
Drought and Saline Stress Tolerance Induced in Somatic Hybrids of Solanum chacoense and Potato Cultivars by Using Mismatch Repair Deficiency

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. In Vitro Drought and Saline Stress
2.3. Plant Cultivation and Ex Vitro Drought Stress Treatment
2.4. Determination of Photosynthesis Efficiency
2.5. Statistical Analysis
3. Results and Discussion
3.1. In Vitro Drought-Stress Selection
3.2. In Vitro Salinity-Stress Selection
3.3. Plants Biomass Accumulation under Drought Condition
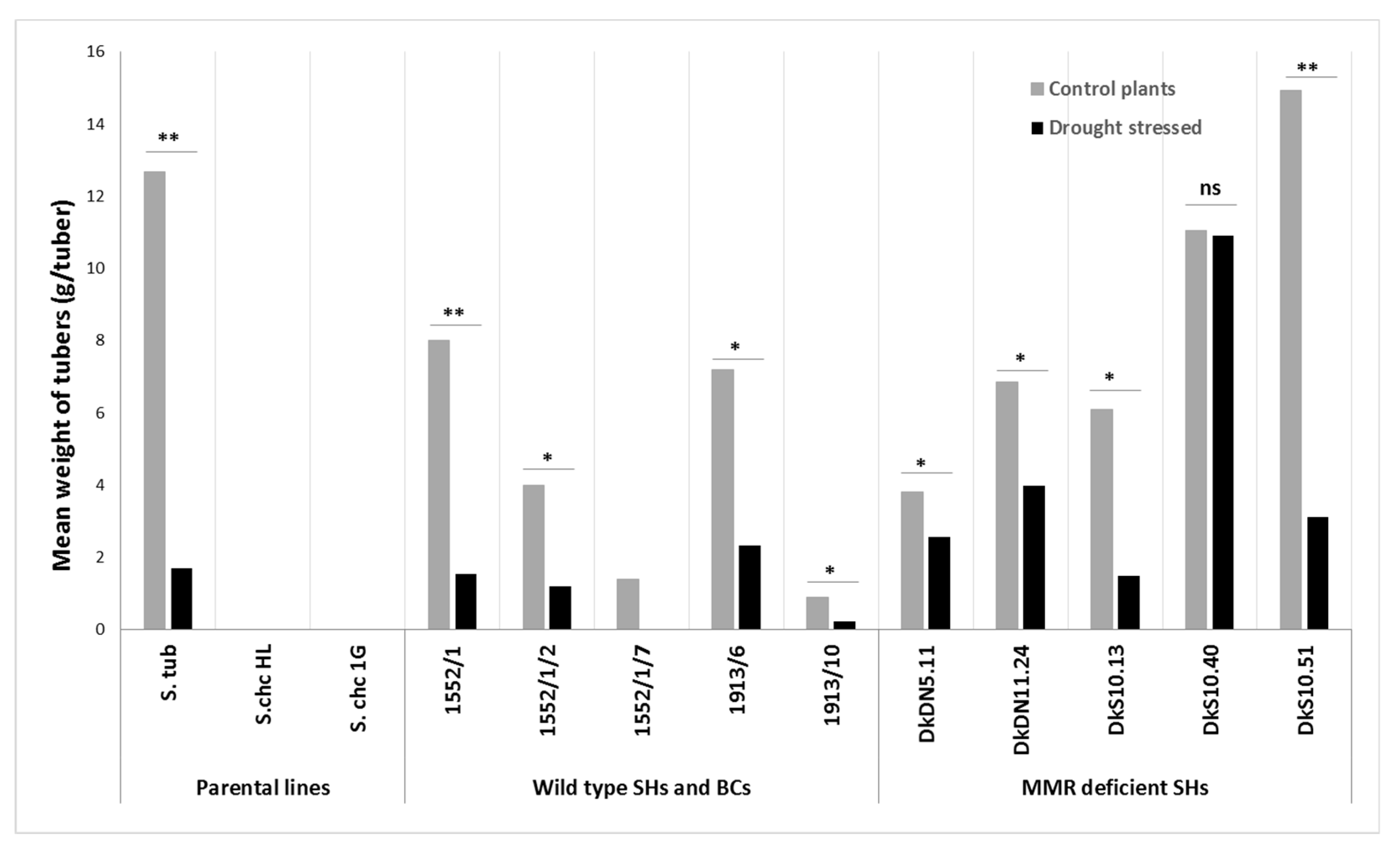
3.4. Drought Stress Effects on Tuber Production
3.5. Drought Stress Effects on Photosynthesis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ridoutt, B.G.; Pfister, S. A revised approach to water footprinting to make transparent the impacts of consumption and production on global freshwater scarcity. Glob. Environ. Chang. 2010, 20, 113–120. [Google Scholar] [CrossRef]
- FAO. “The State of the World’s Land and Water Resources for Food and Agriculture,” Food and Agriculture Organization of the United Nations and Earthscan, 2011. [Online]. Available online: http://www.fao.org/3/i1688e/i1688e.pdf (accessed on 15 May 2021).
- Meier, J.; Zabel, F.; Mauser, W. A global approach to estimate irrigated areas—A comparison between different data and statistics. Hydrol. Earth Syst. Sci. 2018, 22, 1119–1133. [Google Scholar] [CrossRef] [Green Version]
- Jaarsma, R.; de Vries, R.S.M.; de Boer, A.H. Effect of salt stress on Growth, Na+ accumulation and proline metabolism in potato (Solanum tuberosum) cultivars. PLoS ONE 2013, 8, e60183. [Google Scholar] [CrossRef]
- Ahmed, H.A.A.; Şahin, N.K.; Akdoğan, G.; Yaman, C.; Köm, D.; Uranbey, S. Variability in salinity stress tolerance of potato (Solanum tuberosum L.) varieties using in vitro screening. Ciênc. Agrotecnol. 2020, 44, e004220. [Google Scholar] [CrossRef]
- Chaves, M.M.; Fexas, J.; Pinhero, C. Photosynthesis under drought and salt stress: Regulation mechanisms. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anjum, S.A.; Xie, X.-Y.; Wang, L.-C.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Bray, E.A. Plant responses to water deficit. Trends Plant Sci. 1997, 2, 48–54. [Google Scholar] [CrossRef]
- Hu, Y.; Schmidhalter, U. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant Nutr. Soil Sci. 2005, 168, 541–549. [Google Scholar] [CrossRef]
- Smirnoff, N. Plant resistance to environmental stress. Curr. Opin. Biotechnol. 1998, 9, 214–219. [Google Scholar] [CrossRef]
- Rucker, K.S.; Kevin, C.K.; Holbrook, C.C.; Hook, J.E. Identification of peanut genotypes with improved drought avoidance traits. Peanut Sci. 1995, 22, 14–18. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Khan, A.L.; Muneer, S.; Kim, Y.-H.; Al-Rawahi, A.; Al-Harrasi, A. Silicon and salinity: Crosstalk in crop-mediated stress tolerance mechanisms. Front. Plant Sci. 2019, 10, 1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, S.M.; Doherty, S.J.; Croy, R.R.D. Biphasic superoxide generation in potato tubers. A Self-Amplifying Response Stress. Plant Physiol. 2003, 3, 1440–1449. [Google Scholar]
- Reddy, A.R.; Chiatanya, K.V.; Vivekanandan, M. Drought induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef] [PubMed]
- Tamura, T.; Hara, K.; Yamaguchi, Y.; Koizumi, N.; Sano, H. Osmotic stress tolerance of transgenic tobacco expressing a gene encoding a membrane-located receptor-like protein from tobacco plants. Plant Physiol. 2003, 131, 454–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalla Costa, L.; Delle Vedove, G.; Gianquintoi, G.; Giovanardi, R.; Peressotti, A. Yield, water use efficiency and nitrogen uptake in potato: Influence of drought stress. Potato Res. 1997, 40, 19–34. [Google Scholar] [CrossRef]
- Jefferies, R.A. Responses of potato genotypes to drought. I. Expansion of individual leaves and osmotic adjustment. Ann. Appl. Biol. 1993, 122, 93–104. [Google Scholar] [CrossRef]
- Shalhevet, J.; Hsiao, T.C. Salinity and drought—A comparison of their effects on osmotic adjustment, assimilation, transpiration and growth. Irrig. Sci. 1986, 7, 249–264. [Google Scholar] [CrossRef]
- Feddes, R.A. Agrometeorological aspects of emergence, water use, growth and dry matter yield of potatoes. Acta Hortic. 1987, 214, 45–52. [Google Scholar] [CrossRef]
- Frensch, J. Primary response of root and leaf elongation to water deficit in the atmosphere and soil solution. J. Exp. Bot. 1997, 48, 985–999. [Google Scholar] [CrossRef]
- Vos, J.; Haverkort, J. Water availability and potato crop performance. In Potato Biology and Biotechnology: Advances and Perspectives; Vreugdenhil, D., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 333–351. [Google Scholar]
- Hassanpanah, D. Evaluation of potato advanced cultivars against water deficit stress under in vitro and in vivo condition. Biotechnology 2010, 9, 164–169. [Google Scholar] [CrossRef] [Green Version]
- Pino, M.T.; Avila, A.; Molina, A.; Jeknic, Z.; Chen, T.H.H. Enhanced in vitro drought tolerance of Solanum tuberosum and Solanum commersonii plants overexpressing the ScCBF1 gene. Cienc. Investig. Agraria 2013, 40, 171–184. [Google Scholar] [CrossRef] [Green Version]
- Mustata, R.A.; Molnar, I.; Denes, T.E.; Aurori, A.; Rakosy-Tican, E. In vitro screening for drought tolerance of different marker-free transgenic/ putatively transgenic lines—Prepared to be submitted for publication.
- Kiyosue, T.; Yoshiba, Y.; Yamaguchy-Shinozaki, K.; Shinozaki, K. A nuclear gene encoding mitochondrial proline dehydrogenase, an enzyme involved in proline metabolism, is upregulated by proline but downregulated by dehydration in Arabidopsis. Plant Cell 1996, 8, 1323–1335. [Google Scholar] [PubMed] [Green Version]
- Bohnert, H.J.; Jenson, R.G. Plant stress adaptations of making metabolism move. Trends Biotechnol. 1996, 14, 267–274. [Google Scholar] [CrossRef]
- Rehman, A.U.; Bashir, F.; Ayaydin, F.; Kóta, Z.; Páli, T.; Vass, I. Proline is a quencher of singlet oxygen and superoxide both in in vitro systems and isolated thylakoids. Physiol. Plant. 2021, 172, 7–18. [Google Scholar] [CrossRef]
- Rakosy-Tican, E.; Lörincz-Besenyei, E.; Molnár, I.; Thieme, R.; Hartung, F.; Sprink, T.; Antonova, O.; Famelaer, I.; Angenon, G.; Aurori, A. New phenotypes of potato co-induced by mismatch repair deficiency and somatic hybridization. Front. Plant Sci. 2019, 10, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, K.; Pauk, J.; Kondic-Spika, A.; Grausgruber, H.; Allahverdiyev, T.; Sass, L.; Vass, I. Co-occurrence of mild salinity and drought synergistically enhances biomass and grain retardation in wheat. Front. Plant Sci. 2019, 10, 501. [Google Scholar] [CrossRef]
- Feher-Juhasz, E.; Majer, P.; Sass, L.; Lantos Cs Csisar, J.; Turoczy, Z.; Mihaly, R.; Mai, A.; Horvath, V.G.; Vass, I.; Pauk, J. Phenotyping shows improved physiological traits and seed yield of transgenic wheat plants expressing the alfalfa aldose reductase under permanent drought stress. Acta Physiol. Plant 2014, 36, 663–673. [Google Scholar] [CrossRef] [Green Version]
- Chenu, K.; Chapman, S.C.; Hammer, G.; Mclean, G.; Salah, H.B.H.; Tardiue, F. Short-term response of leaf growth rate to water deficit scale up to whole-plant and crop levels: An integrated modelling approach in maize. Plant Cell Environ. 2008, 31, 378–391. [Google Scholar] [CrossRef]
- Molnár, I.; Besenyei, E.; Thieme, R.; Thieme, T.; Aurori, A.; Baricz, A.; Banciu, H.L.; Rakosy-Tican, E. Mismatch repair deficiency increases the transfer of antibiosis and antixenosis properties against Colorado potato beetle in the somatic hybrids Solanum tuberosum (+) S. chacoense. Pest Manag. Sci. 2017, 73, 1428–1437. [Google Scholar] [CrossRef]
- Bates, L.S. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Lobell, D.B.; Burke, M.B.; Tebaldi, C.; Mastrandrea, M.D.; Falcon, W.P.; Naylor, R.L. Prioritizing climate change adaptation needs for food security in 2030. Science 2008, 19, 607–610. [Google Scholar] [CrossRef]
- Fleisher, D.H.; Dathe, A.; Timlin, D.J.; Reddy, V.R. Improving potato drought simulations: Assessing water stress factors using a coupled model. Agric. For. Meteorol. 2015, 200, 144–155. [Google Scholar] [CrossRef]
- Shao, H.B.; Chu, L.Y.; Jaleel, C.A.; Zhao, C.X. Water-deficit stress-induced anatomical changes in higher plants. C. R. Biol. 2008, 331, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Matysik, J.; Alia Bhalu, B.; Mohanty, P. Molecular mechanisms of quenching of reactive oxygen species by proline under stress in plants. Curr. Sci. Assoc. 2002, 82, 525–532. [Google Scholar]
- Sharma, P.; Dubey, R.S. Modulation of nitrate reductase activity in rice seedlings under aluminium toxicity and water stress: Role of osmolytes as enzyme protectant. J. Plant Physiol. 2005, 162, 854–864. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, N.L.; Sairam, R.K.; Tyagi, A. Expression of delta1-pyrroline-5-carboxylate synthetase gene during drought in rice (Oryza sativa L.). Ind. J. Biochem. Biophys. 2005, 42, 366–370. [Google Scholar]
- Szabados, L.; Savoure, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Hare, P.; Cress, W. Metabolic implications of stress induced proline accumulation in plants. Plant Growth Regul. 1997, 21, 79–102. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef] [Green Version]
- Wegener, C.B.; Jansen, G.; Jürgens, H.U. Bioactive compounds in potatoes: Accumulation under drought stress conditions. Funct. Food Health Dis. 2015, 5, 108–116. [Google Scholar] [CrossRef]
- Thieme, R.; Rakosy-Tican, E.; Gavrilenko, T.; Antonova, O.; Schubert, J.; Nachtigall, M.; Heimbach, U.; Thieme, T. Novel somatic hybrids (Solanum tuberosum L. + Solanum tarnii) and their fertile BC1 progenies express extreme resistance to potato virus Y and late blight. Theor. Appl. Genet. 2008, 116, 691–700. [Google Scholar] [CrossRef]
- Thieme, R.; Rakosy-Tican, E.; Nachtigall, M.; Schubert, J.; Hammann, T.; Antonova, O.; Gavrilenko, T.; Heimbach, U.; Thieme, T. Characterization of the multiple resistance traits of somatic hybrids between Solanum cardiophyllum Lindl. and two commercial potato cultivars. Plant Cell Rep. 2010, 29, 1187–1201. [Google Scholar] [CrossRef]
- Rakosy-Tican, E.; Thieme, R.; Nachtigall, M.; Molnár, I.; Denes, T.-E. The recipient potato cultivar influences the genetic makeup of the somatic hybrids between five potato cultivars and one cloned accession of sexually incompatible species Solanum bulbocastanum Dun. Plant Cell Tissue Organ Cult. 2015, 122, 395–407. [Google Scholar] [CrossRef]
- Obidiegwu, J.; Bryan, G.J.; Jones, H.G.; Prashar, A. Coping with drought stress and adaptive responses in potato and perspectives for improvement. Front. Plan. Sci. 2015, 6, 542–565. [Google Scholar] [CrossRef] [Green Version]
- Wishart, J.; George, T.S.; Brown, L.K.; Ramsay, G.; Bradshaw, J.E.; White, P.J.; Gregory, J.P. Measuring variation in potato roots in both field and glasshouse: The search for useful yield predictors and a simple screen for root traits. Plant Soil 2013, 368, 231–249. [Google Scholar] [CrossRef]
- Basu, P.S.; Sharma, A.; Garg, I.D.; Sukumaran, N.P. Tuber sink modifies photosynthetic response in potato under water stress. Environ. Exp. Bot. 1999, 42, 25–39. [Google Scholar] [CrossRef]
- Arabzadeh, N. The impact of drought stress on photosynthetic quantum yield in Haloxylon aphyllum and Haloxylon persicum. Afr. J. Plant Sci. 2013, 7, 185–189. [Google Scholar] [CrossRef] [Green Version]
- Lei, Y.; Yin, C.; Li, C. Differences in some morphological, physiological and biochemical responses to drought stress in two contrasting populations of Populus przewalskii. Physiol. Plant. 2006, 127, 182–191. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence: A Signature of Photosynthesis, Advances in Photosynthesis and Respiration; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; Volume 19, pp. 321–362. [Google Scholar]
- Flagella, Z.; Campanile, R.G.; Stoppelli, M.C.; Caro, A.D.; Fonzo, N.D. Drought tolerance of photosynthetic electron transport under CO2-enriched and normal air in cereal species. Physiol. Plant. 1998, 104, 753–759. [Google Scholar] [CrossRef]
- Lu, C.; Zhang, J. Effects of water stress on photosystem II photochemistry and its thermostability in wheat plants. J. Exp. Bot. 1999, 50, 1199–1206. [Google Scholar] [CrossRef]
- Jefferies, R.A. Effect of drought on chlorophyll fluorescence in potato (Solanum tuberosum L.). Plant water status and the kinetics of chlorophyll fluorescence. Pot. Res. 1992, 35, 25–34. [Google Scholar] [CrossRef]
- Cavender-Bares, J.; Bazzaz, F.A. From leaves to ecosystems: Using chlorophyll fluorescence to assess photosynthesis and plant function in ecological studies. In Chlorophyll a Fluorescence: A Signature of Photosynthesis, Advances in Photosynthesis and Respiration; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; Volume 19, pp. 735–755. [Google Scholar]
- Zivcak, M.; Brestic, M.; Olsovka, K.; Slamka, P. Performance index as a sensitive indicator of water stress in Triticum aestivum L. Plant Soil Environ. 2008, 54, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jefferies, R.A. Drought and chlorophyll fluorescence in field-grown potato (Solanum tuberosum). Physiol. Plant. 1994, 90, 93–97. [Google Scholar] [CrossRef]
- Fracheboud, Y.; Leipner, J. The application of chlorophyll fluorescence to study light, temperature, and drought stress. In Practical Applications of Chlorophyll Fluorescence in Plant Biology; De Ell, J.R., Toivonen, P.M.A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 125–150. [Google Scholar]
- Rakosy-Tican, E. Combining different biotechnological tools for better introgression of resistance genes into crops: The case of potato. J. Biotechnol. 2012, 161, 18. [Google Scholar] [CrossRef]
- Rakosy-Tican, E.; Molnar, I.; Thieme, R.; Taoutaou, A. New data supporting combinatorial biotechnology towards potato resistant to its most voracious pest Colorado potato beetle. Curr. Opin. Biotechnol. 2013, 24, 42–43. [Google Scholar] [CrossRef]
- Rakosy-Tican, L.; Aurori, A.; Aurori, C.M.; Ispas, G.; Famelaer, I. Transformation of wild Solanum species resistant to late blight by using reporter gene GFP and MSH2 genes. Plant Breed. Seed Sci. 2004, 50, 119–128. [Google Scholar]
- Rakosy-Tican, E.; Thieme, R.; Aurori, A.; Erdelyi-Molnár, I.; Besenyei, E.; Mustata, A.; Margineanu, A.M.; Cruceriu, D. The application of combinatorial biotechnology in improving potato resistance to biotic and abiotic stress. Sudia Univ. Babeș-Bolyai Biol. 2016, 61, 79–88. [Google Scholar]
- Foyer, C.H.; Rasool, B.; Davey, J.B.; Hancock, R.D. Cross-tolerance to biotic and abiotic stresses in plants: A focus on resistance to aphid infestation. J. Exp. Bot. 2016, 67, 2025–2037. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physiological Responses | Biochemical Responses | Molecular Responses |
|---|---|---|
| Loss of turgor and osmotic regulation | Decrease in photosynthetic efficiency | Stress response gene expression (ABA) |
| Reduced leaf water potential | Accumulation of stress solutes in extracellular matrix and cytosol | Synthesis of specific protective proteins |
| Reduced CO2 concentration due to stomatal closure | Increase the antioxidative enzyme production | |
| Reduced growth |
| Parent Lines | Combination | SHs and BCs without MMR Deficiency | Combination | SHs with MMR Deficiency |
|---|---|---|---|---|
| S. tuberosum cv. Delikat (4×) | S.tub cv. Desiree + S.chc HL | De. C4 (6×) | S.tub cv. Desiree + S.chc HL DN gene (AtMSH2) | DeDN5.5 (4×) |
| S. chacoense HL (2×) | De. C7 (4–5×) | DeDN11.5 (4×) | ||
| S. chacoense 1G (2×) | S.tub cv. Delikat + S.chc 1G | 1913/6 (5×) | DeDN11.29 (4×) | |
| 1913/10 (4–5×) | S.tub cv. Delikat + S.chc HL DN gene | DkDN5.3 (6×) | ||
| 1552/1 * (5×) | DkDN5.6 (4×) | |||
| BC1: S.tub cv. Delikat + S.chc 1G × S.tub cv. Romanze | 1552/1/1 (4–5×) | DkDN5.11 * (6×) | ||
| 1552/1/2 * (4–5×) | DkDN5.17 (6×) | |||
| 1552/1/3 (4–5×) | DkDN5.25 (4–6×) | |||
| 1552/1/4 (5×) | DkDN11.24 (6×) | |||
| 1552/1/7 (4×) | DkDN11.26 (5–6×) | |||
| 1552/1/18 (4×) | DkDN11.34 (4–5×) | |||
| 1553/1/7 (3–4×) | S.tub cv. Delikat + S.chc HL AS gene | DkS10.5 (4×) | ||
| BC2: BC1 × S.tub cv. Sonate | 1552/1/7/1 (4×) | DkS10.11 (4×) | ||
| 1552/1/7/2 (4–5×) | DkS10.13 * (4×) | |||
| DkS10.20 (4–6×) | ||||
| DkS10.35 (4–5×) | ||||
| DkS10.40 * (4×) | ||||
| DkS10.43 (4×) | ||||
| DkS10.47 (4×) | ||||
| DkS10.51 (4×) | ||||
| DkS10.61 (4×)) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molnár, I.; Cozma, L.; Dénes, T.-É.; Vass, I.; Vass, I.-Z.; Rakosy-Tican, E. Drought and Saline Stress Tolerance Induced in Somatic Hybrids of Solanum chacoense and Potato Cultivars by Using Mismatch Repair Deficiency. Agriculture 2021, 11, 696. https://doi.org/10.3390/agriculture11080696
Molnár I, Cozma L, Dénes T-É, Vass I, Vass I-Z, Rakosy-Tican E. Drought and Saline Stress Tolerance Induced in Somatic Hybrids of Solanum chacoense and Potato Cultivars by Using Mismatch Repair Deficiency. Agriculture. 2021; 11(8):696. https://doi.org/10.3390/agriculture11080696
Chicago/Turabian StyleMolnár, Imola, Lavinia Cozma, Tünde-Éva Dénes, Imre Vass, István-Zoltán Vass, and Elena Rakosy-Tican. 2021. "Drought and Saline Stress Tolerance Induced in Somatic Hybrids of Solanum chacoense and Potato Cultivars by Using Mismatch Repair Deficiency" Agriculture 11, no. 8: 696. https://doi.org/10.3390/agriculture11080696
APA StyleMolnár, I., Cozma, L., Dénes, T. -É., Vass, I., Vass, I. -Z., & Rakosy-Tican, E. (2021). Drought and Saline Stress Tolerance Induced in Somatic Hybrids of Solanum chacoense and Potato Cultivars by Using Mismatch Repair Deficiency. Agriculture, 11(8), 696. https://doi.org/10.3390/agriculture11080696