
Seasonal Dynamics of Soil Moisture in an Integrated-Crop-Livestock-Forestry System in Central-West Brazil

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
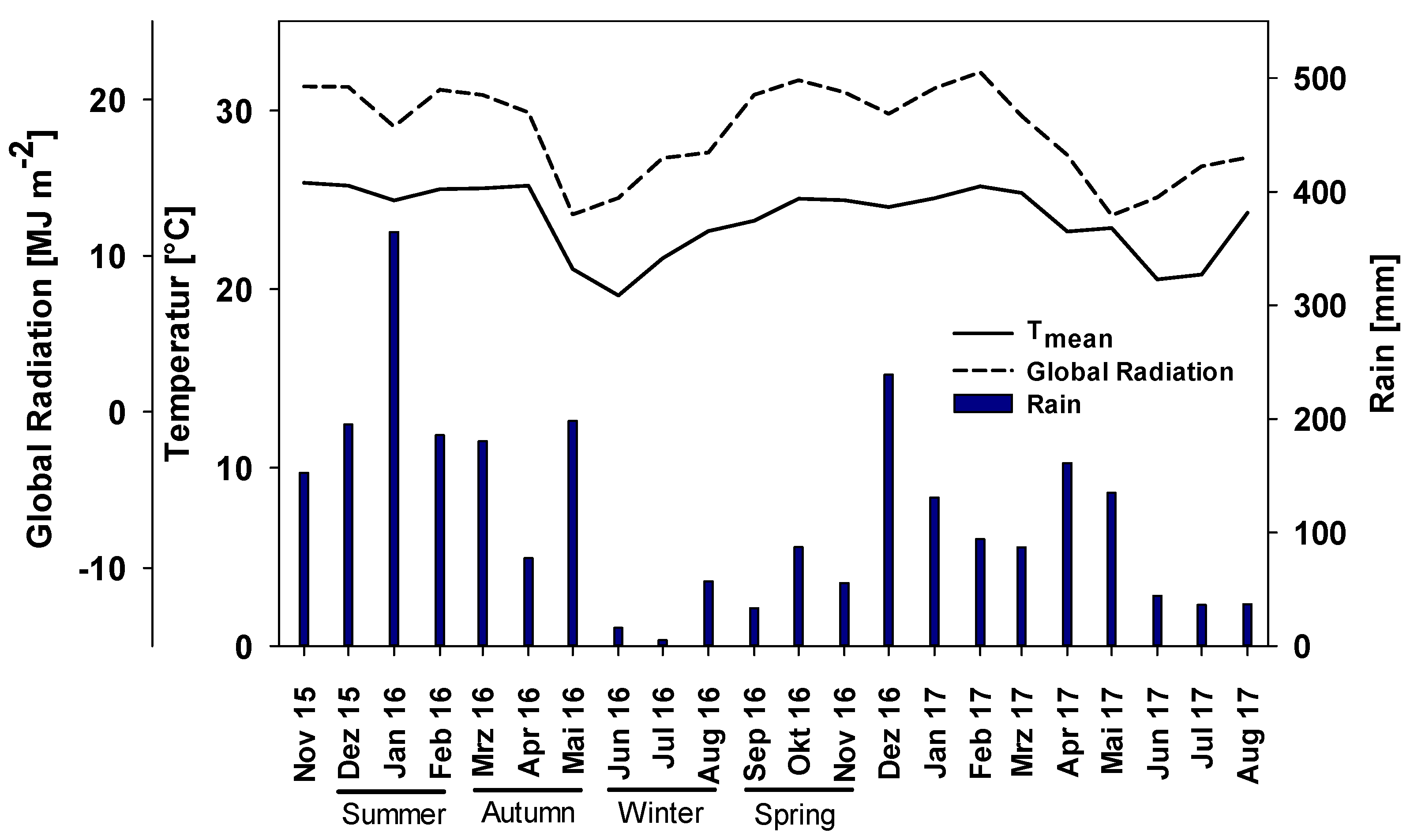
2.1. Experimental Site
2.2. ICLF System
2.3. PAR, SM and AGBM
2.4. Data Analysis
3. Results
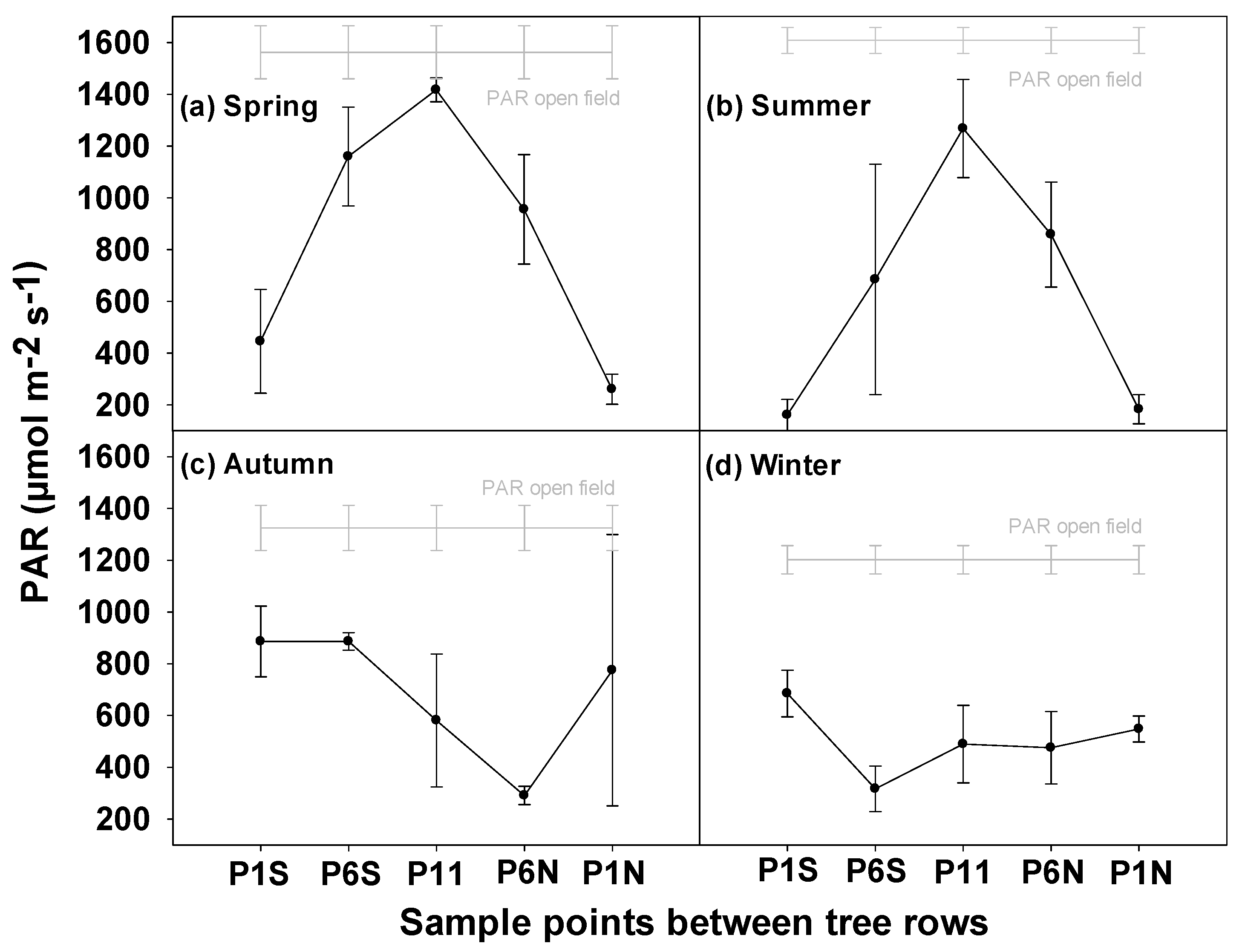
3.1. PAR between the Tree Rows during Different Seasons
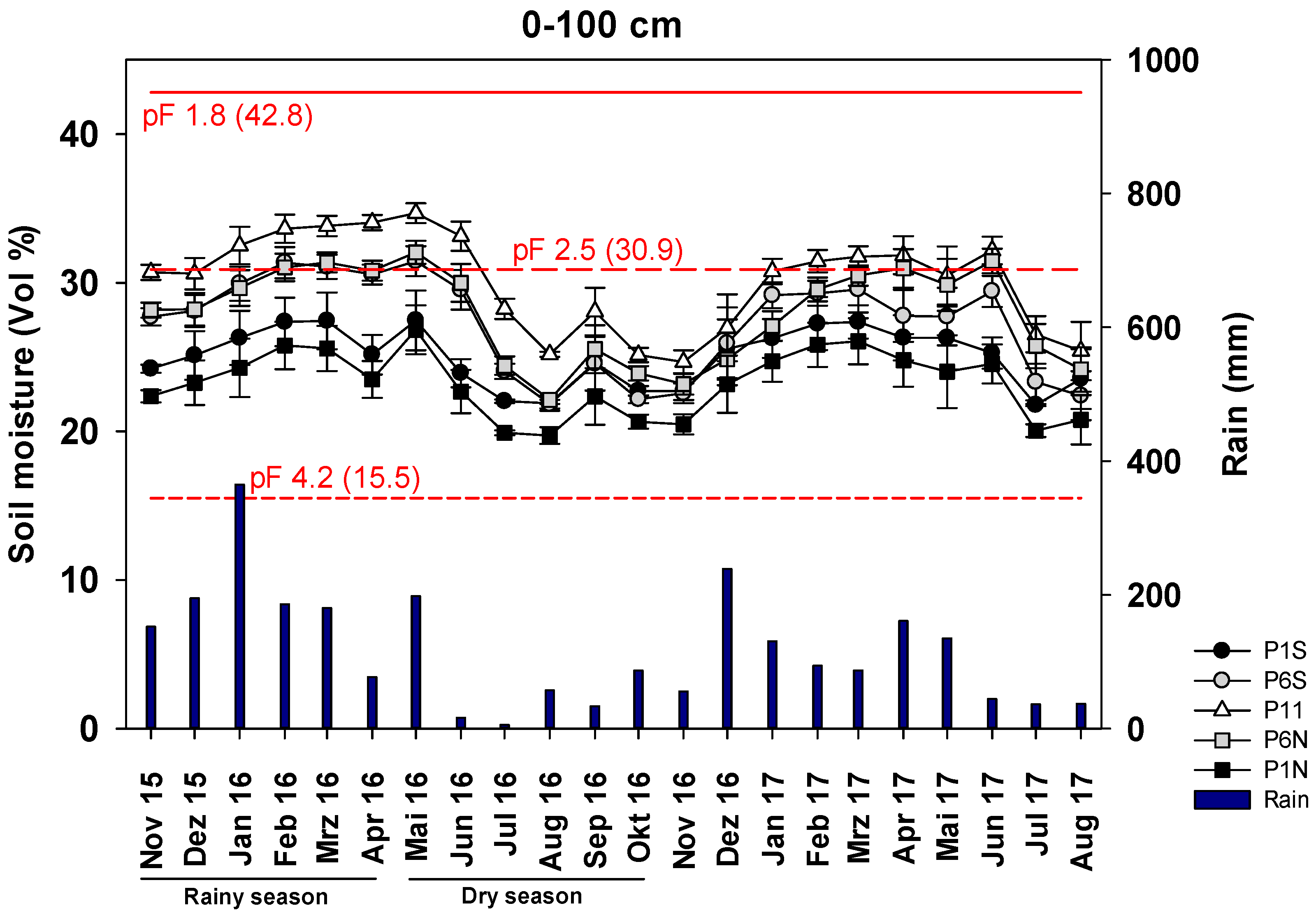
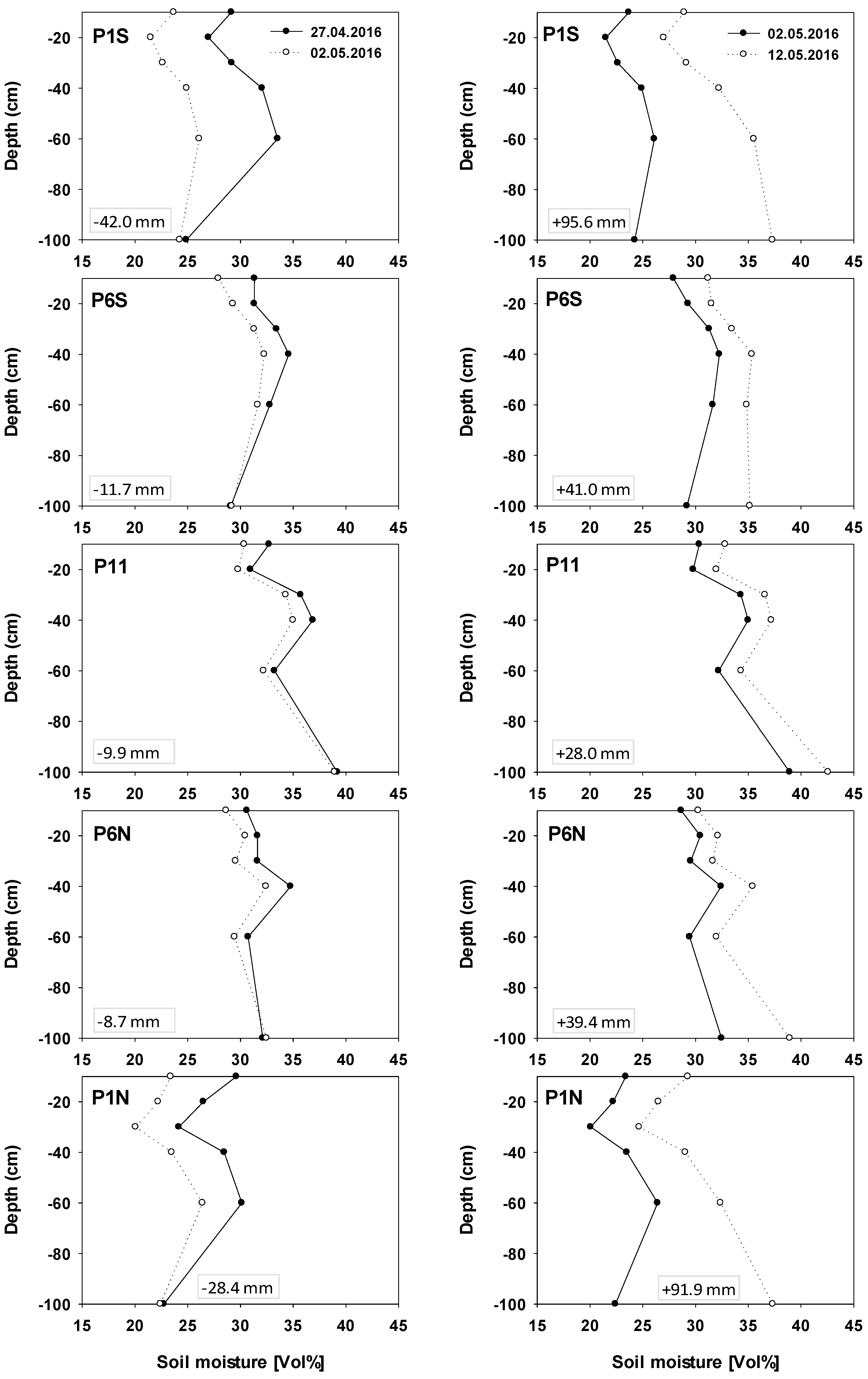
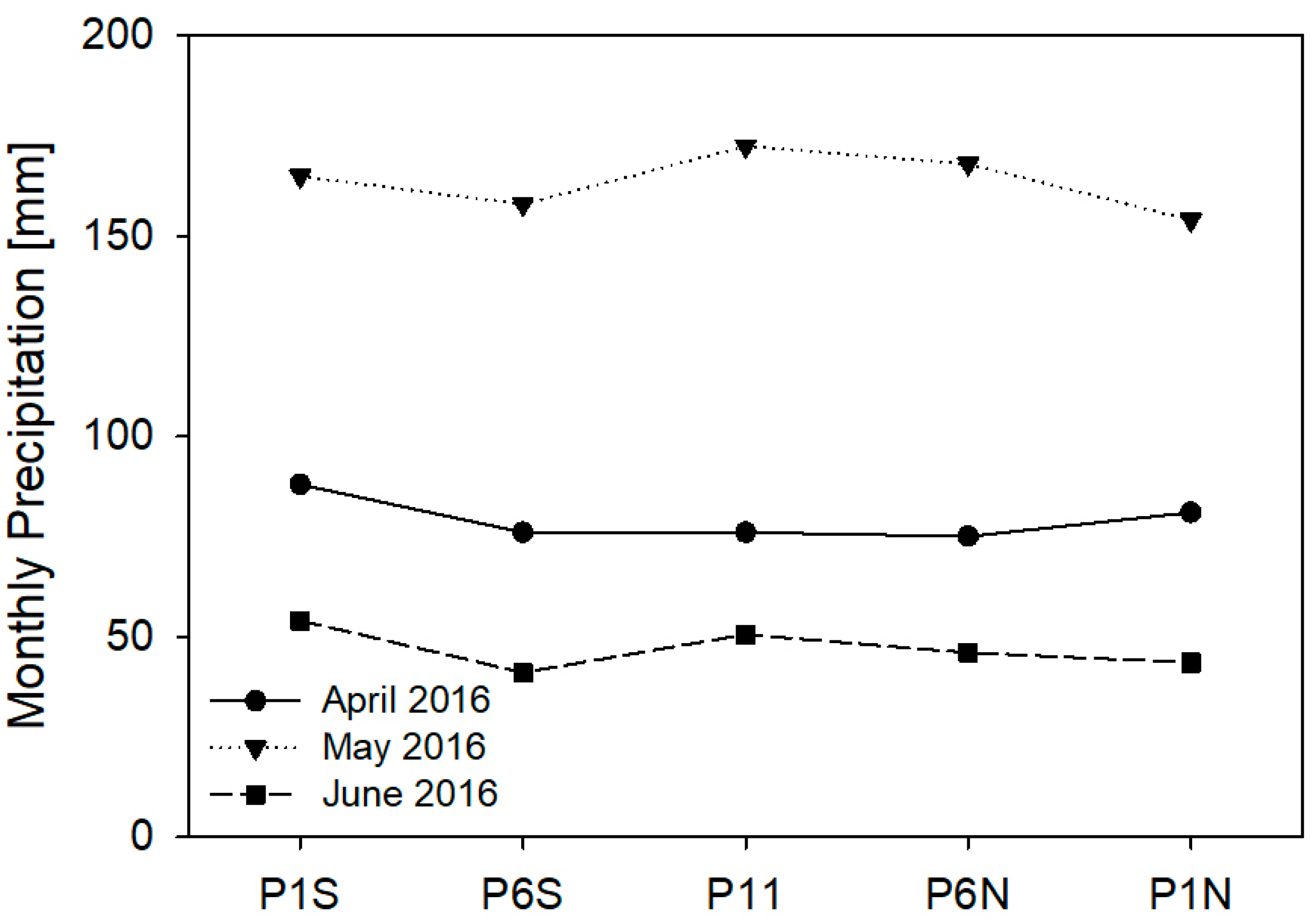
3.2. SM in ICLF between the Tree Rows during Different Seasons
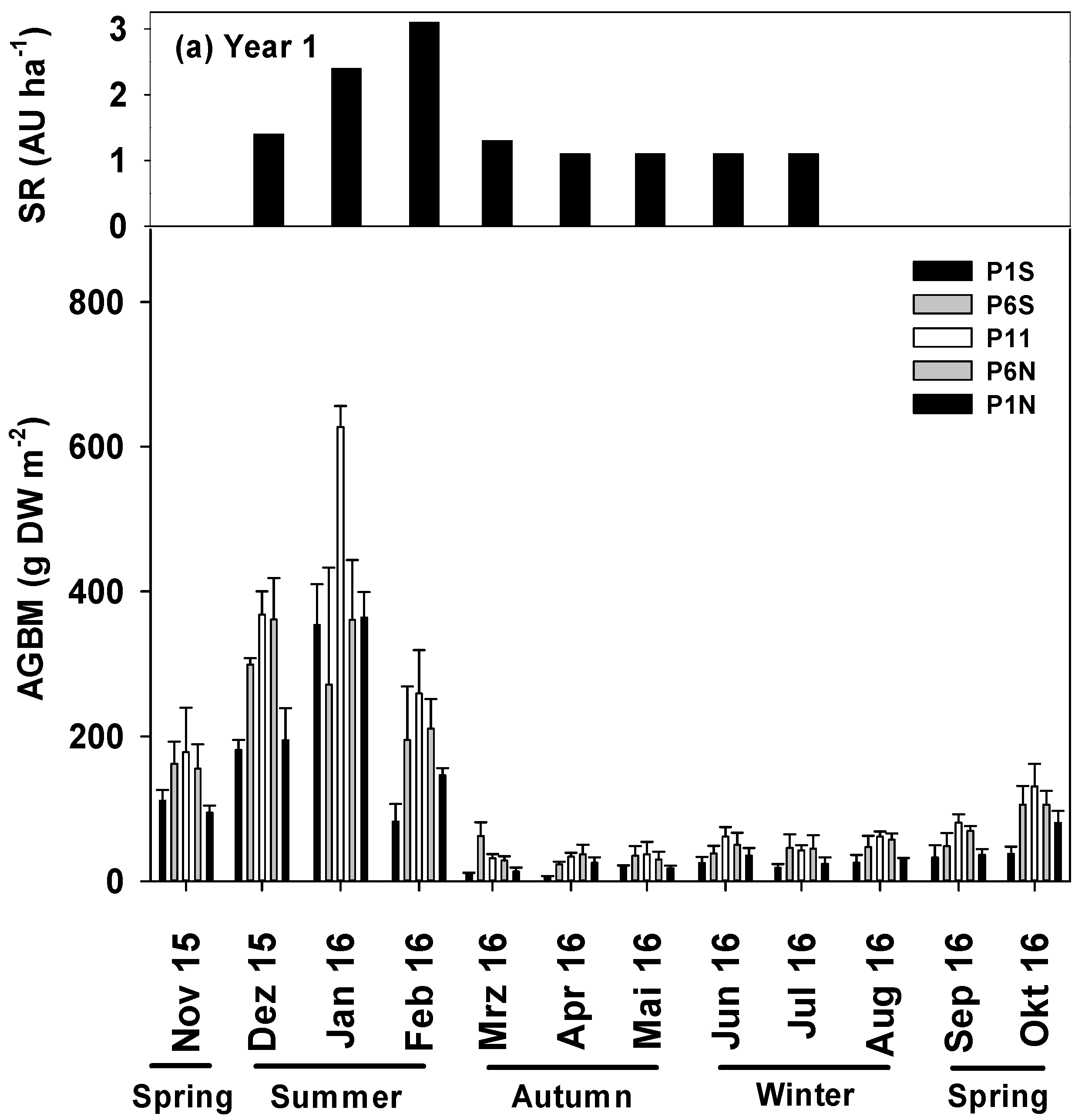
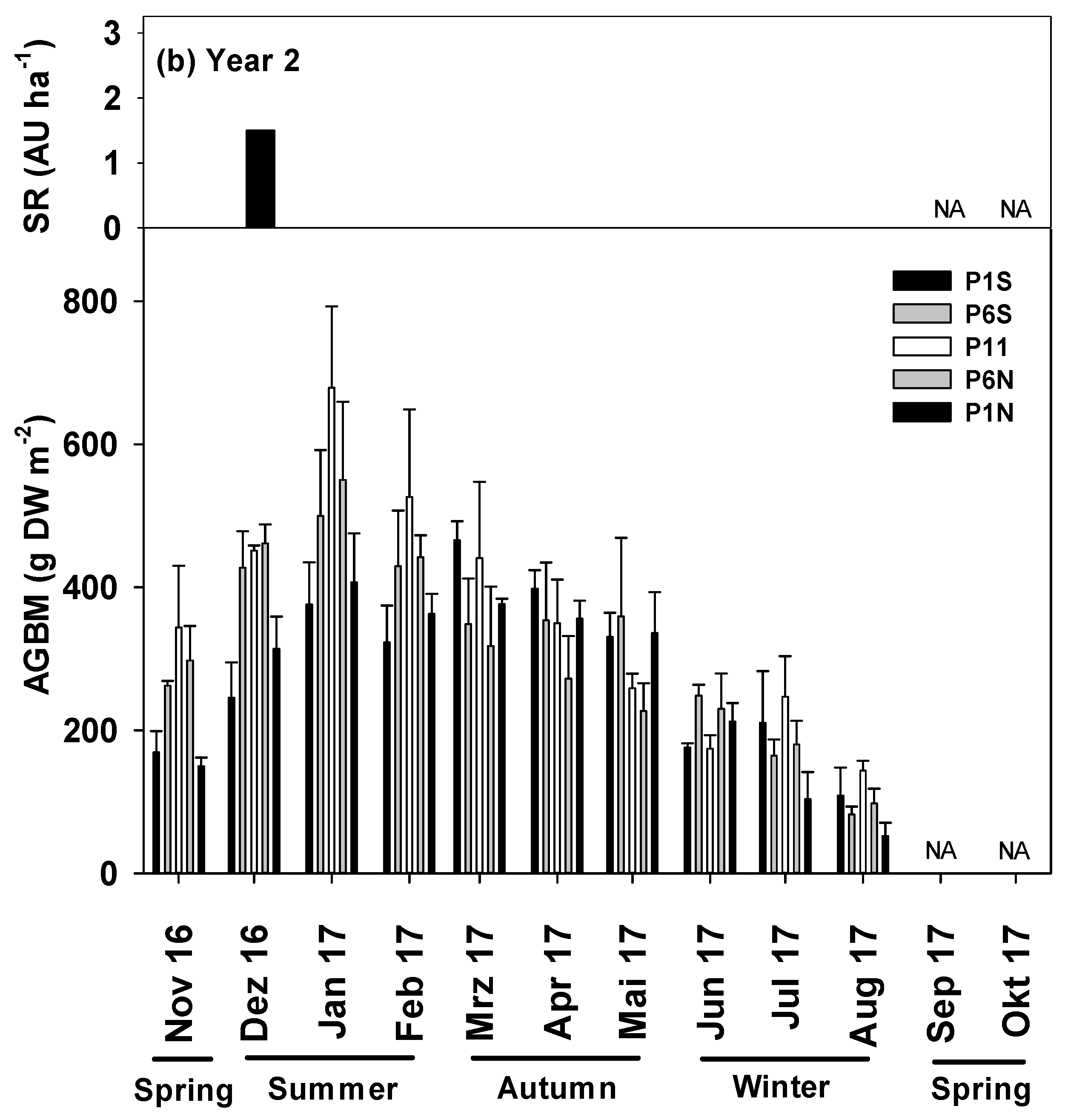
3.3. AGBM between the Tree Rows during Different Seasons
3.4. Correlation between PAR and AGBM and SM 0–100 cm and AGBM between the Tree Rows
4. Discussion
4.1. PAR, SM and AGBM Distribution throughout the Year
4.2. PAR, SM and AGBM Gradients between the Tree Rows
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oliveira, P.T.S.; Nearing, M.A.; Moran, M.S.; Goodrich, D.C.; Wendland, E.; Gupta, H.V. Trends in Water Balance Components across the Brazilian Cerrado. Water Resour. Res. 2014, 50, 7100–7114. [Google Scholar] [CrossRef] [Green Version]
- Sano, E.E.; Rosa, R.; Brito, J.L.S.; Ferreira, L.G. Land Cover Mapping of the Tropical Savanna Region in Brazil. Environ. Monit. Assess. 2010, 166, 113–124. [Google Scholar] [CrossRef]
- Santos, D.D.C.; Júnior, R.G.; Vilela, L.; Pulrolnik, K.; Bufon, V.B.; França, A.F.D.S. Forage dry mass accumulation and structural characteristics of Piatã grass in silvopastoral systems in the Brazilian savannah. Agric. Ecosyst. Environ. 2016, 233, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Macedo, M.C.M. Pastagens no ecossistema Cerrados: Evolução das pesquisas para o desenvolvimento sustentável. REUNIÃO Anu. Soc. Bras. Zootec. 2005, 42, 56–84. [Google Scholar]
- Macedo, M.C.M.; Zimmer, A.H.; Kichel, A.N.; de Almeida, R.G.; de Araujo, A.R. Degradação de pastagens, alternativas de recuperação e renovação, e formas de mitigação. In Encontro De Adubacao De Pastagens Da Scot Consultoria - Tec - Fertil; 2013; pp. 158–181. Available online: https://www.alice.cnptia.embrapa.br/bitstream/doc/976514/1/DegradacaopastagensalternativasrecuperacaoMMacedoScot.pdf (accessed on 10 March 2021).
- Balbino, L.C.; Cordeiro, L.A.M.; Porfírio-Da-Silva, V.; De Moraes, A.; Martínez, G.B.; Alvarenga, R.C.; Kichel, A.N.; Fontaneli, R.S.; Dos Santos, H.P.; Franchini, J.C.; et al. Evolução tecnológica e arranjos produtivos de sistemas de integração lavoura-pecuária-floresta no Brasil. Pesqui. Agropecuária Bras. 2011, 46. [Google Scholar] [CrossRef] [Green Version]
- Dias-Filho, M.B. Degradação de Pastagens: Processos, Causas e Estratégias de Recuperação; MBDF: Belém, Brazil, 2011. [Google Scholar]
- Alves, B.J.R.; Madari, B.E.; Boddey, R.M. Integrated Crop–Livestock–Forestry Systems: Prospects for a Sustainable Agricultural Intensification. Nutr. Cycl. Agroecosystems 2017, 108, 1–4. [Google Scholar] [CrossRef]
- de Moraes, A.; Carvalho, P.C.D.F.; Anghinoni, I.; Lustosa, S.B.C.; Costa, S.E.V.G.D.A.; Kunrath, T.R. Integrated crop–livestock systems in the Brazilian subtropics. Eur. J. Agron. 2014, 57, 4–9. [Google Scholar] [CrossRef]
- Lemaire, G.; Franzluebbers, A.; Carvalho, P.C.D.F.; Dedieu, B. Integrated crop–livestock systems: Strategies to achieve synergy between agricultural production and environmental quality. Agric. Ecosyst. Environ. 2014, 190, 4–8. [Google Scholar] [CrossRef]
- Bono, J.A.M.; Macedo, M.C.M.; Tormena, C.A.; Nanni, M.R.; Gomes, E.P.; Müller, M.M.L. Infiltração de Água No Solo Em Um Latossolo Vermelho Da Região Sudoeste Dos Cerrados Com Diferentes Sistemas de Uso e Manejo. Rev. Bras. Ciênc. Solo 2012, 36, 1845–1853. [Google Scholar] [CrossRef] [Green Version]
- Nair, P.R. An Introduction to Agroforestry; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1993. [Google Scholar]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; ISBN 978-1-107-05799-9. [Google Scholar]
- Pezzopane, J.R.M.; Bosi, C.; Nicodemo, M.L.F.; Santos, P.M.; Da Cruz, P.G.; Parmejiani, R.S. Microclimate and soil moisture in a silvopastoral system in southeastern Brazil. Bragantia 2015, 74, 110–119. [Google Scholar] [CrossRef] [Green Version]
- Bosi, C.; Pezzopane, J.R.M.; Sentelhas, P.C. Soil Water Availability in a Full Sun Pasture and in a Silvopastoral System with Eucalyptus. Agrofor. Syst. 2019. [Google Scholar] [CrossRef]
- Lin, B.B. The Role of Agroforestry in Reducing Water Loss through Soil Evaporation and Crop Transpiration in Coffee Agroecosystems. Agric. For. Meteorol. 2010, 150, 510–518. [Google Scholar] [CrossRef]
- Rodrigues, C.O.D.; Araújo, S.A.D.C.; Viana, M.C.M.; Rocha, N.S.; Braz, T.G.D.S.; Villela, S.D.J. Light relations and performance of signal grass in silvopastoral system. Acta Sci. Anim. Sci. 2014, 36, 129. [Google Scholar] [CrossRef]
- Burner, D.M.; Belesky, D.P. Relative Effects of Irrigation and Intense Shade on Productivity of Alley-Cropped Tall Fescue Herbage. Agrofor. Syst. 2008, 73, 127–139. [Google Scholar] [CrossRef]
- WRB. World Reference Base for Soil Resources 2014: International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; FAO: Rome, Italy, 2014; ISBN 978-92-5-108370-3. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 10 March 2021).
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means; 2019; R Package Version 1.3.4; Available online: https://CRAN.R-project.org/package=emmeans (accessed on 10 March 2021).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. {nlme}: Linear and Nonlinear Mixed Effects Models; 2018; R Package Version 3.1-137; Available online: https://CRAN.R-project.org/package=nlme (accessed on 10 March 2021).
- Feldhake, C.M. Microclimate of a Natural Pasture under Planted Robinia Pseudoacacia in Central Appalachia, West Virginia. Agrofor. Syst. 2001, 53, 297–303. [Google Scholar] [CrossRef]
- Guenni, O.; Marín, D.; Baruch, Z. Responses to Drought of Five Brachiaria Species. I. Biomass Production, Leaf Growth, Root Distribution, Water Use and Forage Quality. Plant. Soil 2002, 243, 229–241. [Google Scholar] [CrossRef]
- De Araujo, L.C.; Santos, P.M.; Mendonça, F.C.; Mourão, G.B. Establishment of Brachiaria Brizantha Cv. Marandu, under Levels of Soil Water Availability in Stages of Growth of the Plants. Rev. Bras. Zootec. 2011, 40, 1405–1411. [Google Scholar] [CrossRef] [Green Version]
- Pedreira, B.C.; Pedreira, C.G.S.; Boote, K.J.; Lara, M.A.S.; Alderman, P.D. Adapting the CROPGRO Perennial Forage Model to Predict Growth of Brachiaria Brizantha. Field Crops Res. 2011, 120, 370–379. [Google Scholar] [CrossRef]
- Do Valle, C.B.; Euclides, V.; Valerio, J.; Macedo, M.; Fernandes, C.; Dias Filho, M. Brachiaria brizanta cv. Piatã: Uma forrageira para a diversificação de pastagens tropicais. Seed News. 2007, 11, 28–30. [Google Scholar]
- De Araujo, L.C.; Santos, P.M.; Rodriguez, D.; Pezzopane, J.R.M. Key factors that influence for seasonal production of Guinea grass. Sci. Agric. 2018, 75, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Sarath, G.; Baird, L.M.; Mitchell, R.B. Senescence, Dormancy and Tillering in Perennial C4 Grasses. Plant. Sci. 2014, 217–218, 140–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, J.R.; Wild, D.W.M. Improvement of Nitrogen Nutrition and Grass Growth under Shading. Forages Plant. Crop. HM Shelton WW Stür ACIAR Proc. 1991, 32, 77. [Google Scholar]
- Paciullo, D.S.C.; De Carvalho, C.A.B.; Aroeira, L.J.M.; Morenz, M.J.F.; Lopes, F.C.F.; Rossiello, R.O.P. Morfofisiologia e valor nutritivo do capim-braquiária sob sombreamento natural e a sol pleno. Pesqui. Agropecuária Bras. 2007, 42, 573–579. [Google Scholar] [CrossRef] [Green Version]
- Soares, A.B.; Sartor, L.R.; Adami, P.F.; Varella, A.C.; Fonseca, L.; Mezzalira, J.C. Influência da luminosidade no comportamento de onze espécies forrageiras perenes de verão. Rev. Bras. Zootec. 2009, 38, 443–451. [Google Scholar] [CrossRef] [Green Version]
- Frelih-Larsen, A.; Hinzmann, M.; Ittner, S. The ‘Invisible’ Subsoil: An Exploratory View of Societal Acceptance of Subsoil Management in Germany. Sustainability 2018, 10, 3006. [Google Scholar] [CrossRef] [Green Version]
- Rosenbaum, U.; Bogena, H.R.; Herbst, M.; Huisman, J.A.; Peterson, T.J.; Weuthen, A.; Western, A.W.; Vereecken, H. Seasonal and Event Dynamics of Spatial Soil Moisture Patterns at the Small Catchment Scale. Water Resour. Res. 2012, 48. [Google Scholar] [CrossRef] [Green Version]
- Paciullo, D.S.C.; Fernandes, P.B.; Gomide, C.A.D.M.; De Castro, C.R.T.; Sobrinho, F.D.S.; De Carvalho, C.A.B. The growth dynamics in Brachiaria species according to nitrogen dose and shade. Rev. Bras. Zootec. 2011, 40, 270–276. [Google Scholar] [CrossRef] [Green Version]
- Paciullo, D.S.C.; Campos, N.R.; Gomide, C.A.M.; De Castro, C.R.T.; Tavela, R.C.; Rossiello, R.O.P. Crescimento de capim-braquiária influenciado pelo grau de sombreamento e pela estação do ano. Pesqui. Agropecuária Bras. 2008, 43, 917–923. [Google Scholar] [CrossRef] [Green Version]
- Bouillet, J.-P.; Laclau, J.-P.; Arnaud, M.; M’Bou, A.T.; Saint-André, L.; Jourdan, C. Changes with Age in the Spatial Distribution of Roots of Eucalyptus Clone in Congo: Impact on Water and Nutrient Uptake. For. Ecol. Manag. 2002, 171, 43–57. [Google Scholar] [CrossRef]
- Laclau, J.-P.; Arnaud, M.; Bouillet, J.-P.; Ranger, J. Spatial Distribution of Eucalyptus Roots in a Deep Sandy Soil in the Congo: Relationships with the Ability of the Stand to Take up Water and Nutrients. Tree Physiol. 2001, 21, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Way, D.A.; Katul, G.G.; Manzoni, S.; Vico, G. Increasing Water Use Efficiency along the C3 to C4 Evolutionary Pathway: A Stomatal Optimization Perspective. J. Exp. Bot. 2014, 65, 3683–3693. [Google Scholar] [CrossRef] [Green Version]
- Tian, S.; Cacho, J.F.; Youssef, M.A.; Chescheir, G.M.; Fischer, M.; Nettles, J.E.; King, J.S. Switchgrass Growth and Pine–Switchgrass Interactions in Established Intercropping Systems. GCB Bioenergy 2017, 9, 845–857. [Google Scholar] [CrossRef]
- Wilson, J.R. Influence of Planting Four Tree Species on the Yield and Soil Water Status of Green Panic Pasture in Subhumid South-East Queensland. Trop. Grassl. 1998, 32, 209–220. [Google Scholar]
- Fisher, M.J.; Kerridge, P.C. The Agronomy and Physiology of Brachiaria Species. In Brachiaria: Biology, Agronomy and Improvement; Miles, J.W., Ed.; CIAT Publication. No 259 CIAT; Tropical Forages and Communication Unit: Campo Grande, Brazil, 1996; pp. 43–52. [Google Scholar]
- Silva-Pando, F.J.; González-Hernández, M.P.; Rozados-Lorenzo, M.J. Pasture Production in a Silvopastoral System in Relation with Microclimate Variables in the Atlantic Coast of Spain. Agrofor. Syst. 2002, 56, 203–211. [Google Scholar] [CrossRef]
- De Oliveira, C.C.; Alves, F.V.; Martins, P.G.M.D.A.; Junior, N.K.; Alves, G.F.; De Almeida, R.G.; Mastelaro, A.P.; Silva, E.V.D.C.E. Vaginal temperature as indicative of thermoregulatory response in Nellore heifers under different microclimatic conditions. PLoS ONE 2019, 14, e0223190. [Google Scholar] [CrossRef] [Green Version]
- Franzluebbers, A.J.; Chappell, J.C.; Shi, W.; Cubbage, F.W. Greenhouse Gas Emissions in an Agroforestry System of the Southeastern USA. Nutr. Cycl. Agroecosystems 2017, 108, 85–100. [Google Scholar] [CrossRef]
- Sato, J.H.; De Carvalho, A.M.; De Figueiredo, C.C.; Coser, T.R.; De Sousa, T.R.; Vilela, L.; Marchão, R.L. Nitrous oxide fluxes in a Brazilian clayey oxisol after 24 years of integrated crop-livestock management. Nutr. Cycl. Agroecosystems 2017, 108, 55–68. [Google Scholar] [CrossRef]
- Sant’Anna, S.A.C.; Jantalia, C.P.; Sá, J.M.; Vilela, L.; Marchão, R.L.; Alves, B.J.R.; Urquiaga, S.; Boddey, R. Changes in Soil Organic Carbon during 22 Years of Pastures, Cropping or Integrated Crop/Livestock Systems in the Brazilian Cerrado. Nutr. Cycl. Agroecosystems 2016. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Moisture (Vol%) | SM = Y × M × SP + R | |||
|---|---|---|---|---|
| 0–100 cm | Df | SS | MS | p |
| Year (Y) | 1 | 5.0 | 5.0 | 0.39010 |
| Month (M) | 20 | 2329.7 | 116.5 | <0.001 *** |
| Sample point (SP) | 4 | 1766.9 | 441.7 | <0.001 *** |
| Replication (R) | 2 | 169.5 | 84.7 | <0.001 *** |
| Interaction (Y × SP) | 4 | 35.2 | 8.8 | 0.26710 |
| Interaction (M × SP) | 80 | 273.0 | 3.4 | 0.99970 |
| Residuals | 218 | 1463.7 | 6.7 | |
| Soil Moisture (Vol%) | SM = Y × S × SP + R | |||
|---|---|---|---|---|
| (a) Depth: 10 cm | Df | SS | MS | p |
| Year (Y) | 1 | 552 | 552 | <0.001 *** |
| Season (S) | 3 | 2965 | 988 | <0.001 *** |
| Sample point (SP) | 4 | 184 | 46 | <0.001 *** |
| Replication (R) | 2 | 4 | 2 | 0.8 |
| Interaction (Y × S) | 3 | 307 | 102 | <0.001 *** |
| Interaction (Y × SP) | 4 | 53 | 13 | 0.2 |
| Interaction (S × SP) | 12 | 91 | 8 | 0.5 |
| Interaction (Y × S × SP) | 12 | 42 | 4 | 0.9 |
| Residuals | 288 | 2248 | 8 | |
| (b) Depth: 20 cm | Df | SS | MS | p |
| Year (Y) | 1 | 133 | 133 | <0.001 *** |
| Season (S) | 3 | 1398 | 466 | <0.001 *** |
| Sample point (SP) | 4 | 2296 | 574 | <0.001 *** |
| Replication (R) | 2 | 39 | 19 | 0.9 |
| Interaction (Y × S) | 3 | 75 | 25 | < 0.05 * |
| Interaction (Y × SP) | 4 | 61 | 15 | 0.1 |
| Interaction (S × SP) | 12 | 32 | 3 | 1 |
| Interaction (Y × S × SP) | 12 | 36 | 3 | 1 |
| Residuals | 288 | 2275 | 8 | |
| (c) Depth: 30 cm | Df | SS | MS | p |
| Year (Y) | 1 | 172 | 172 | <0.001 *** |
| Season (S) | 3 | 1390 | 463 | <0.001 *** |
| Sample point (SP) | 4 | 3143 | 784 | <0.001 *** |
| Replication (R) | 2 | 111 | 56 | <0.01 ** |
| Interaction (Y × S) | 3 | 9 | 3 | 0.8 |
| Interaction (Y × SP) | 4 | 84 | 21 | 0.07 |
| Interaction (S × SP) | 12 | 42 | 3 | 1 |
| Interaction (Y × S × SP) | 12 | 10 | 1 | 1 |
| Residuals | 288 | 2794 | 10 | |
| (d) Depth: 40 cm | Df | SS | MS | p |
| Year (Y) | 1 | 311 | 311 | <0.001 *** |
| Season (S) | 3 | 1270 | 423 | <0.001 *** |
| Sample point (SP) | 4 | 2984 | 746 | <0.001 *** |
| Replication (R) | 2 | 351 | 176 | <0.001 *** |
| Interaction (Y × S) | 3 | 2 | 1 | 1 |
| Interaction (Y × SP) | 4 | 116 | 29 | <0.05 * |
| Interaction (S × SP) | 12 | 50 | 4 | 1 |
| Interaction (Y × S × SP) | 12 | 13 | 1 | 1 |
| Residuals | 288 | 3039 | 11 | |
| (e) Depth: 60 cm | Df | SS | MS | p |
| Year (Y) | 1 | 15 | 15 | 0.4 |
| Season (S) | 3 | 1405 | 468 | <0.001 *** |
| Sample point (SP) | 4 | 541 | 135 | <0.001 *** |
| Replication (R) | 2 | 145 | 72 | <0.05 * |
| Interaction (Y × S) | 3 | 42 | 14 | 0.5 |
| Interaction (Y × SP) | 4 | 78 | 19 | 0.4 |
| Interaction (S × SP) | 12 | 79 | 7 | 1 |
| Interaction (Y × S × SP) | 12 | 39 | 3 | 1 |
| Residuals | 288 | 5386 | 19 | |
| (f) Depth: 100 cm | Df | SS | MS | p |
| Year (Y) | 1 | 138 | 138 | <0.001 *** |
| Season (S) | 3 | 1165 | 388 | <0.001 *** |
| Sample point (SP) | 4 | 5089 | 1272 | <0.001 *** |
| Replication (R) | 2 | 438 | 219 | <0.001 *** |
| Interaction (Y × S) | 3 | 531 | 177 | <0.001 *** |
| Interaction (Y × SP) | 4 | 126 | 31 | <0.05 * |
| Interaction (S × SP) | 12 | 137 | 11 | 0.8 |
| Interaction (Y × S × SP) | 12 | 82 | 7 | 1 |
| Residuals | 288 | 2886 | 10 | |
| AGBM | BM = Y × M × SP + R | |||
|---|---|---|---|---|
| (g DW m−2) | Df | SS | MS | p |
| Year (Y) | 1 | 1,767,996 | 1,767,996 | <0.001 *** |
| Month (M) | 20 | 5,914,719 | 295,736 | <0.001 *** |
| Sample point (SP) | 4 | 298,911 | 74,728 | <0.001 *** |
| Replication | 2 | 136,707 | 68,353 | <0.001 *** |
| Interaction (Y × SP) | 4 | 6134 | 1533 | 0.882625 |
| Interaction (M × SP) | 80 | 653,342 | 8167 | <0.01 ** |
| Residuals | 218 | 1,142,667 | 5242 | |
| AGBM | |||||
|---|---|---|---|---|---|
| (g DW m−2) | Spring | Summer | Autumn | Winter | |
| Sample Points | Year 1 | Average | |||
| P1S | 111.0 ± 14.9 aAB 1 | 205.7 ± 43.7 bA | 11.7 ± 2.3 cB | 23.0 ± 4.3 cB | 83.2 ± 20.2b |
| P6S | 162.1 ± 30.5 aB | 290.3 ± 43.0 bA | 40.1 ± 8.9 aC | 43.6 ± 7.8 bC | 128.4 ± 24.5ab |
| P11 | 178.0 ± 61.7 aB | 418.1 ± 58.6 aA | 34.1 ± 5.5 aC | 55.0 ± 5.8 aC | 170.0 ± 35.8a |
| P6N | 155.1± 33.9 aB | 310.8 ± 40.1 abA | 31.7 ± 5.4 abC | 50.7 ± 7.9 abC | 133.5 ± 25.6ab |
| P1N | 94.6 ± 9.6 aB | 234.8 ± 37.0 bA | 18.7 ± 3.5 bcB | 29.8 ± 4.3 cB | 94.4 ± 20.6b |
| Average | 140.1 ± 15.8B | 291.9 ± 22.2A | 27.3 ± 2.9C | 40.4 ± 3.2C | |
| Sample Points | Year 2 | Average | |||
| P1S | 79.9 ± 24.6 bB | 329.1 ± 34.6 cA | 398.3 ± 24.2 aA | 165.3 ± 28.1 abB | 243.1 ± 25.3ab |
| P6S | 138.8 ± 33.3 abB | 452.3 ± 39.6 abA | 353.8 ± 43.5 abA | 165.1 ± 25.4 abB | 277.5 ± 28.0ab |
| P11 | 185.1 ± 48.4 aC | 552.2 ± 58.7 aA | 349.8 ± 44.5 abB | 188.4 ± 23.4 aC | 318.9 ± 33.5a |
| P6N | 157.6 ± 38.5 abC | 484.6 ± 37.5 aA | 272.4 ± 34.1 bB | 169.3 ± 26.5 abBC | 271.0 ± 27.6ab |
| P1N | 88.9 ± 17.7 abB | 361.3 ± 28.3 bcA | 356.3 ± 19.0 abA | 122.9 ± 27.6 bB | 232.4 ± 24.3b |
| Average | 130.0 ± 15.8C | 433.0 ± 21.4A | 346.1 ± 16.0B | 162.2 ± 11.6C | |
| Pearson Correlation Coefficient r | ||||
|---|---|---|---|---|
| Spring | Summer | Autumn | Winter | |
| PAR vs. AGBM | 0.90 *** | 0.94 *** | −0.12 | −0.44 |
| SM 0–100 cm vs. AGBM | 0.8 ** | 0.87 *** | 0.49 | 0.87 ** |
| N = 10 |
| PAR Reduction (%) | Season | ||||
|---|---|---|---|---|---|
| Sample Point | Spring | Summer | Autumn | Winter | Average |
| P1S | 71 | 90 | 33 | 43 | 59 |
| P6S | 26 | 57 | 33 | 74 | 47 |
| P11 | 9 | 21 | 56 | 59 | 36 |
| P6N | 39 | 47 | 78 | 60 | 56 |
| P1N | 83 | 89 | 42 | 54 | 67 |
| Average | 46 | 61 | 48 | 58 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glatzle, S.; Stuerz, S.; Giese, M.; Pereira, M.; de Almeida, R.G.; Bungenstab, D.J.; Macedo, M.C.M.; Asch, F. Seasonal Dynamics of Soil Moisture in an Integrated-Crop-Livestock-Forestry System in Central-West Brazil. Agriculture 2021, 11, 245. https://doi.org/10.3390/agriculture11030245
Glatzle S, Stuerz S, Giese M, Pereira M, de Almeida RG, Bungenstab DJ, Macedo MCM, Asch F. Seasonal Dynamics of Soil Moisture in an Integrated-Crop-Livestock-Forestry System in Central-West Brazil. Agriculture. 2021; 11(3):245. https://doi.org/10.3390/agriculture11030245
Chicago/Turabian StyleGlatzle, Sarah, Sabine Stuerz, Marcus Giese, Mariana Pereira, Roberto Giolo de Almeida, Davi José Bungenstab, Manuel Claudio M. Macedo, and Folkard Asch. 2021. "Seasonal Dynamics of Soil Moisture in an Integrated-Crop-Livestock-Forestry System in Central-West Brazil" Agriculture 11, no. 3: 245. https://doi.org/10.3390/agriculture11030245
APA StyleGlatzle, S., Stuerz, S., Giese, M., Pereira, M., de Almeida, R. G., Bungenstab, D. J., Macedo, M. C. M., & Asch, F. (2021). Seasonal Dynamics of Soil Moisture in an Integrated-Crop-Livestock-Forestry System in Central-West Brazil. Agriculture, 11(3), 245. https://doi.org/10.3390/agriculture11030245