location | CDS (bp) | AA | Intron Nr. | pI | kDa | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|
| BraHDA1 | Brara.A02927.1 | A01:24968034..24970355 | 765 | 254 | 7 | 4.98 | 28.8 | Cytosol |
| BraHDA2 | Brara.A03767.1 | A01:29972774..29976555 | 1509 | 502 | 5 | 5.17 | 56.0 | Nucleus |
| BraHDA3 | Brara.A00453.1 | A01:2203688..2206445 | 1278 | 425 | 8 | 6.06 | 45.9 | Chloroplast |
| BraHDA4 | Brara.B03745.1 | A02:30976246..30978653 | 1668 | 555 | 8 | 5.93 | 62.0 | Cytosol |
| BraHDA5 | Brara.C03690.1 | A03:19011956..19014935 | 1797 | 598 | 13 | 6.00 | 65.8 | Nucleus |
| BraHDA6 | Brara.C04226.1 | A03:22262637..22266032 | 1956 | 651 | 12 | 5.23 | 72.1 | Cytosol |
| BraHDA7 | Brara.E02343.1 | A05:21170590..21174613 | 1638 | 545 | 16 | 5.60 | 59.8 | Nucleus |
| BraHDA8 | Brara.F01687.1 | A06:10370998..10372699 | 1230 | 409 | 4 | 4.97 | 45.4 | Cytosol |
| BraHDA9 | Brara.F01690.1 | A06:10386796..10389169 | 1200 | 399 | 4 | 5.82 | 44.4 | Cytosol |
| BraHDA10 | Brara.F01971.1 | A06:14950658..14953055 | 1272 | 423 | 4 | 8.70 | 47.6 | Mitochondrial |
| BraHDA11 | Brara.F02002.1 | A06:16050873..16053859 | 1281 | 426 | 13 | 5.32 | 48.7 | Cytosol |
| BraHDA12 | Brara.F02210.1 | A06:18915988..18918237 | 921 | 306 | 4 | 4.96 | 34.3 | Nucleus |
| BraHDA13 | Brara.F02773.1 | A06:22749037..22751166 | 1119 | 372 | 9 | 6.17 | 42.0 | Cytosol |
| BraHDA14 | Brara.I00647.1 | A09:3621688..3625470 | 1959 | 652 | 12 | 5.32 | 71.5 | Peroxisome |
| BraHDA15 | Brara.I00727.1 | A09:4109663..4112303 | 1413 | 470 | 5 | 4.94 | 52.7 | Nucleus |
| BraHDA16 | Brara.I05307.1 | A09:43269577..43271821 | 1161 | 386 | 2 | 5.68 | 42.2 | Cytosol |
| BraHDT1 | Brara.A02332.1 | A01:17662954..17671061 | 786 | 261 | 7 | 6.52 | 28.0 | Nucleus |
| BraHDT2 | Brara.J02801.1 | A10:19273043..19274967 | 858 | 285 | 6 | 4.52 | 31.2 | Nucleus |
| BraSRT1 | Brara.B00303.1 | A02:1417334..1419795 | 1047 | 357 | 8 | 8.96 | 39.7 | Mitochondrial |
| BraSRT2 | Brara.C01390.1 | A03:6592244..6595515 | 1407 | 468 | 13 | 6.49 | 52.4 | Nucleus |
| Gene 1 | Gene 2 | S | N | Ks | Ka | Ka/Ks | Mya |
|---|---|---|---|---|---|---|---|
| BraHDA8 | BraHDA9 | 298.2 | 898.8 | 0.1640 | 0.1429 | 0.8713 | 5.47 |
| BraHDA1 | BraHDA5 | 176.0 | 574.0 | 1.2612 | 0.6244 | 0.4951 | 42.04 |
| BraHDA1 | BraHDA7 | 165.2 | 545.8 | 1.2066 | 0.4168 | 0.3454 | 40.22 |
| BraHDA4 | BraHDA6 | 371.3 | 1194.7 | 0.8011 | 0.3165 | 0.3951 | 26.70 |
| BraHDA4 | BraHDA14 | 399.2 | 1229.8 | 0.8870 | 0.3867 | 0.4360 | 29.57 |
| BraHDA5 | BraHDA7 | 319.5 | 925.5 | 0.4043 | 0.1344 | 0.3324 | 13.48 |
| BraHDA15 | BraHDA8 | 302.9 | 906.1 | 1.1871 | 0.3172 | 0.2672 | 39.57 |
| BraHDA15 | BraHDA12 | 215.5 | 693.5 | 0.3818 | 0.0342 | 0.0895 | 12.73 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eom, S.H.; Hyun, T.K. Comprehensive Analysis of the Histone Deacetylase Gene Family in Chinese Cabbage (Brassica rapa): From Evolution and Expression Pattern to Functional Analysis of BraHDA3. Agriculture 2021, 11, 244. https://doi.org/10.3390/agriculture11030244
Eom SH, Hyun TK. Comprehensive Analysis of the Histone Deacetylase Gene Family in Chinese Cabbage (Brassica rapa): From Evolution and Expression Pattern to Functional Analysis of BraHDA3. Agriculture. 2021; 11(3):244. https://doi.org/10.3390/agriculture11030244
Chicago/Turabian StyleEom, Seung Hee, and Tae Kyung Hyun. 2021. "Comprehensive Analysis of the Histone Deacetylase Gene Family in Chinese Cabbage (Brassica rapa): From Evolution and Expression Pattern to Functional Analysis of BraHDA3" Agriculture 11, no. 3: 244. https://doi.org/10.3390/agriculture11030244
APA StyleEom, S. H., & Hyun, T. K. (2021). Comprehensive Analysis of the Histone Deacetylase Gene Family in Chinese Cabbage (Brassica rapa): From Evolution and Expression Pattern to Functional Analysis of BraHDA3. Agriculture, 11(3), 244. https://doi.org/10.3390/agriculture11030244