
Selection, Formulation, and Field Evaluation of Bacillus amyloliquefaciens PMB01 for Its Application to Manage Tomato Bacterial Wilt Disease




Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Media
2.2. Isolation of Rhizobacteria and In-Vitro Antagonistic Assays
2.3. Identification and Characterization of Antagonistic Rhizobacterium
2.3.1. Molecular Identification
2.3.2. BIOLOG GEN III System
2.3.3. Assays for Sensitivity to Stress, Fungicides, Copper Agents, and Antibiotics
2.4. Preparation of Fermentation Broth for Inhibitory Assays
2.5. Greenhouse Trials for Managing Tomato Bacterial Wilt
2.6. Field Trials
2.7. Statistical Analysis
3. Results
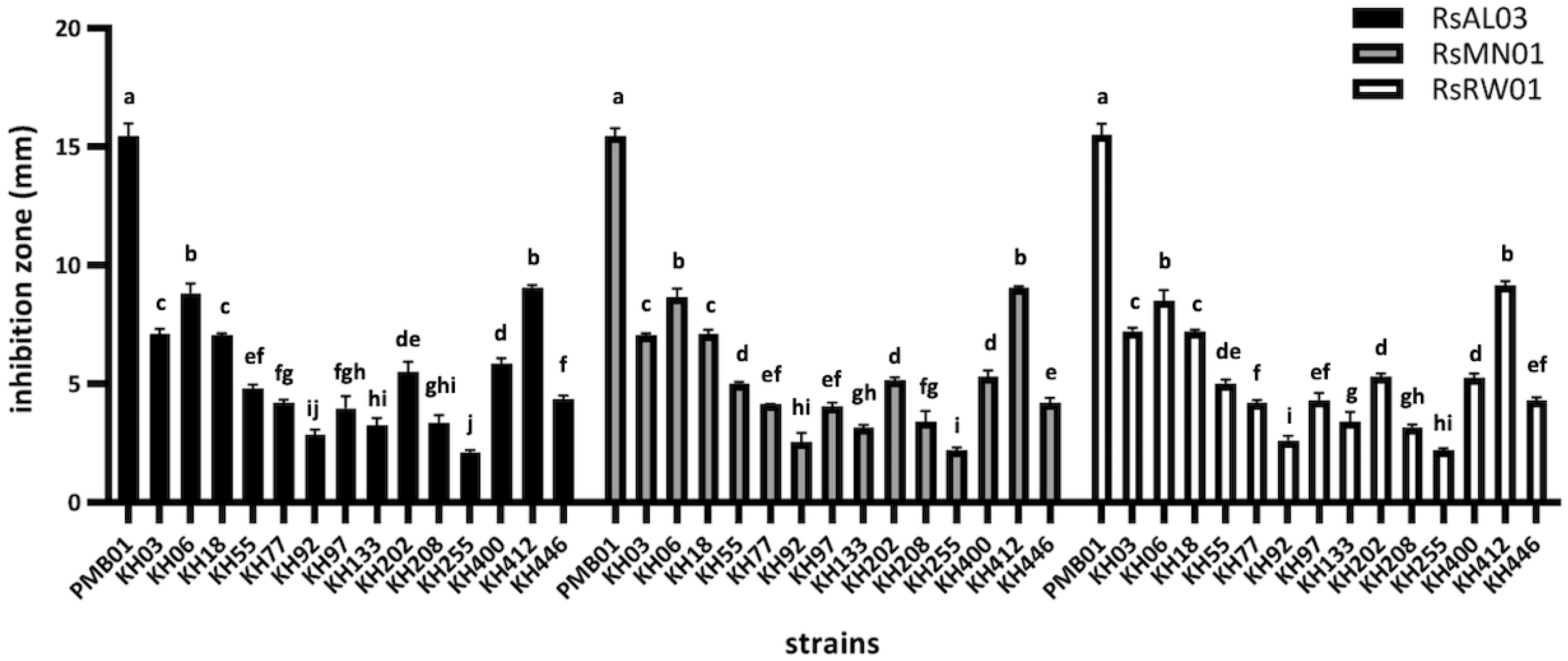
3.1. Isolation of Rhizobacteria and In-Vitro Antagonistic Activity Assays
3.2. Identification of PMB01
3.3. PMB01 Is Highly Resistant to Stress and Fungicides
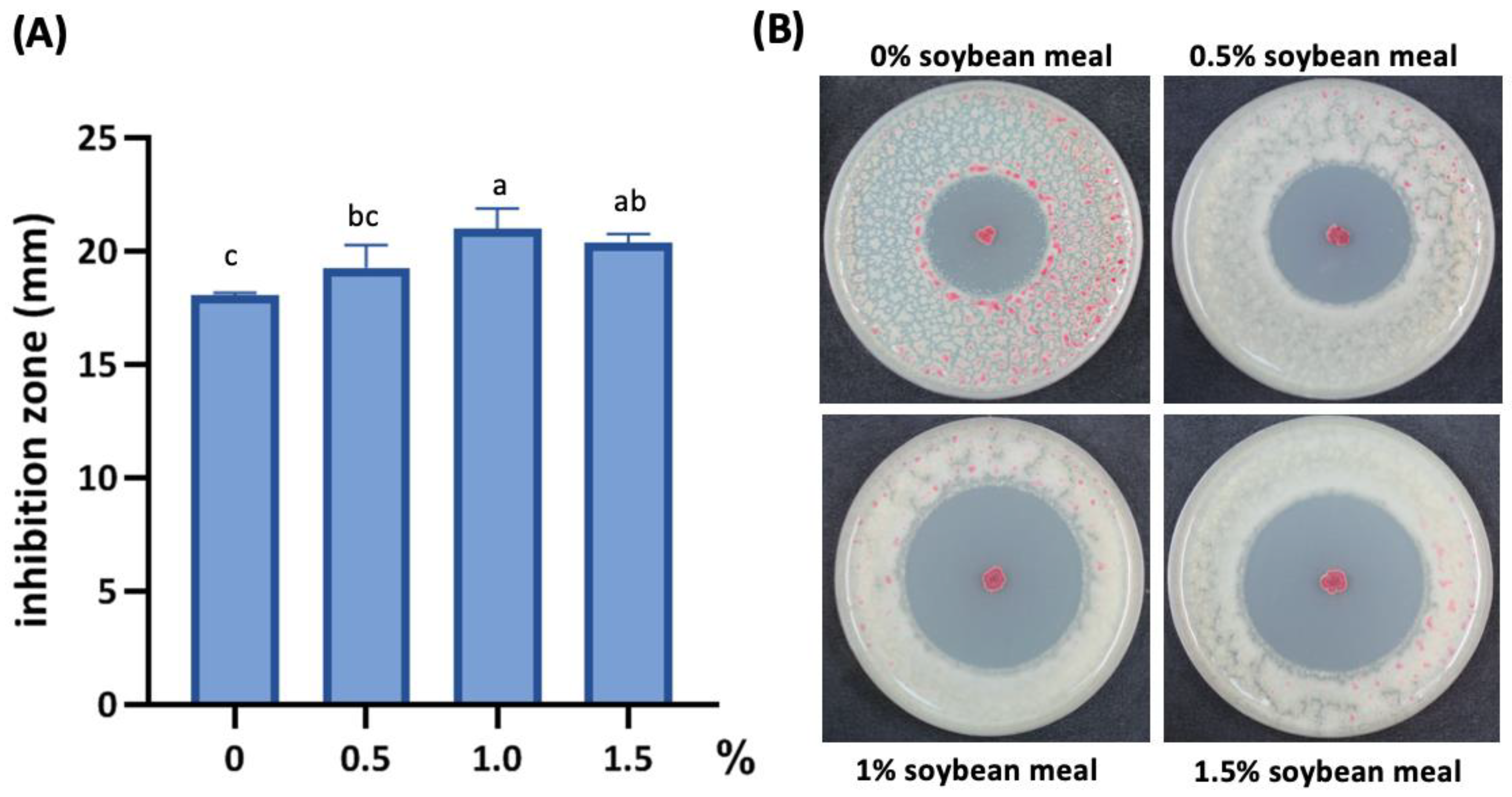
3.4. Soybean Meal Enhances the Inhibitory Activity of PMB01
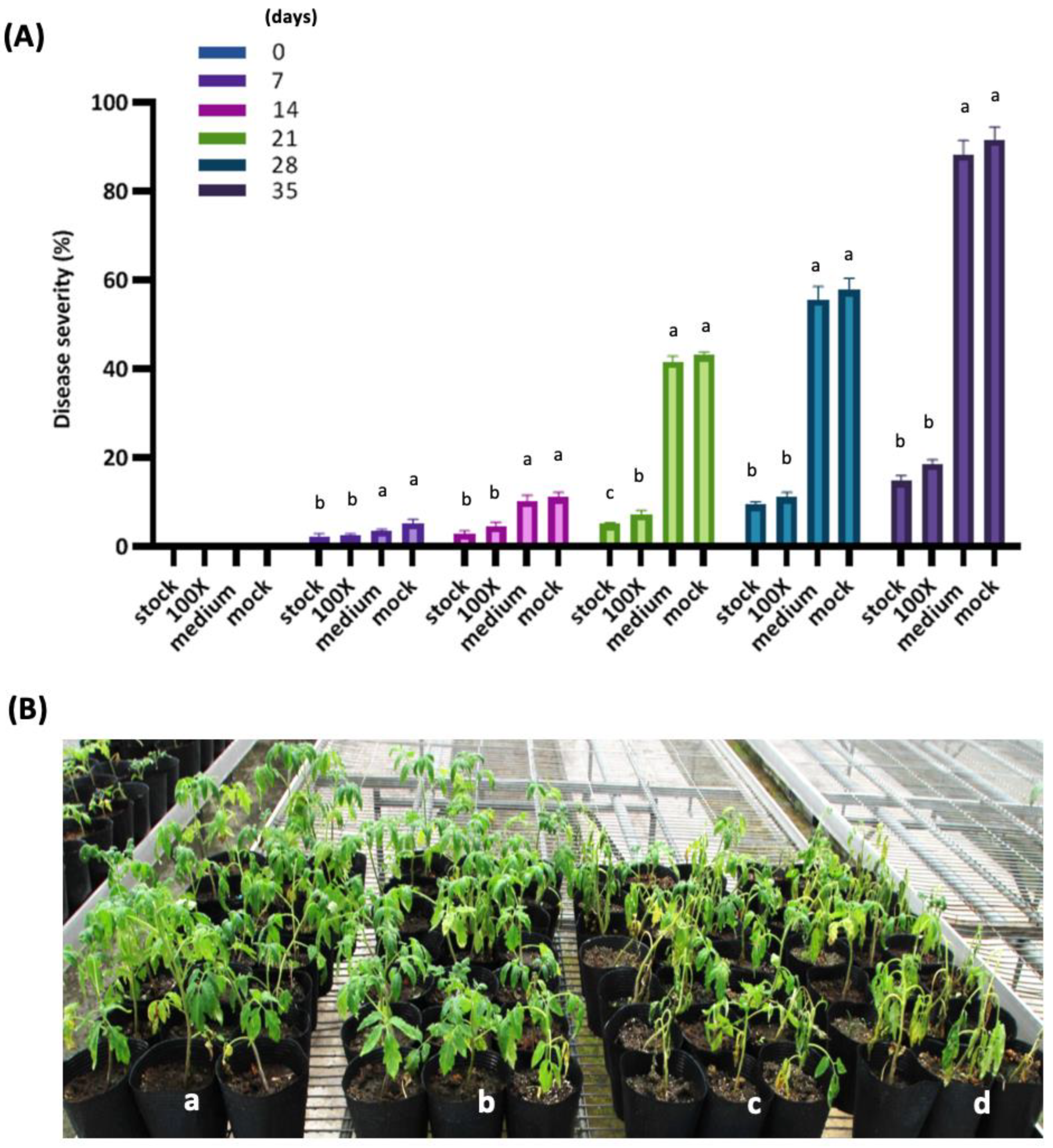
3.5. PMB01 Suppresses Bacterial Wilt Disease in Greenhouse Trials
3.6. PMB01 Formulated as a Wettable Powder Remains Active for 24 Months
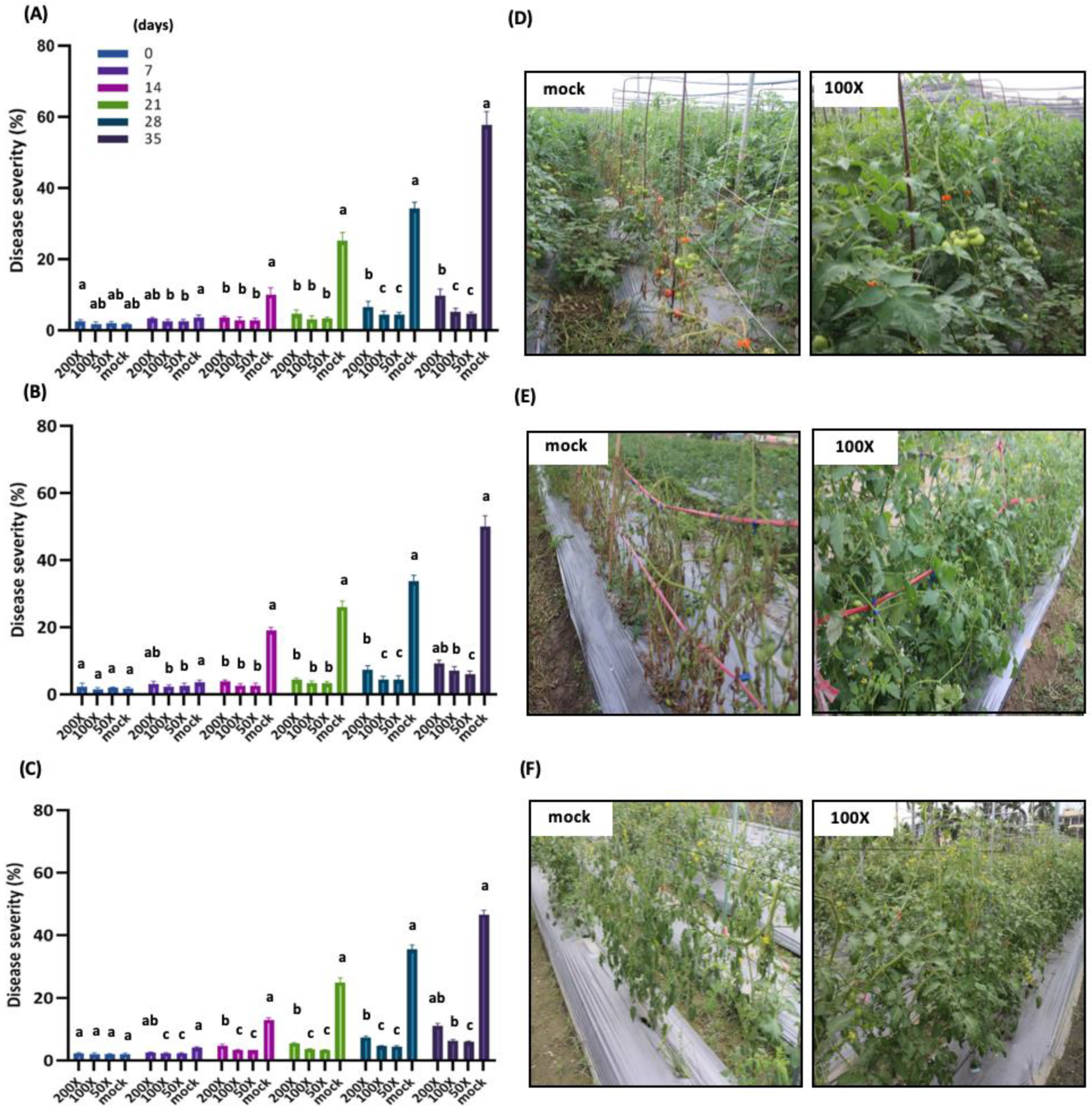
3.7. Applying PMB01 WP Suppresses Tomato Bacterial Wilt in the Field
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Pedro-Jové, R.; Puigvert, M.; Sebastià, P.; Macho, A.P.; Monteiro, J.S.; Coll, N.S.; Setúbal, J.C.; Valls, M. Dynamic expression of Ralstonia solanacearum virulence factors and metabolism-controlling genes during plant infection. BMC Genom. 2021, 22, 170. [Google Scholar] [CrossRef] [PubMed]
- Hayward, A.C. Biology and epidemiology of bacterial wilt caused by Pseudomonas solanacearum. Annu. Rev. Phytopathol. 1991, 29, 65–87. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Lin, Y.S.; Chung, W.H. Bacterial wilt of sweet potato caused by Ralstonia solanacearum in Taiwan. J. Gen. Plant Pathol. 2012, 78, 80–84. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.H.; Guo, Y.H.; Zhang, L.X.; Qi, H.Y.; Fang, Z.D. Screening for antagonism agents against cayenne pepper bacterial wilt. Chinese J. Biol. Control 2001, 17, 101–106. [Google Scholar]
- Narasimha Murthy, K.; Soumya, K.; Udayashankar, A.C.; Srinivas, C. Chapter 7—Biocontrol potential of plant growth-promoting rhizobacteria (PGPR) against Ralstonia solanacearum: Current and future prospects. In Biocontrol Agents and Secondary Metabolite; Sudisha, J., Ed.; Woodhead Publishing: Sawston, UK, 2021; pp. 153–180. [Google Scholar]
- Lin, C.H.; Chuang, M.H.; Wang, J.F. First report of bacterial wilt caused by Ralstonia solanacearum on chard in Taiwan. Plant Dis. Rep. 2015, 99, 282. [Google Scholar] [CrossRef]
- Kelman, A.; Sequeira, L. Root-to-root spread of Pseudomonas solancearum. Phytopathology 1965, 55, 304–309. [Google Scholar]
- McCarter, S.M.; Jaworski, C.A. Greenhouse studies on the spread of Pseudomonas solanacearum in tomato plants by clipping. Plant Dis. Rep. 1968, 52, 330–334. [Google Scholar]
- Hayward, A.C. Research on Bacterial Wilt: Perspective on International Linkages and Access to the Literature; American Phytopathological Society (APS) Press: St. Paul, MN, USA, 2005; pp. 1–8. [Google Scholar]
- Wang, J.F.; Lin, C.H. Integrated Management of Tomato Bacterial Wilt; AVRDC-The World Vegetable Center: Tainan, Taiwan, 2005; pp. 1–12. [Google Scholar]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef]
- Saddler, G.S. Management of Bacterial Wilt Disease; American Phytopathological Society (APS) Press: St. Paul, MN, USA, 2005; pp. 121–132. [Google Scholar]
- Verma, R.K.; Shekhawat, G.S. Effect of crop rotation and chemical soil treatment on bacterial wilt of potato. Indian Phytopathol. 1991, 44, 5–8. [Google Scholar]
- Fravel, D.R. Commercialization and implementation of biocontrol. Annu. Rev. Phytopathol. 2005, 43, 337–359. [Google Scholar] [CrossRef]
- Umesha, S.; Singh, P.K.; Singh, R.P. Chapter 6-Microbial biotechnology and sustainable agriculture. In Biotechnology for Sustainable Agriculture; Singh, R.L., Mondal, S., Eds.; Woodhead Publishing: Cambridge, UK, 2018; pp. 185–205. [Google Scholar]
- Sellitto, V.M.; Zara, S.; Fracchetti, F.; Capozzi, V.; Nardi, T. Microbial biocontrol as an alternative to synthetic fungicides: Boundaries between pre- and postharvest applications on vegetables and fruits. Fermentation 2021, 7, 60. [Google Scholar] [CrossRef]
- Emmert, E.A.; Handelsman, J. Biocontrol of plant disease: A (gram-) positive perspective. FEMS Microbiol. Lett. 1999, 171, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Moura, A.B.; da Romeiro, R.S. Use of actinomycetes preselected for the control of Ralstonia solanacearum as tomato plant growth promoters. Rev. Ceres 2000, 47, 613–626. [Google Scholar]
- Fira, D.; Dimkic, I.; Beric, T.; Lozo, J.; Stankovic, S. Biological control of plant pathogens by Bacillus species. J. Biotechnol. 2018, 285, 44–55. [Google Scholar] [CrossRef]
- Elazouni, I.; Abdel-Aziz, S.; Rabea, A. Microbial efficacy as biological agents for potato enrichment as well as bio-controls against wilt disease caused by Ralstonia solanacearum. World J. Microbiol. Biotechnol. 2019, 35, 30. [Google Scholar] [CrossRef] [PubMed]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Barka, E.A. Biological control of plant pathogens: A global perspective. Microorganisms 2022, 10, 596. [Google Scholar] [CrossRef]
- Agri, U.; Chaudhary, P.; Sharma, A. In vitro compatibility evaluation of agriusable nanochitosan on beneficial plant growth promoting rhizobacteria and maize plant. Natl. Acad. Sci. Lett. 2021, 44, 555–559. [Google Scholar] [CrossRef]
- Chaudhary, P.; Khati, P.; Gangola, S.; Kumar, A.; Kumar, R.; Sharma, A. Impact of nanochitosan and Bacillus spp. on health, productivity and defence response in Zea mays under field condition. 3 Biotech 2021, 11, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, P.; Singh, S.; Chaudhary, A.; Sharma, A.; Kumar, G. Overview of biofertilizers in crop production and stress management for sustainable agriculture. Front. Plant Sci. 2022, 13, 930340. [Google Scholar] [CrossRef] [PubMed]
- Baysal, O.; Lai, D.; Xu, H.H.; Siragusa, M.; Caliskan, M.; Carimi, F.; da Silva, J.A.; Tor, M. A proteomic approach provides new insights into the control of soil-borne plant pathogens by Bacillus species. PLoS ONE 2013, 8, e53182. [Google Scholar] [CrossRef] [Green Version]
- Raaijmakers, J.M.; De Bruijn, I.; Nybroe, O.; Ongena, M. Natural functions of lipopeptides from Bacillus and Pseudomonas: More than surfactants and antibiotics. FEMS Microbiol. Rev. 2010, 34, 1037–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Lilge, L.; Ersig, N.; Hubel, P.; Aschern, M.; Pillai, E.; Klausmann, P.; Pfannstiel, J.; Henkel, M.; Heravi, K.M.; Hausmann, R. Surfactin shows relatively low antimicrobial activity against Bacillus subtilis and other bacterial model organisms in the absence of synergistic metabolites. Microorganisms 2022, 10, 779. [Google Scholar] [CrossRef]
- Wu, B.; Wang, X.; Yang, L.; Yang, H.; Zeng, H.; Qiu, Y.; Wang, C.; Yu, J.; Li, J.; Xu, D.; et al. Effects of Bacillus amyloliquefaciens ZM9 on bacterial wilt and rhizosphere microbial communities of tobacco. Appl. Soil Ecol. 2016, 103, 1–12. [Google Scholar] [CrossRef]
- Perez, R.H.; Zendo, T.; Sonomoto, K. Circular and leaderless bacteriocins: Biosynthesis, mode of action, applications, and prospects. Front. Microbiol. 2018, 9, 2085. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Arora, N.K. Bacterial formulations and delivery systems against pests in sustainable Agro-Food production. Ref. Module Food Sci. 2016, 1–11. [Google Scholar] [CrossRef]
- Shaikh, S.S.; Sayyed, R.Z. Role of plant growth-promoting rhizobacteria and their formulation in biocontrol of plant diseases. In Plant Microbes Symbiosis: Applied Facets; Arora, N.K., Ed.; Springer: New Delhi, India, 2015; pp. 337–351. [Google Scholar]
- Yuliar, Y.; Nion, Y.A.; Toyota, K. Recent trends in control methods for bacterial wilt diseases caused by Ralstonia solanacearum. Environ. Microbiol. 2015, 30, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Edwards, U.; Rogall, T.; Blocker, H.; Emde, M.; Bottger, E.C. Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucleic Acids Res. 1989, 17, 7843–7853. [Google Scholar] [CrossRef] [Green Version]
- De Clerck, E.; Vanhoutte, T.; Hebb, T.; Geerinck, J.; Devos, J.; De Vos, P. Isolation, characterization, and identification of bacterial contaminants in semifinal gelatin extracts. Appl. Environ. Microbiol. 2004, 70, 3664–3672. [Google Scholar] [CrossRef] [Green Version]
- Gould, G.W.; Jones, A.; Wrighton, C. Limitations of the initiation of germination of bacterial spores as a spore control procedure. J. Appl. Bacteriol. 1968, 31, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Kelman, A. The relationship of pathogenicity in Pseudomonas solanacearum to colony appearance on a tetrazolium medium. Phytopathology 1954, 44, 693–695. [Google Scholar]
- Hyakumachi, M.; Nishimura, M.; Arakawa, T.; Asano, S.; Yoshida, S.; Tsushima, S.; Takahashi, H. Bacillus thuringiensis suppresses bacterial wilt disease caused by Ralstonia solanacearum with systemic induction of defense-related gene expression in tomato. Microbes Environ. 2013, 28, 128–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addelman, S. The generalized randomized block design. Am. Stat. 1969, 23, 35–36. [Google Scholar]
- Roberts, D.P.; Denny, T.P.; Schell, M.A. Cloning of the egl gene of Pseudomonas solanacearum and analysis of its role in phytopathogenicity. J. Bacteriol. 1988, 170, 1445–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaurilcikiene, I.; Deveikyte, I.; Petraitiene, R. Epidemic progress of Cercospora beticola Sacc. in Beta vulgaris L. under different conditions and cultivar resistance. Biologija 2006, 4, 54–59. [Google Scholar]
- Whipps, J.M. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot. 2001, 52, 487–511. [Google Scholar] [CrossRef]
- Singh, D.; Yadav, D.; Chaudhary, G.; Rana, V.; Sharma, R. Potential of Bacillus amyloliquefaciens for biocontrol of bacterial wilt of tomato incited by Ralstonia solanacearum. J. Plant Pathol. Microbiol. 2016, 7, 327. [Google Scholar] [CrossRef]
- Keswani, C.; Singh, H.B.; Hermosa, R.; Garcia-Estrada, C.; Caradus, J.; He, Y.W.; Mezaache-Aichour, S.; Glare, T.R.; Borriss, R.; Vinale, F.; et al. Antimicrobial secondary metabolites from agriculturally important fungi as next biocontrol agents. Appl. Microbiol. Biotechnol. 2019, 103, 9287–9303. [Google Scholar] [CrossRef]
- Chu, D.; Ilyas, N.; Peng, L.; Wang, X.; Wang, D.; Xu, Z.; Gao, Q.; Tan, X.; Zhang, C.; Li, Y.; et al. Genomic insights on fighting bacterial wilt by a novel Bacillus amyloliquefaciens strain Cas02. Microb. Biotechnol. 2022, 15, 1152–1167. [Google Scholar] [CrossRef]
- Abo-Elyousr, K.A.M.; Hassan, S.A. Biological control of Ralstonia solanacearum (Smith), the causal pathogen of bacterial wilt disease by using Pantoea spp. Egypt J. Biol. Pest. Control 2021, 31, 113. [Google Scholar] [CrossRef]
- Mulatu, A.; Alemu, T.; Megersa, N.; Vetukuri, R.R. Optimization of culture conditions and production of Bio-Fungicides from Trichoderma species under solid-state fermentation using mathematical modeling. Microorganisms 2021, 9, 1675. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, P.; Carvalho, F. Chapter 19-Microbial enzymes for the food industry. In Biotechnology of Microbial Enzymes; Brahmachari, G., Ed.; Academic Press: San Diego, CA, USA, 2017; pp. 513–544. [Google Scholar]
- Ao, T. Using exogenous enzymes to increase the nutritional value of soybean meal in poultry diet. In Soybean and Nutrition; InTechOpen: London, UK, 2011. [Google Scholar] [CrossRef] [Green Version]
- Daliri, E.B.M.; Lee, B.H.; Park, M.H.; Kim, J.H.; Oh, D.H. Novel angiotensin I-converting enzyme inhibitory peptides from soybean protein isolates fermented by Pediococcus pentosaceus SDL1409. LWT 2018, 93, 88–93. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, J.; Wang, L.; Ma, Z.; Zhao, G.; Ge, Z.; Zhu, H.; Qiao, J. Improving nitrogen source utilization from defatted soybean meal for nisin production by enhancing proteolytic function of Lactococcus lactis F44. Sci. Rep. 2017, 7, 6189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, A.C.; Lin, H.L.; Shiu, Y.L.; Tyan, Y.C.; Liu, C.H. Isolation and characterization of antimicrobial peptides derived from Bacillus subtilis E20-fermented soybean meal and its use for preventing Vibrio infection in shrimp aquaculture. Fish Shellfish Immunol. 2017, 67, 270–279. [Google Scholar] [CrossRef]
- Sanjukta, S.; Rai, A.K. Production of bioactive peptides during soybean fermentation and their potential health benefits. Trends Food Sci. Technol. 2016, 50, 1–10. [Google Scholar] [CrossRef]
- Bharti, V.; Ibrahim, S. Biopesticides: Production, formulation and application systems. Int. J. Curr. Microbiol. App. Sci. 2020, 9, 3931–3946. [Google Scholar] [CrossRef]
- Huang, J.; Wei, Z.; Tan, S.; Mei, X.; Yin, S.; Shen, Q.; Xu, Y. The rhizosphere soil of diseased tomato plants as a source for novel microorganisms to control bacterial wilt. Appl. Soil Ecol. 2013, 72, 79–84. [Google Scholar] [CrossRef]
- Singh, S.; Gautam, R.K.; Singh, D.R.; Sharma, T.V.R.S.; Sakthivel, K.; Roy, S.D. Genetic approaches for mitigating losses caused by bacterial wilt of tomato in tropical islands. Eur. J. Plant Pathol. 2015, 143, 205–221. [Google Scholar] [CrossRef]
- Kijima, T. Biological control of soil-borne diseases with antagonistic bacteria. Proc. Kanto-Tosan Plant Prot. Soc. 1992, 39, 1–5. [Google Scholar]
- Kohl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of action of microbial biological control agents against plant diseases: Relevance beyond efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, V.B.; Meyer, T.; Arias, A.A.; Ongena, M.; Oni, F.E.; Hofte, M. Bacillus cyclic lipopeptides iturin and fengycin control rice blast caused by Pyricularia oryzae in potting and acid sulfate soils by direct antagonism and induced systemic resistance. Microorganisms 2021, 9, 1441. [Google Scholar] [CrossRef] [PubMed]
- Ngalimat, M.S.; Yahaya, R.S.R.; Baharudin, M.M.A.; Yaminudin, S.M.; Karim, M.; Ahmad, S.A.; Sabri, S. A review on the biotechnological applications of the operational group Bacillus amyloliquefaciens. Microorganisms 2021, 9, 614. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (Hours) | Cells (CFU/mL) | Spores (CFU/mL) | Sporulation (%) |
|---|---|---|---|
| 0 | 6.28 ± 0.04 | 0 ± 0 ** | 0 |
| 24 | 7.34 ± 0.07 | 5.46 ± 0.13 ** | 74.39 |
| 48 | 8.26 ± 0.07 | 8.07 ± 0.06 * | 97.75 |
| 72 | 9.54 ± 0.02 | 9.54 ± 0.01 | 100 |
| 96 | 9.55 ± 0.03 | 9.55 ± 0.03 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chou, H.-P.; Huang, Y.-C.; Lin, Y.-H.; Deng, W.-L. Selection, Formulation, and Field Evaluation of Bacillus amyloliquefaciens PMB01 for Its Application to Manage Tomato Bacterial Wilt Disease. Agriculture 2022, 12, 1714. https://doi.org/10.3390/agriculture12101714
Chou H-P, Huang Y-C, Lin Y-H, Deng W-L. Selection, Formulation, and Field Evaluation of Bacillus amyloliquefaciens PMB01 for Its Application to Manage Tomato Bacterial Wilt Disease. Agriculture. 2022; 12(10):1714. https://doi.org/10.3390/agriculture12101714
Chicago/Turabian StyleChou, Hau-Ping, Yi-Chiao Huang, Yi-Hsien Lin, and Wen-Ling Deng. 2022. "Selection, Formulation, and Field Evaluation of Bacillus amyloliquefaciens PMB01 for Its Application to Manage Tomato Bacterial Wilt Disease" Agriculture 12, no. 10: 1714. https://doi.org/10.3390/agriculture12101714
APA StyleChou, H. -P., Huang, Y. -C., Lin, Y. -H., & Deng, W. -L. (2022). Selection, Formulation, and Field Evaluation of Bacillus amyloliquefaciens PMB01 for Its Application to Manage Tomato Bacterial Wilt Disease. Agriculture, 12(10), 1714. https://doi.org/10.3390/agriculture12101714