3.2. Analysis of Production
The increase in salinity significantly affected the yield of ‘Tahiti’ acid lime grafted onto the rootstocks in the first year of production (
Table 4), with effects of the interaction between rootstocks and water salinity levels on the number of fruits per plant (NFPL), weight of fruits per plant (WFPL), and average fruit weight (AFW) (
p ≤ 0.05). When considering the single factors, significant effects of rootstocks were observed for AFW (
p ≤ 0.01), and salinity affected (
p ≤ 0.01) all variables analyzed.
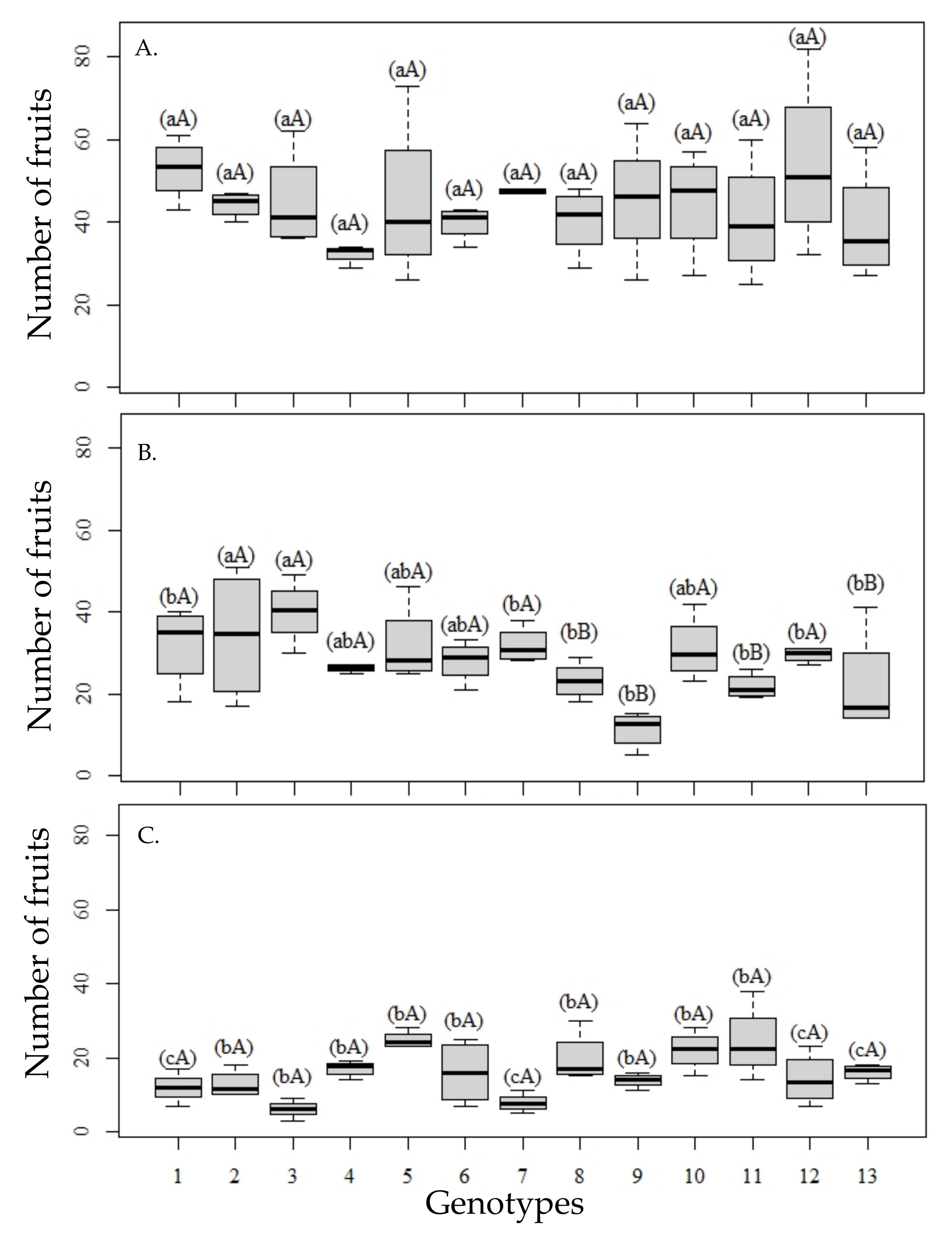
The increased water salinity reduces the number of fruits per plant, but differently among the rootstocks (
Figure 1). Under 0.14 dS m
−1 (
Figure 1A), there were no differences between genotypes, according to the means from the cluster test, although genotypes 6 and 7, corresponding to hybrids between the common ‘Sunki’ mandarin (TSKC) and
P. trifoliata Benecke (TRBK)—007 (TSKC × TRBK—007) and between the ‘Sunki of Florida’ mandarin (TSKFL) and the selection of trifoliate orange (
P. trifoliata) (TSKFL × TRBK—030), respectively, in addition to the citrandarins ‘San Diego’ (genotype 4) and ‘Indio’ (genotype 2), had lower variability in the number of fruits (
Figure 1).
Water with 2.4 dS m
−1 (
Figure 1B) caused a reduction in the number of fruits per plant, with the distinction of two groups of genotypes, with the lowest means observed in genotypes 8, 9, 11, and 13, corresponding to the hybrids TSKC × ‘Troyer’ citrange (CTTR)—012, TSKFL × CTTR—013, TSKC × (‘Rangpur’ lime (LCR) ×
P. trifoliata (TR))—040, and citrimoniandarin and TSKC × ‘Argentina’ citrange (CTARG)—019, respectively, which denotes higher sensitivity, already at this salinity level. On the other hand, citrandarins TSKC × TRBK—007, ‘San Diego’ and TSKFL × TRBK—030, and the citrimoniandarin TSKC × (LCR × TR)—059, besides being in the group of genotypes with the highest number of fruits, showed greater stability in the first year.
Irrigation with waters of 4.8 dS m
−1 (
Figure 1C) did not show distinction among genotypes according to the Scott–Knott test; however, in genotypes such as ‘Rangpur Santa Cruz’ lime, TSKFL × TRBK—030, TSKC × (LCR × TR)—059, and TSKC × CTARG—019, maximum reductions were observed in the number of fruits per plant compared to the values obtained when the plants were irrigated with waters of 0.14 dS m
−1. Furthermore, under the condition of higher water salinity, the mean values observed mostly showed less variation, which can be observed by the size of the boxplot.
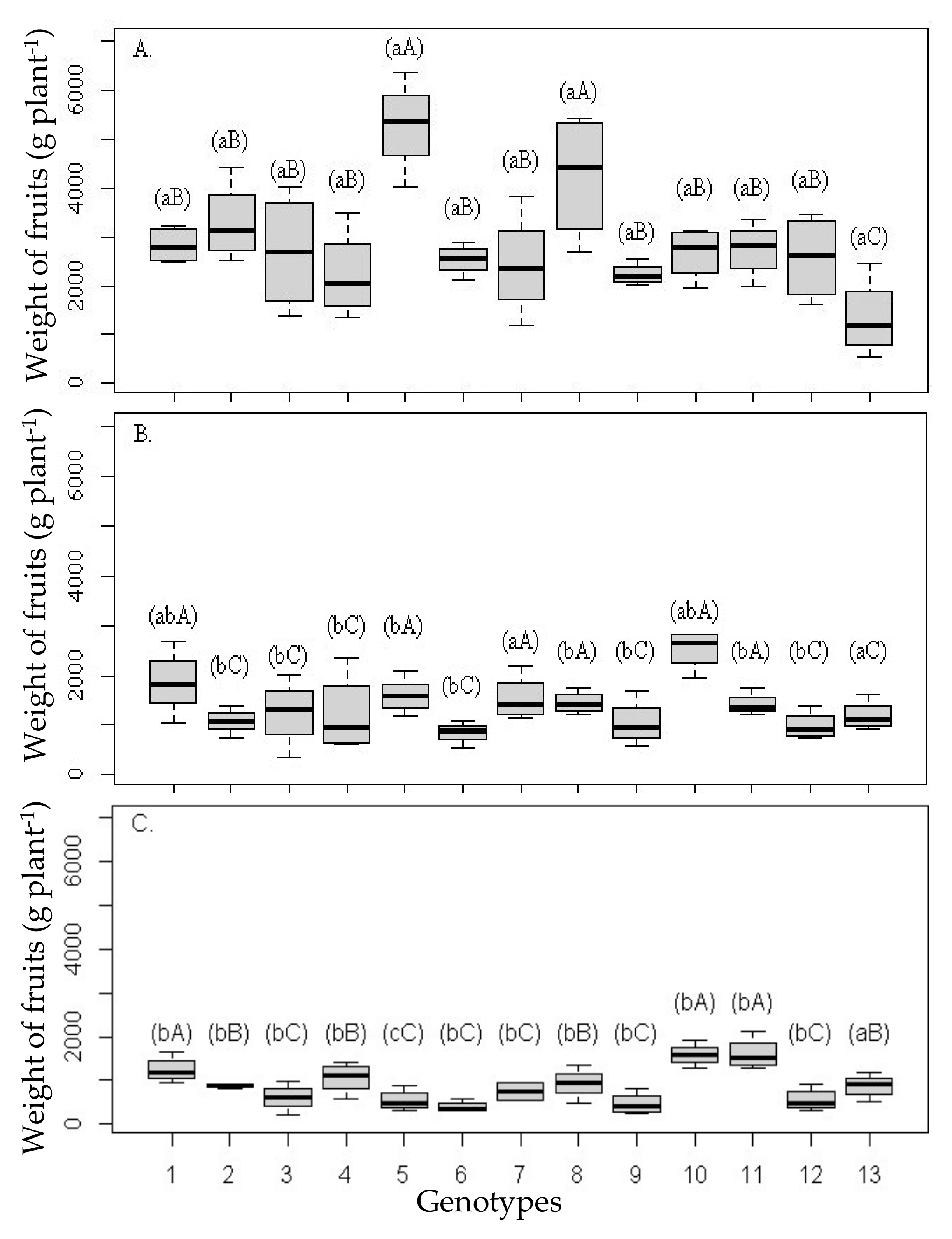
Weight of fruits per plant (
Figure 2A–C) was reduced by salinity in all scion/rootstock combinations; however, distinct groups were formed only when applying water with lower salinity, with the highest values of fruit weight in plants grafted with ‘Rangpur Santa Cruz’ lime, ‘Indio’ and ‘Riverside’ citrandarins, ‘Sunki Tropical’ mandarin, and with hybrids TSKFL × TRBK—030, HTR—069, and TSKC × (LCR × TR)—059, according to the Scott–Knott test (
p ≤ 0.05).
When analyzing the effect of salinity on the mean fruit production per rootstock plant, it is possible to verify greater relative reductions in plants grafted with ‘Santa Cruz Rangpur’ lime, ‘Indio’ citrandarin, and the hybrid TSKFL × TRBK—030, with a reduction in fruit weight greater than 60% when the plants were irrigated with water of 4.8 dS m
−1 (
Figure 2C) compared to the results obtained in these combinations irrigated with water of 0.14 dS m
−1 (
Figure 2A).
The ‘Sunki Tropical’ mandarin, HTR—069 citrangor, and TSKC × (LCR × TR)—059 citrimoniandarin, on the other hand, gave the ‘Tahiti’ acid lime greater stability in variable weight of fruits (
Figure 2), even with the increase in the salinity level in the first year of cultivation, i.e., there was a smaller reduction in the mean production of fruits with the increase in water salinity.
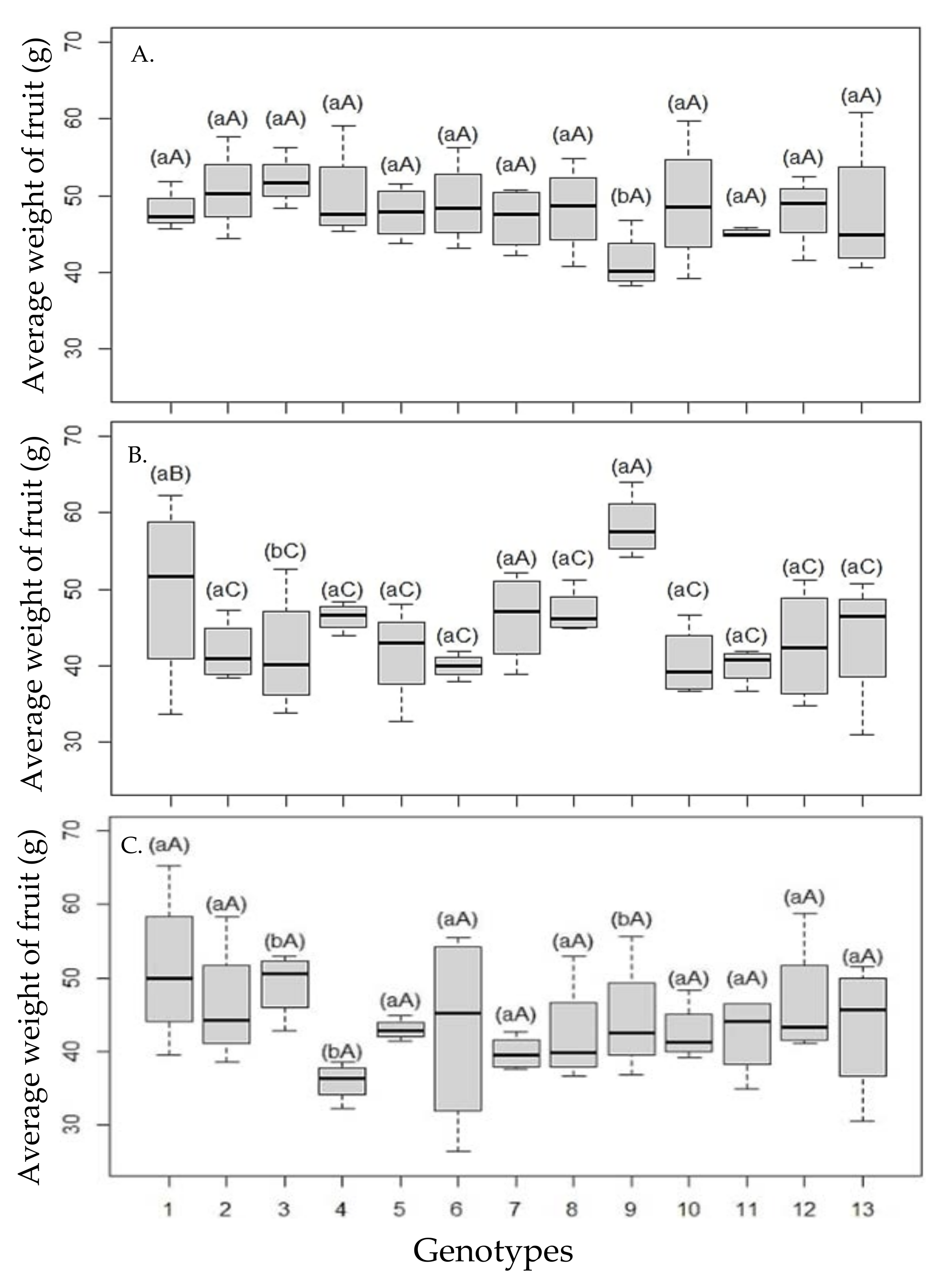
The salinity caused a loss of production in the number and weight of fruits per plant, but in general, the plants tried to maintain the mean weight of fruit in the first year of cultivation (
Figure 3A–C), even with the increase in water salinity, with a significant reduction in the mean weight of ‘Tahiti’ fruit when grafted on ‘Riverside’ and ‘San Diego’ citrandarins.
As for the distinction between genotypes used as rootstocks in salinity levels, differentiation is highlighted when irrigated with water of 2.4 dS m−1, with the formation of three groups of genotypes, highlighting the ‘Santa Cruz Rangpur’ lime and TSKFL × CTTR—013 as materials that conferred the highest average weight of fruit to the plants, thus maintaining the quality.
When analyzing the production of the second year (
Table 5), there were effects of the interaction between rootstocks and water salinity levels on the number of fruits per plant (NFPL), fruit production (WFPL), and average fruit weight (AFW) (
p ≤ 0.05). When considering the factors independently, significant effects were not observed only for the genotypes used as rootstocks in the average fruit weight (AFW) (
p ≤ 0.01), whereas salinity (
p ≤ 0.01) caused effects on all production variables studied.
As occurred in the first year of production, the number of fruits per plant was reduced by the increase in ECw (
Figure 4), and the means grouping test showed a higher number of fruits when the rootstocks were ‘Sunki Tropical’ mandarin and the hybrid TSKC × CTTR—012, under lower salinity (0.14 dS m
−1) (
Figure 4A). It can also be verified that there was no significant difference between the other rootstocks studied, and the hybrid TSKC × CTARG led to a lower average number of fruits per plant, while ‘Rangpur Santa Cruz’ lime and the citrandarin TSKC × TRBK—007 were related to the lower variability in the production of the second year.
Application of waters with 2.4 dS m
−1 (
Figure 4B) caused, in addition to the overall reduction in the number of fruits per plant, the formation of three groups of genotypes, with the highest means observed when the rootstock was the citrangor HTR—069. The TSKC × TRBK—007 and TSKFL × TRBK—030 citrandarins, TSKC × CTTR—012 citrangedarin, and TSKC × (LCR × TR)—040 citrimoniandarin led to lower variability in production in the second year of cultivation.
Water with salinity of 4.8 dS m
−1 caused reductions in the number of fruits in all genotypes (
Figure 4C). However, as in the first year of production, lower reductions were observed when the rootstocks were HTR—069 and TSKC × (LCR × TR)—040, which indicates that these rootstocks are better indicated for ‘Tahiti’ acid lime under salinity.
The weight of fruits per plant (
Figure 5) was also reduced by the increase in water salinity in all scion/rootstock combinations, and it was possible to identify three distinct groups of combinations at the three salinity levels.
Regarding irrigation with water of 0.14 dS m
−1 (
Figure 5A), the highest means of the weight of fruits per plant were observed for the rootstocks ‘Sunki Tropical’ mandarin and TSKC × CTTR—012 citrangedarin.
When the water of 2.4 dS m
−1 was used in irrigation (
Figure 5B), higher means of weight of fruits per plant were observed for ‘Santa Cruz Rangpur’ lime, ‘Sunki Tropical’ mandarin, TSKFL × TRBK—030, TSKC × CTTR—012, HTR—069, and TSKC × (LCR × TR)—040.
Conversely, when applying water of 4.8 dS m
−1 (
Figure 5C), the rootstocks ‘Rangpur Santa Cruz’ lime, HTR—069, and TSKC × (LCR × TR)—040 remained in the group of higher means according to the Scott–Knott test (
p ≤ 0.05), especially the trifoliate hybrid HTR—069, which showed lower variability, proving to have a good level of salinity tolerance.
The mean weight of fruits in the second year of production was significantly reduced by the salinity of the water in some genotypes, different from what occurred in the first year (
Figure 6A–C), with the greatest reductions in the weight of the fruits being verified. However, the ‘Tahiti’ acid lime grafted on ‘San Diego’ and TSKFL × TRBK—030 citrandarins were also highlighted, which were sensitive to salinity in the first year, too, in addition to plants grafted on ‘Sunki Tropical’ mandarin, TSKC × CTTR—012, and TSKC × (LCR × TR)—059, with an estimated mean reduction of 50 g fruit
−1 when irrigated with water of 0.14 dS m
−1 (
Figure 6A) and a mean fruit mass between 35 and 40 g when they were irrigated with water of 4.8 dS m
−1 (
Figure 6C).
The matrices of the analytical performance of the production data with the soil analyses (
Figure 7A,B) show the correlations of the first and second years of cultivation, where it is possible to observe negative numbers (without highlighting) related to the negative correlation and positive numbers (highlighted in bold) related to the positive correlation. It was also observed that the higher value means the prediction of the correlation between the variables; for example, the increase in ECse had a positive and predictive correlation with Ca
2+, Mg
2+, and Na
+ contents in both years, while on the other hand, ECse had a negative and predictive correlation with production variables. However, highlighting the behavior of K
+ in the soil during the first and the second years, correlations were predictive with ECse, but positive in the first year and negative in the second year.
In the first year, a positive correlation was observed between K+ (0.75) and ECse. However, the opposite was observed in the second year, with a negative correlation (−0.39). In the two years of cultivation, the interaction between water salinity and the soil variables pH, ECse, Ca2+, Mg 2+, and SARse was positive and predictive, with a very strong correlation, close to 1, as water with higher electrical conductivity was employed in irrigation.
3.3. Chlorophyll a Fluorescence Analysis
The interaction between rootstocks and water salinity levels did not affect chlorophyll
a fluorescence analysis after adaptation to the dark, and no significant differences were observed among rootstocks or between salinity levels in initial fluorescence (F
0), maximum fluorescence (Fm), variable fluorescence (Fv), and quantum efficiency of photosystem II (Fv/Fm), evaluated at 270 days after the beginning of stress (
Table 6).
The fluorescence variables in the second year of cultivation, initial fluorescence (F
0), maximum fluorescence (Fm), and variable fluorescence (Fv), showed significant effects caused by the genotype x salinity interaction (
p ≤ 0.05) (
Table 7), unlike the first year of cultivation.
The effect of salinity on initial fluorescence (F
0), maximum fluorescence (Fm), and variable fluorescence (Fv) varied among genotypes. When initial fluorescence was analyzed at the conductivity of 2.4 dS m
−1 (
Figure 8B), two groups of genotypes (scion/rootstock combinations) were formed, with the highest means observed in those in which the rootstocks were ‘San Diego’ citrandarin, ‘Sunki Tropical’ mandarin, TSKC × TRBK—030 citrandarin, and TSKFL × CTTR—013 citrangedarin.
It was also possible to notice a distinction between salinity levels in plants grafted onto TSKFL × CTTR—013 citrangedarin and TSKC × (LCR × TR)—040 and TSKC × (LCR × TR)—059 citrimoniandarins. The highest values were observed when applying water of 2.4 dS m
−1 in the combinations of the hybrids TSKFL × CTTR—013 and TSKC × (LCR × TR)—059, and when applying water of 0.14 dS m
−1 in plants which had the hybrid TSKC × (LCR × TR)—040 as a rootstock (
Figure 8A).
The maximum fluorescence of plants (Fm) (
Figure 9) was significantly reduced only in ‘Tahiti’ plants grafted onto TSKC × (LCR × TR)—040 citrimoniandarin and TSKC × CTARG—019 citrangedarin, especially when they were irrigated with waters of 4.8 dS m
−1, highlighting this variable as an indicator of ionic stress.
Variable fluorescence (Fv) was different among the genotypes only at the lowest level of water salinity, highlighting two groups of genotypes (
Figure 10). However, the increase in water salinity reduced (
p ≤ 0.05) the variable fluorescence in ‘Tahiti’ plants grafted onto the hybrids TSKC × (LCR× TR)—040 and TSKC × CTARG—019, which could be related to the reduction in the maximum fluorescence values observed in these genotypes and to the increase in the minimum fluorescence values recorded in TSKC × (LCR × TR)—040 citrimoniandarin.
3.4. Gas Exchange
There was a significant effect (
p ≤ 0.05) of the interaction between rootstocks and water salinity levels (
Table 8) on the CO
2 assimilation rate (
A), stomatal conductance (
gs), transpiration (
E), and intrinsic carboxylation efficiency (
CEi) (
Table 8). Similarly, there was also a significant effect (
p ≤ 0.01) of the salinity factor on the internal CO
2 concentration (
Ci), with no significant effect of any factor on the intrinsic water use efficiency (
WUEi).
For the genotype factor, significant effects were found only in the variables A, E, gs, and CEi (p ≤ 0.01), highlighting that water salinity affects citrus rootstocks differently in most of the gas exchange variables studied.
The CO
2 assimilation rate (
A) varied among the 13 scion/rootstock combinations according to the water salinity level (
Figure 11), leading to the formation of 3 groups of genotypes when the plants were irrigated with waters of 0.14 dS m
−1 (
Figure 11A) and 2 groups of genotypes when the plants were irrigated with waters of 2.40 dS m
−1 (
Figure 11B) and 4.80 dS m
−1 (
Figure 11C) according to the Scott–Knott test (
p ≤ 0.05). However, it was found that salinity only reduced the net photosynthesis of ‘Tahiti’ plants grafted onto ‘Santa Cruz Rangpur’ lime, ‘Riverside’ citrandarin, TSKC × CTTR—012 and TSKC × CTARG—019 citrangedarins, and TSKC × (LCR × TR)—040 citrimoniandarin.
When irrigation was performed with waters of 4.8 dS m−1, it was found that ‘Indio’ citrandarin, ‘Sunki Tropical’ mandarin, and the hybrids TSKC × TRBK—007, TSKFL × TRBK—030, HTR—069, and TSKC × (LCR × TR)—059 led to a higher value of assimilation rate in the scion variety, being grouped among the genotypes with the highest means.
On the other hand, the largest reductions in assimilation rate (A) were noted for Tahiti grafted on ‘Rangpur Santa Cruz’ lime, ‘Riverside’ citrandarin, TSKC × CTTR—012 citrangedarin, TSKC × (CSF × TR)—040 citrimoniandarin, and TSKC × CTARG—019 citrangedarin.
Figure 12 contains the boxplots related to the means of transpiration (
E) of the 13 scion/rootstock combinations under irrigation with waters of 0.14 dS m
−1 (
Figure 12A), 2.40 dS m
−1 (
Figure 12B), and 4.80 dS m
−1 (
Figure 12C), and it is possible to verify the rootstocks ‘Santa Cruz Rangpur’ lime and the ‘Riverside’ citrandarin between the genotypes with the highest mean values at all salinity levels. However, the increase in salinity, in general, caused a reduction in plant transpiration, but this reduction was more significant when water with an ECw of 4.8 dS m
−1 was applied to the hybrids TSKC × CTTR—012, TSKC × (LCR × TR)—040, and TSKC × CTARG—019, and the rootstock ‘Santa Cruz Rangpur’ lime.
Stomatal conductance (
gs) (
Figure 13) was reduced by salinity differently among the rootstocks, and higher values of
gs mean that the stomata of the plant are more open, which allows the influx of CO
2 and, consequently, substrate for photosynthesis. In this context, although ‘Santa Cruz Rangpur’ lime and ‘Riverside’ citrandarin were in the group of the highest means when irrigated with waters of 0.14 dS m
−1 (
Figure 13A), when irrigated with water of 4.8 dS m
−1 (
Figure 13C), they showed reductions of the order of 54.5% and 30.8% in
gs values, respectively.
The evaluation of gas exchange (
Table 9) in the second year of cultivation revealed a significant effect of the interaction between rootstocks and salinity levels only for stomatal conductance (
gs) (
p ≤ 0.01). For the genotype factor, there was no significant effect in any of the gas exchange variables. Salinity as an isolated factor affected the CO
2 assimilation rate (
A) and transpiration (
E), indicating, besides the ionic effect already observed in fluorescence reactions, osmotic effects on the plants, a situation that confirms the stress condition.
The CO
2 assimilation rates (
A) were not statistically different among the 13 scion/rootstock combinations at any of the salinity levels of irrigation water (
Figure 14A–C), i.e., no groups of genotypes with higher means were formed. However, the effect of salinity was different among the genotypes, with a reduction in net photosynthesis in ‘Tahiti’ plants grafted onto the hybrid TSKC × CTARG—019, indicative of its sensitivity since decreases were observed in gas exchange and fluorescence variables.
There was no difference (
p > 0.05) between rootstocks in terms of transpiration rate (
E) and CO
2 assimilation rate of ‘Tahiti’ acid lime (
Figure 15A–C). Even with the application of saline water with electrical conductivity levels of 2.4 and 4.8 dS m
−1, the recorded values of
E were between 1.0 and 3.0 mmol H
2O m
−2 s
−1, except for the values observed in ‘Indio’ citrandarin, which showed greater variation under an ECw of 4.8 dS m
−1. The reduction in transpiration values should be highlighted, especially in plants grafted onto TSKC × CTARG—019 citrangedarin.
Stomatal conductance (
gs) did not vary between the rootstocks at each water salinity level applied (
Figure 16), but it was observed that the effect of salinity was different among genotypes, especially because there was a significant reduction in the values of stomatal conductance in the hybrid TSKC × CTARG—019 under an ECw of 4.8 dS m
−1.


 ,
, 



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}