
Grassland Restoration at a Graded Ski Slope: Effects of Propagation Material and Fertilisation on Plant Cover and Vegetation

Abstract
:1. Introduction
- To test different types of wild-harvested propagation material and compare them to seeds of forage cultivars for plant cover persistence and species composition. The hypothesis was that the cover obtained with different wild-harvested materials would have different species composition and persist longer than those generated by forage cultivars.
- To test different fertilisation timing and frequency to improve the plant cover. The hypothesis was that more frequent fertilisation would accelerate the establishment of plant cover and the induced higher cover would persist even after the fertilisation cessation.
2. Materials and Methods
2.1. The Experimental Site
2.2. The Experiment
2.3. Univariate Analyses on Ground Cover Traits, Species Transfer, and Richness
2.4. Multivariate Analyses on Vegetation
3. Results
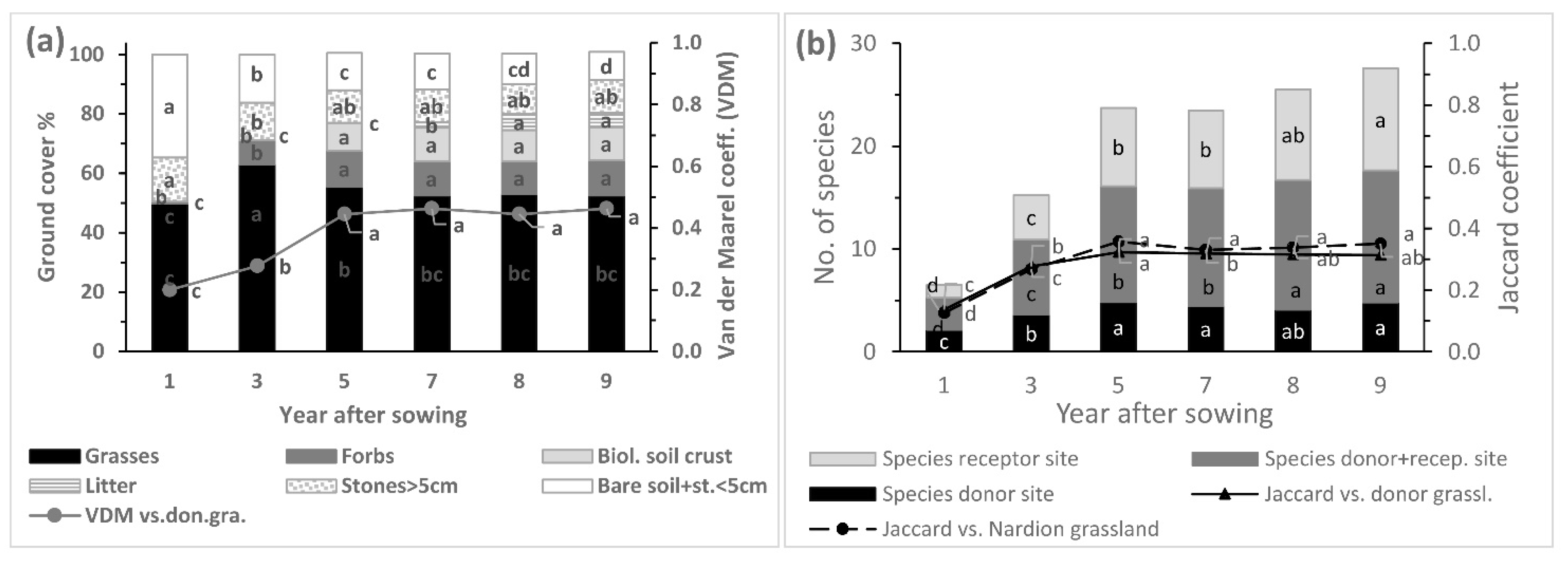
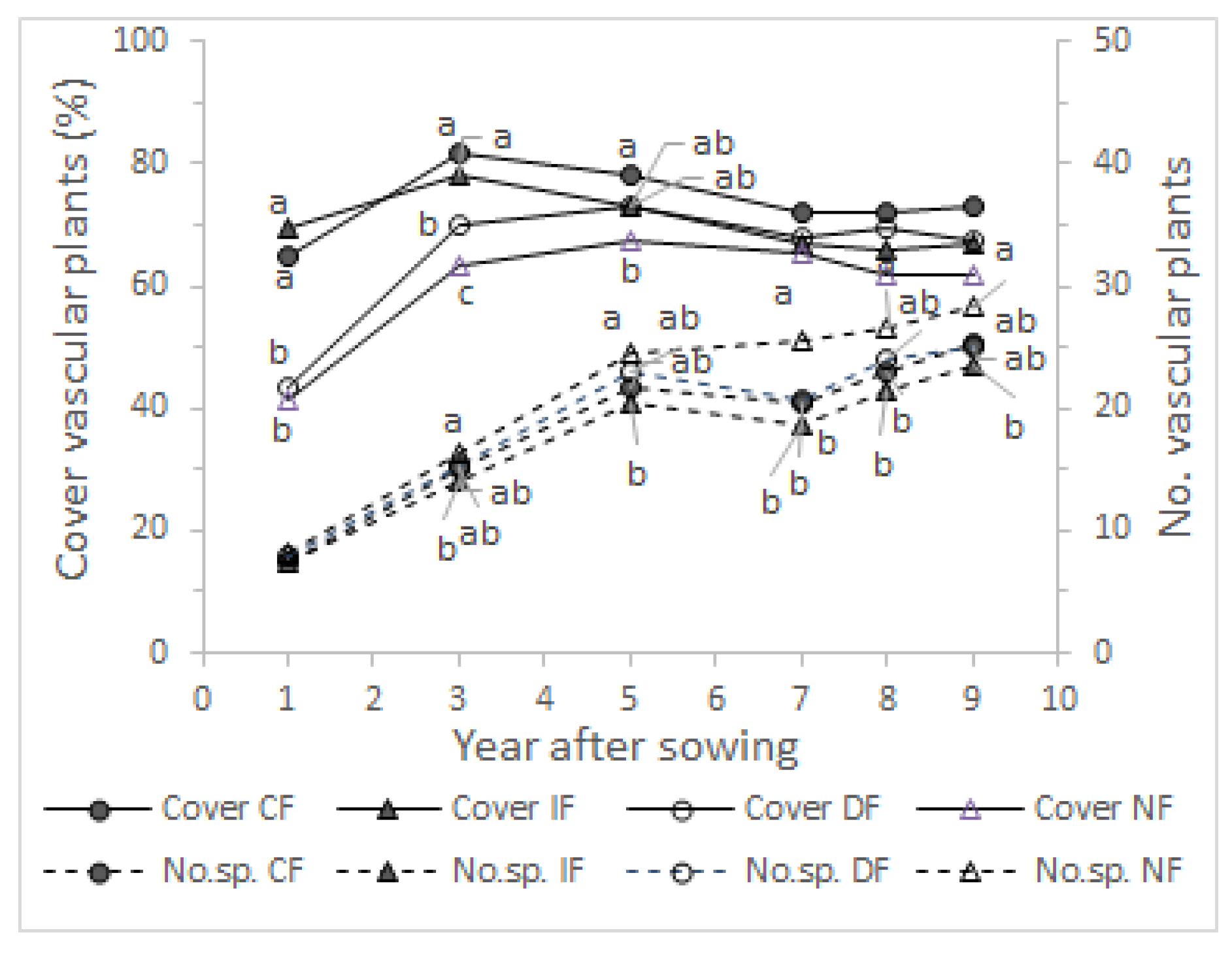
3.1. Ground Cover
3.2. Species Number
3.3. Species Transfer
3.4. Species Composition
3.5. Vegetation Structure
4. Discussion
4.1. Ground Cover
4.2. Species Number
4.3. Species Transfer
4.4. Species Composition and Vegetation Structure
5. Practical Implications for Ski Slope Revegetation
- Using propagation material from (semi-)natural grasslands with climate and soil similar to the restoration site: any material type performs well.
- If native propagation material is unavailable, adopting seed mixtures of short-lived cultivars can create an initial but non-persistent plant cover.
- Retaining nearby pre-existing vegetation as intact as possible, as it is an efficient seed source for colonising native species.
- Limiting fertilisation to the first year, as it reduces the number of colonising species and has a transient effect on the plant cover. The anti-erosion effect of the rapid achievement of a full plant cover aimed at with fertilisation can be replaced by organic mulching. If necessary, only phosphorous and potassium can be applied, as they are not leached and favour the establishment of N-fixing species, which can naturally increase the soil nitrogen fertility.
6. Conclusions
Supplementary Materials
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vanat, L. International Report on Snow & Mountain Tourism: Overview of the Key Industry Figures for Ski Resorts, 12th edition, 2020. Available online: http://www.vanat.ch/RM-world-report-2020.pdf (accessed on 13 February 2021).
- Felber, H.U.; Hirsch, M.; Walther, P. Landschaftseingrifffe für den Skisport: Wegleitung zur Berücksichtigung des Natur- und Landschaftsschutzes; Eidgenossische Departement des Innern: Bern, Switzerland, 1991.
- Pintaldi, E.; Hudek, C.; Stanchi, S.; Spiegelberger, T.; Rivella, E.; Freppaz, M. Sustainable soil management in ski areas: Threats and challenges. Sustainability 2017, 9, 2150. [Google Scholar] [CrossRef] [Green Version]
- Burt, J.W.; Rice, K.J. Not all ski slopes are created equal: Disturbance intensity affects ecosystem properties. Ecol. Appl. 2009, 19, 2242–2253. [Google Scholar] [CrossRef]
- Forbes, B.C.; Jefferies, R.L. Revegetation of disturbed arctic sites: Constraints and applications. Biol. Conserv. 1999, 88, 15–24. [Google Scholar] [CrossRef]
- Barni, E.; Freppaz, M.; Siniscalco, C. Interactions between Vegetation, Roots, and Soil Stability in Restored High-altitude Ski Runs in the Alps. Arct. Antarct. Alp. Res. 2007, 39, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Urbanska, K.M. Restoration ecology research above the timberline: Colonization of safety islands on a machine-graded alpine ski run. Biodivers. Conserv. 1997, 6, 1655–1670. [Google Scholar] [CrossRef]
- Rydgren, K.; Auestad, I.; Hamre, L.N.; Hagen, D.; Rosef, L.; Skjerdal, G. Long-term persistence of seeded grass species: An unwanted side effect of ecological restoration. Environ. Sci. Pollut. Res. 2016, 23, 13591–13597. [Google Scholar] [CrossRef]
- Delarze, R. Dynamique de la vegetation sur les pistes ensemencees de Crans-Montana (Valais, Suisse). Effets de l’altitude. Botanica Helvetica 1994, 104, 3–16. [Google Scholar]
- Gretarsdottir, J.; Aradottir, A.L.; Vandvik, V.; Heegaard, E.; Birks, H.J.B. Long-term effects of reclamation treatments on plant succession in Iceland. Restor. Ecol. 2004, 12, 268–278. [Google Scholar] [CrossRef]
- Ash, H.J.; Gemmell, R.P.; Bradshaw, A.D. The introduction of native plant species on industrial waste heaps: A test of immigration and other factors affecting primary succession. J. Appl. Ecol. 1994, 31, 74–84. [Google Scholar] [CrossRef]
- Krautzer, B.; Wittmann, H.; Peratoner, G.; Graiss, W.; Partl, C.; Parente, G.; Venerus, S.; Rixen, C.; Streit, M. Site-Specific High Zone Restoration in the Alpine Region: The Current Technological Development; HBLFA Raumberg-Gumpenstein: Irdning, Austria, 2006. [Google Scholar]
- Argenti, G.; Ferrari, L. Plant cover evolution and naturalisation of revegetated ski runs in an Apennine ski resort (Italy). iForest 2009, 2, 178–182. [Google Scholar] [CrossRef] [Green Version]
- Roux-Fouillet, P.; Wipf, S.; Rixen, C. Long-term impacts of ski piste management on alpine vegetation and soils. J. Appl. Ecol. 2011, 48, 906–915. [Google Scholar] [CrossRef]
- Rydgren, K.; Halvorsen, R.; Odland, A.; Skjerdal, G. Restoration of alpine spoil heaps: Successional rates predict vegetation recovery in 50 years. Ecol. Eng. 2011, 37, 294–301. [Google Scholar] [CrossRef]
- Skeffington, R.A.; Bradshaw, A.D. Nitrogen fixation by plants growing on reclaimed China clay wastes. J. Appl. Ecol. 1980, 17, 469–477. [Google Scholar] [CrossRef]
- Hudek, C.; Barni, E.; Stanchi, S.; D’Amico, M.; Pintaldi, E.; Freppaz, M. Mid and long-term ecological impacts of ski run construction: On alpine ecosystems. Sci. Rep. 2020, 10, 11654. [Google Scholar] [CrossRef] [PubMed]
- OFP (Office Federal des Forets et de la Protection du Paysage). Directives Concernant les Atteintes Portees au Paysage dans l’Interet du Ski; OFCIM: Bern, Switzerland, 1979.
- Feucht, B.; Rieger, E.; Tamegger, C.; Jahn, F.; Jongepierová, I. Agricultural production of seeds from regional provenance. In Practical Handbook for Seed Harvest and Ecological Restoration of Species-Rich Grasslands; Scotton, M., Kirmer, A., Krautzer, B., Eds.; CLEUP: Padova, Italy, 2012; pp. 33–38. [Google Scholar]
- Scotton, M.; Rieger, E.; Feucht, B.; Tamegger, C.; Jahn, F.; Ševčíková, M.; Semanová, I.; Krautzer, B.; Graiss, W.; Haslgrübler, P.; et al. Techniques for harvesting seeds and plant material in species-rich grasslands. In Practical Handbook for Seed Harvest and Ecological Restoration of Species-Rich Grasslands; Scotton, M., Kirmer, A., Krautzer, B., Eds.; CLEUP: Padova, Italy, 2012. [Google Scholar]
- Malaval, S.; Dupin, B.; Dantin, G. Conservation et restauration de la flore dans un contexte anthropisé, quelles solutions? Sci. Eaux Territ. 2015, 16, 70–75. [Google Scholar] [CrossRef]
- Scotton, M. Establishing a semi-natural grassland: Effects of harvesting time and sowing density on species composition and structure of a restored Arrhenatherum elatius meadow. Agric. Ecosyst. Environ. 2016, 220, 35–44. [Google Scholar] [CrossRef]
- Locher, O.N.; Streit, M.; Frei, M.; Andrey, C.; Blaser, R.; Meyer, J.; Muller, U.; Reidy, B.; Rixen, C.; Schutz, M. Linee Guida per Il Rinverdimento ad Alta Quota; Verein für Ingenieurbiologie: Wädenswil, Switzerland, 2008. [Google Scholar]
- Rubel, F.; Brugger, K.; Haslinger, K.; Auer, I. The climate of the European Alps: Shift of very high resolution Köppen-Geiger climate zones 1800–2100. Climatology 2017, 26, 115–125. [Google Scholar] [CrossRef]
- Theurillat, J.-P.; Aeschimann, D.; Küpfer, P.; Spichiger, R. The higher vegetation units of the Alps. Colloques Phytosociologiques 1995, 23, 189–239. [Google Scholar]
- Pignatti, S. Flora d’Italia; Edagricole: Bologna, Italy, 1982. [Google Scholar]
- Scotton, M.; Piccinin, L.; Dainese, M.; Sancin, F. Seed harvesting for ecological restoration: Efficiency of haymaking and seed-stripping on different grassland types in the eastern Italian Alps. Ecol. Restor. 2009, 27, 66–75. [Google Scholar] [CrossRef]
- Schiechtl, H.M. Bioengineering for Land Reclamation and Conservation; University of Alberta Press: Edmonton, AB, Canada, 1980. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier Science BV: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Van der Maarel, E. Transformation of cover-abundance values in phytosociology and its effects on community similarity. Vegetatio 1979, 39, 97–114. [Google Scholar]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Kiehl, K.; Kirmer, A.; Donath, T.W.; Rasran, L.; Hölzel, N. Species introduction in restoration projects—Evaluation of different techniques for the establishment of semi-natural grasslands in Central and Northwestern Europe. Basic Appl. Ecol. 2010, 11, 285–299. [Google Scholar] [CrossRef]
- Rosenweig, M. Species Diversity in Space and Time; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- SAS Institute Inc. SAS/STAT® 13.2.; SAS Institute Inc.: Cary, NC, USA, 2002–2012. [Google Scholar]
- Leps, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Wildi, O.; Orloci, L. Numerical Exploration of Community Patterns; SPB Academic Publishing: The Hague, The Netherlands, 1990. [Google Scholar]
- Mosimann, T. Das Stabilitätspotential alpiner Geoökosysteme gegenüber Bodenstörungen durch Skipistenbau. Verhandlungen Gesellschaft Ökologie 1984, 12, 167–176. [Google Scholar]
- Belnap, J.; Kaltenecker, J.H.; Rosentreter, R.; Williams, J.; Leonard, S.; Eldridge, D. Biological Soil Crusts: Ecology and Management; United States Department of the Interior, Bureau of Land Management: Denver, CO, USA, 2001.
- Elmarsdottir, A.; Aradottir, A.L.; Trlica, M.J. Microsite availability and establishment of native species on degraded and reclaimed sites. J. Appl. Ecol. 2003, 40, 815–823. [Google Scholar] [CrossRef]
- Petersen, S.L.; Roundy, B.A.; Bryant, R.M. Revegetation Methods for High-Elevation Roadsides at Bryce Canyon National Park, Utah. Restor. Ecol. 2004, 12, 248–257. [Google Scholar] [CrossRef]
- Bradshaw, A. Restoration of mined lands—Using natural processes. Ecol. Eng. 1997, 8, 255–269. [Google Scholar] [CrossRef]
- Tinsley, M.J.; Simmons, M.T.; Windhager, S. The establishment of native versus non-native herbaceous seed mixes on a revegetated roadside in Central Texas. Ecol. Eng. 2006, 26, 231–240. [Google Scholar] [CrossRef] [Green Version]
- Scotton, M. Calcareous grassland restoration at a coarse quarry waste dump in the Italian Alps. Ecol. Eng. 2018, 117, 174–181. [Google Scholar] [CrossRef]
- Gentili, R.; Casati, E.; Ferrario, A.; Monti, A.; Montagnani, C.; Caronni, S.; Citterio, S. Vegetation cover and biodiversity levels are driven by backfilling material in quarry restoration. Catena 2020, 195, 104839. [Google Scholar] [CrossRef]
- Klug-Pümpel, B.; Krampitz, C. Conservation in Alpine Ecosystems: The plant cover of ski runs reflects natura1 as well as anthropogenic environmental factors. Die Bodenkultur 1996, 47, 97–117. [Google Scholar]
- Conrad, M.K.; Tischew, S. Grassland restoration in practice: Do we achieve the targets? A case study from Saxony-Anhalt/Germany. Ecol. Eng. 2011, 37, 1149–1157. [Google Scholar] [CrossRef]
- Brenner, F.J.; Werner, M.; Pike, J. Ecosystem development and natural succession in surface coal mine reclamation. Min. Environ. 1984, 6, 10–22. [Google Scholar] [CrossRef]
- Baasch, A.; Engst, K.; Schmiede, R.; May, K.; Tischew, S. Enhancing success in grassland restoration by adding regionally propagated target species. Ecol. Eng. 2016, 94, 583–591. [Google Scholar] [CrossRef]
- Řehounková, K.; Prach, K. Spontaneous vegetation succession in gravel-sand pits: A potential for restoration. Restor. Ecol. 2007, 16, 305–312. [Google Scholar] [CrossRef]
- Grime, J.P. Competitive exclusion m herbaceous vegetation. Nature 1973, 242, 344–347. [Google Scholar] [CrossRef]
- Callaway, R.M.; Brooker, R.W.; Choler, P.; Kikvidze, Z.; Lortie, C.J.; Michalet, R.; Paolini, L.; Pugnaire, F.L.; Newingham, B.; Aschehoug, E.T.; et al. Positive interactions among alpine plants increase with stress. Nature 2002, 417, 844–848. [Google Scholar] [CrossRef]
- Pywell, R.F.; Bullock, J.M.; Roy, D.B.; Warman, L.; Walker, K.J.; Rothery, P. Plant traits as predictors of performance in ecological restoration. J. Appl. Ecol. 2003, 40, 65–77. [Google Scholar] [CrossRef]
- Thormann, A.; Kiehl, K.; Pfadenhauer, J. Einfluss unterschiedlicher Renaturierungsmaßnahmen auf die langfristige Vegetationsentwicklung neu angelegter Kalkmagerrasen. Angewandte Landschaftsökologie 2003, 55, 73–106. [Google Scholar]
- Landolt, E.; Bäumler, B.; Erhardt, A.; Hegg, O.; Klötzli, F.; Lämmler, W.; Nobis, M.; Rudmann-Maurer, K.; Schweingruber, F.H.; Theurillat, J.-P.; et al. Flora indicativa. Ökologische Zeigerwerte und Biologische Kennzeichen zur Flora der Schweiz und der Alpen. Ecological Indicator Values and Biological Attributes of the Flora of Switzerland and the Alps; Haupt Verlag AG.: Bern, Switzerland, 2010. [Google Scholar]
- Hagen, D.; Hansen, T.I.; Graae, B.J.; Rydgren, K. To seed or not to seed in alpine restoration: Introduced grass species outcompete rather than facilitate native species. Ecol. Eng. 2014, 64, 255–261. [Google Scholar] [CrossRef]
- Isselin-Nondedeu, F.; Bédécarrats, A. Assessing the dominance of Phleum pratense cv. climax, a species commonly used for ski trail restoration. Appl. Veg. Sci. 2009, 12, 155–165. [Google Scholar] [CrossRef]
- Höglind, M.; Schapendonk, A.H.C.M.; van Oijen, M. Timothy growth in Scandinavia: Combining quantitative information and simulation modelling. New Phytol. 2001, 151, 355–367. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Factor | ||||
|---|---|---|---|---|
| Propagation Material | Fertilisation 4 | |||
| Level (code) | Area ratio 3 | Sowing rate (no seed m−2) | Level (code) | Fertilisation year after sowing year |
| Green hay, high rate (G) 1 | 2:1 | 12,414 | Continuous fertilisation (CF) | 1, 2, 3, and 4 |
| Green hay, low rate (g) 1 | 1:1 | 6207 | Initial fertilisation (IF) | 1, and 2 |
| Seed-stripped hay flower (S) 1 | 2:1 | 10,511 | Delayed fertilisation (DF) | 3, and 4 |
| Dray hay (D) 1 | 2:1 | 6932 | No fertilisation (NF) | - |
| Mixture forage cultivars (C) 2 | - | 40,777 | - | - |
| No sowing (N) | - | 0 | - | - |
| Treatment Means | Factor Main and Interaction Effect 3 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Propagation Material 1 | G | S | g | D | N | C | Mean | Mean | PM | F | PMxF | Y | PMxY | F x Y |
| Fertilisation 2 | Mean | Mean | Mean | Mean | Mean | Mean | CF | NF | ||||||
| Cover stones diameter > 5 cm (%) | 13.1 | 15.2 | 10.8 | 11.3 | 15.8 | 13.7 | 11.9b | 14.8a | ns | *** | ** | * | *** | ns |
| Cover fine earth + stones diam. < 5 cm (%) | 13.20 | 13.10 | 17.20 | 13.30 | 23.50 | 16.90 | 12.2b | 19.a | ns | *** | *** | *** | * | *** |
| Cover litter (%) | 2.97 | 2.12 | 0.35 | 1.79 | 0.30 | 0.38 | 1.8 | 2.2 | ns | ns | * | *** | *** | ns |
| Cover biological soil crust (%) | 5.7 | 5.6 | 5.5 | 6.1 | 9.6 | 9.2 | 6.46b | 7.40a | ns | *** | *** | *** | *** | ns |
| Cover vascular plants (%) | 65.a | 64.a | 66.1a | 67.4a | 50.9b | 59.8ab | 67.7a | 56.7b | ** | *** | *** | *** | *** | *** |
| Cover grasses (%) | 56.1a | 56.8a | 56.1a | 58.3a | 41.6b | 50.3a | 58.6a | 47.7b | *** | *** | *** | *** | *** | *** |
| Cover forbs (%) | 8.9 | 7.2 | 10.0 | 9.2 | 9.4 | 9.5 | 9.1 | 9.0 | ns | ns | *** | *** | ** | ns |
| No. species total | 18.7 | 21.0 | 19.3 | 21.9 | 23.0 | 21.2 | 20.1b | 21.6a | ns | *** | *** | *** | *** | ns |
| No. species from donor grassland | 4.3abc | 3.7abc | 4.4ab | 5.0a | 3.7abc | 2.8c | 3.9 | 4.1 | * | ns | * | *** | ns | ns |
| No. sp. from donor and Nardion grassl. | 10.2a | 10.7a | 10.5a | 11.4a | 9.1a | 6.8b | 9.2b | 10.3a | *** | *** | *** | *** | ns | ns |
| No. species from surrounding | 4.2c | 6.7abc | 4.3c | 5.5abc | 10.1a | 8.7ab | 6.4 | 6.7 | ** | ns | *** | *** | *** | ns |
| No. species from comm. seed mixture | 0b | 0b | 0b | 0b | 0.14b | 2.89a | 0.53 | 0.49 | *** | ns | * | ** | *** | * |
| Van der Maarel coeff. vs. donor grassl. | 0.5 | 0.4 | 0.3 | 0.5 | 0.3 | 0.3 | 0.4 | 0.4 | ns | ns | 0.001 | *** | *** | ns |
| Jaccard coeff. vs. donor grassland | 0.29b | 0.29b | 0.30ab | 0.34a | 0.24c | 0.22c | 0.28 | 0.28 | *** | ns | *** | *** | ns | ns |
| Jaccard coeff. vs. Nardion grassland | 0.23 | 0.34 | 0.23 | 0.29 | 0.33 | 0.35 | 0.29 | 0.30 | ns | ns | ** | *** | *** | ns |
| Analysis | Explanatory Variable | Covariable | Trace | Ftrace | p |
|---|---|---|---|---|---|
| Treatment | Year, Block | 0.183 | 5.3 | 0.001 | |
| Species composition | Propagation material | Year, Fertilisation, Block | 0.142 | 8.8 | 0.001 |
| (presence-absence) | Fertilisation | Year, PropMaterial, Block | 0.009 | 2.9 | 0.026 |
| Year | Plot, Block | 0.168 | 12.7 | 0.001 | |
| Treatment × Year | Year, Plot, Block | 0.165 | 1.2 | 0.001 | |
| Treatment | Year, Block | 0.213 | 8.4 | 0.001 | |
| Vegetation structure | Propagation material | Year, Fertilisation, Block | 0.185 | 15.7 | 0.001 |
| (% cover) | Fertilisation | Year, PropMaterial, Block | 0.007 | 3.0 | 0.1708 |
| Year | Plot, Block | 0.28 | 30.9 | 0.001 | |
| Treatment × Year | Year, Plot, Block | 0.151 | 2.0 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scotton, M. Grassland Restoration at a Graded Ski Slope: Effects of Propagation Material and Fertilisation on Plant Cover and Vegetation. Agriculture 2021, 11, 381. https://doi.org/10.3390/agriculture11050381
Scotton M. Grassland Restoration at a Graded Ski Slope: Effects of Propagation Material and Fertilisation on Plant Cover and Vegetation. Agriculture. 2021; 11(5):381. https://doi.org/10.3390/agriculture11050381
Chicago/Turabian StyleScotton, Michele. 2021. "Grassland Restoration at a Graded Ski Slope: Effects of Propagation Material and Fertilisation on Plant Cover and Vegetation" Agriculture 11, no. 5: 381. https://doi.org/10.3390/agriculture11050381
APA StyleScotton, M. (2021). Grassland Restoration at a Graded Ski Slope: Effects of Propagation Material and Fertilisation on Plant Cover and Vegetation. Agriculture, 11(5), 381. https://doi.org/10.3390/agriculture11050381