
DNA Barcoding of Endangered and Rarely Occurring Plants in Faifa Mountains (Jazan, Saudi Arabia)


Abstract
:1. Introduction
2. Materials and Methods
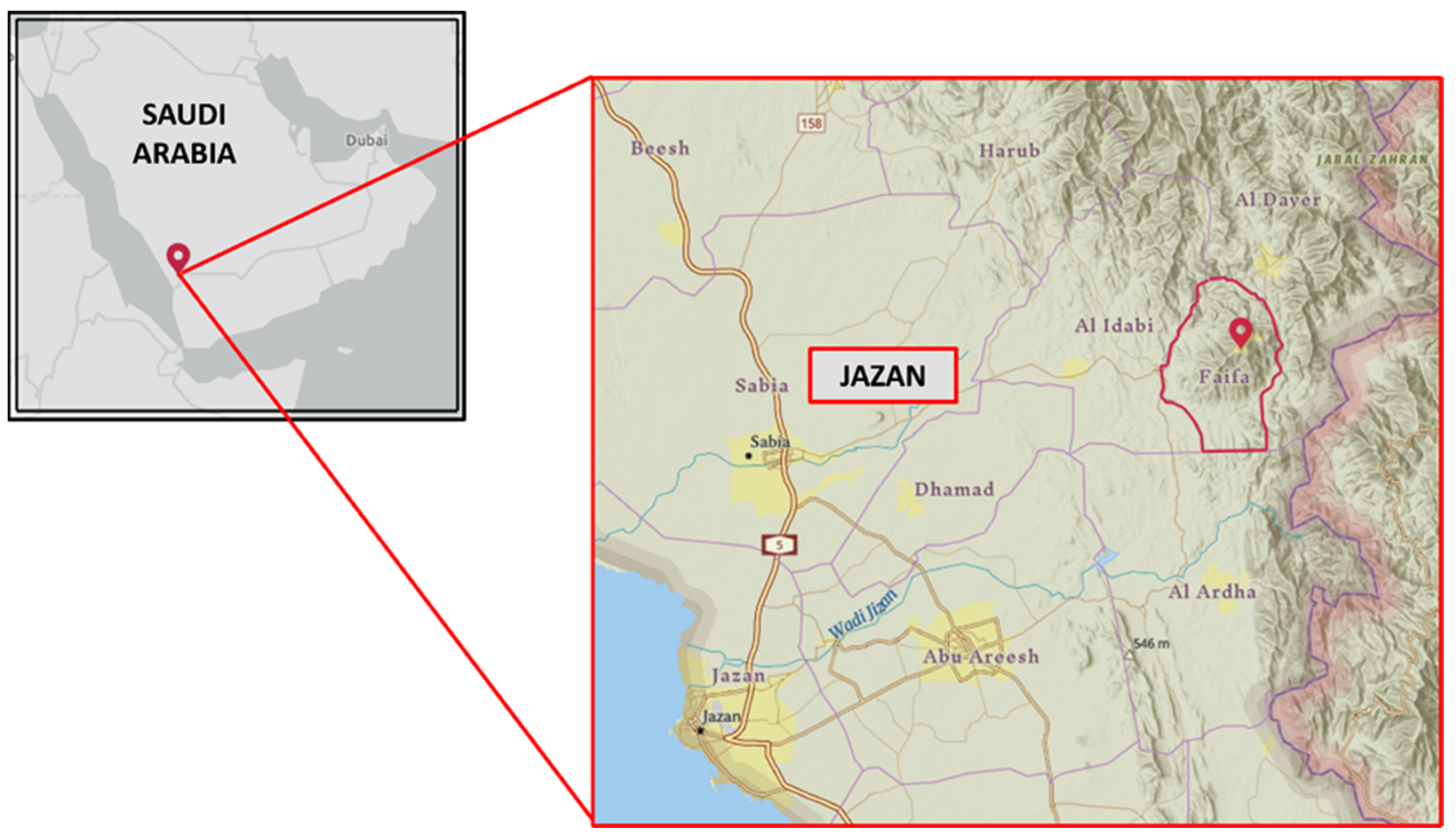
2.1. Study Area
2.2. Sample Collection
2.3. DNA Extraction, PCR Amplification, and Sequencing
2.4. Sequence Alignment and Data Analysis
3. Results
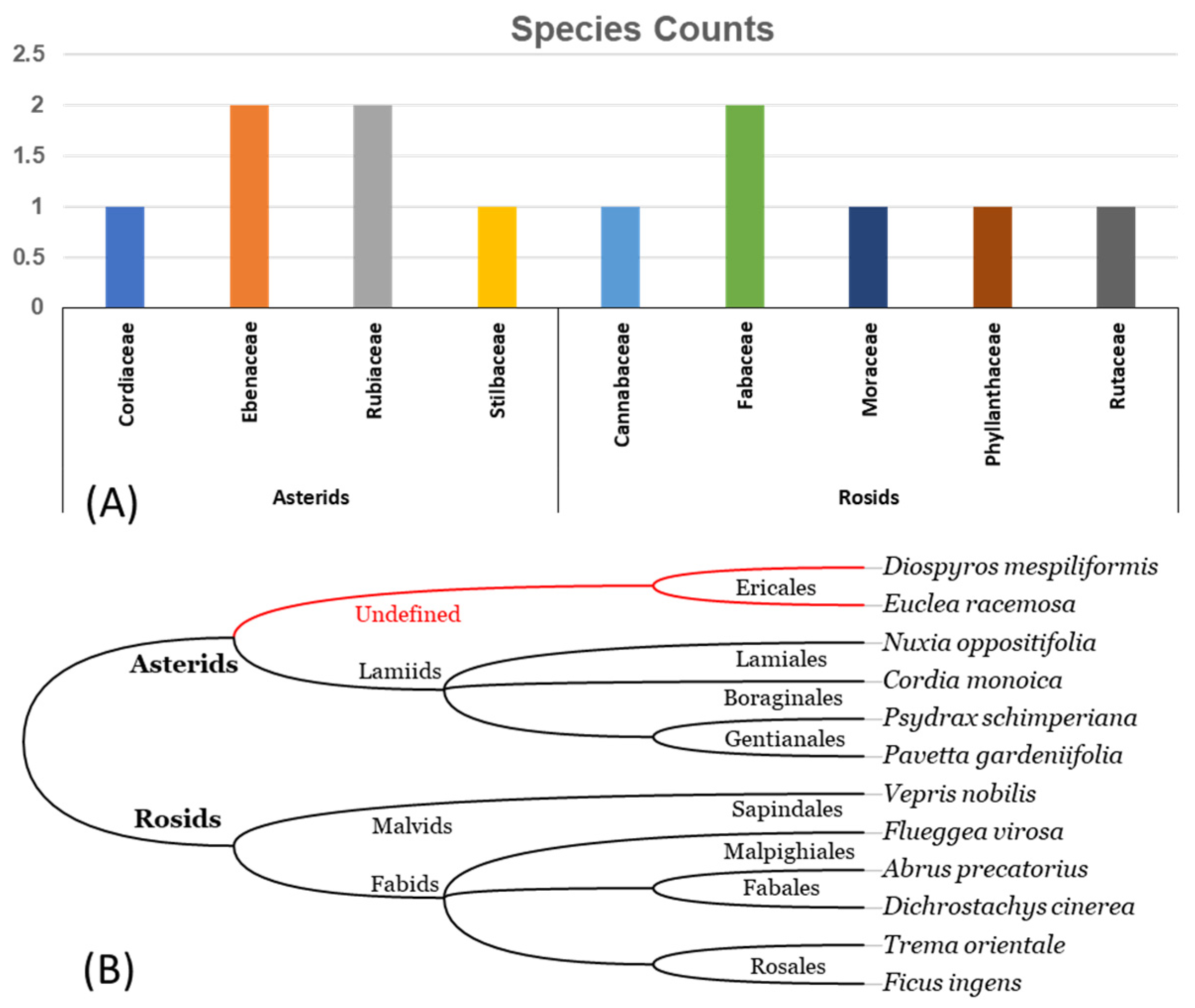
3.1. Morphological Inspection
3.2. Molecular Identification Using DNA Barcodes
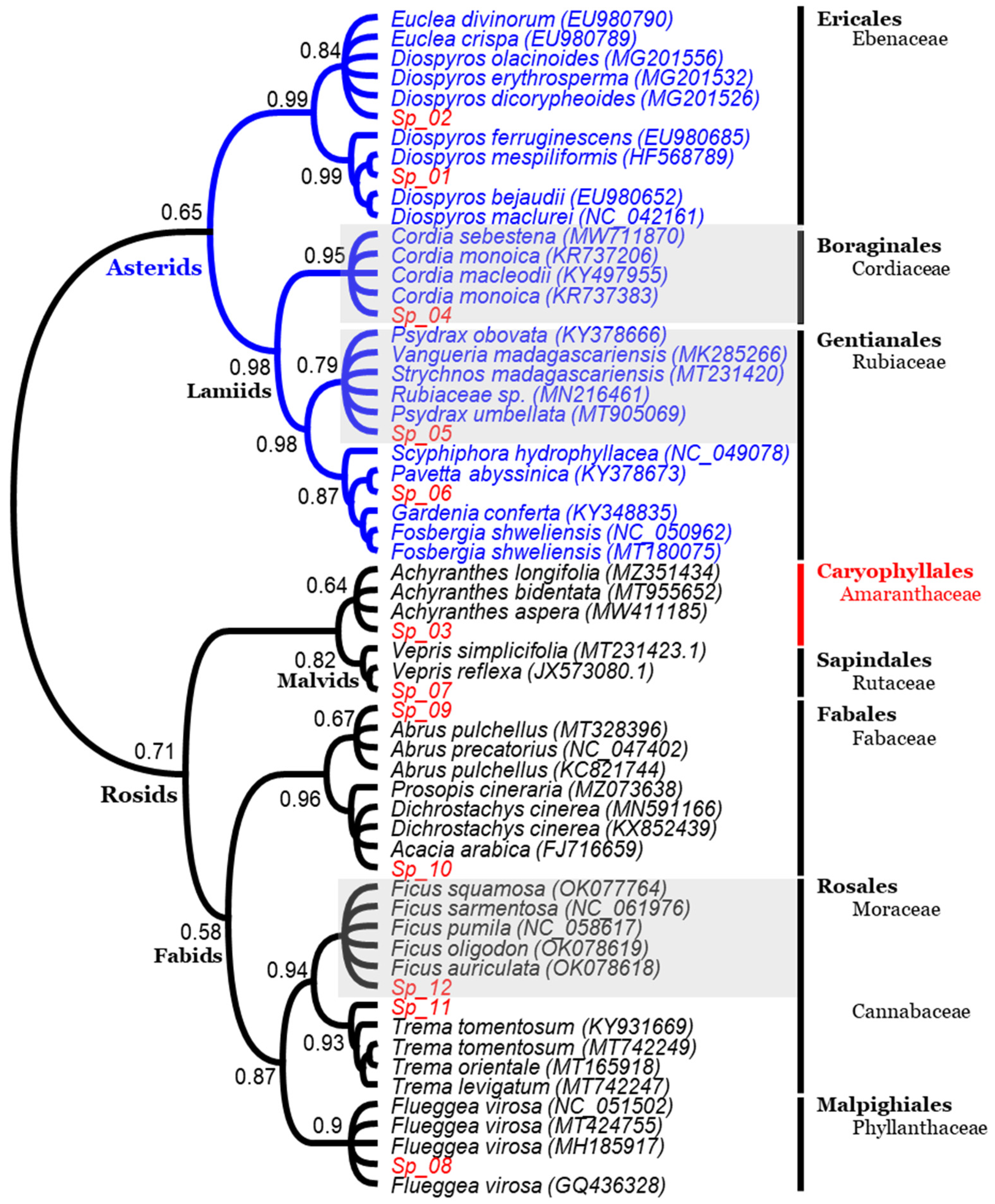
3.2.1. Chloroplast Rbcl Gene
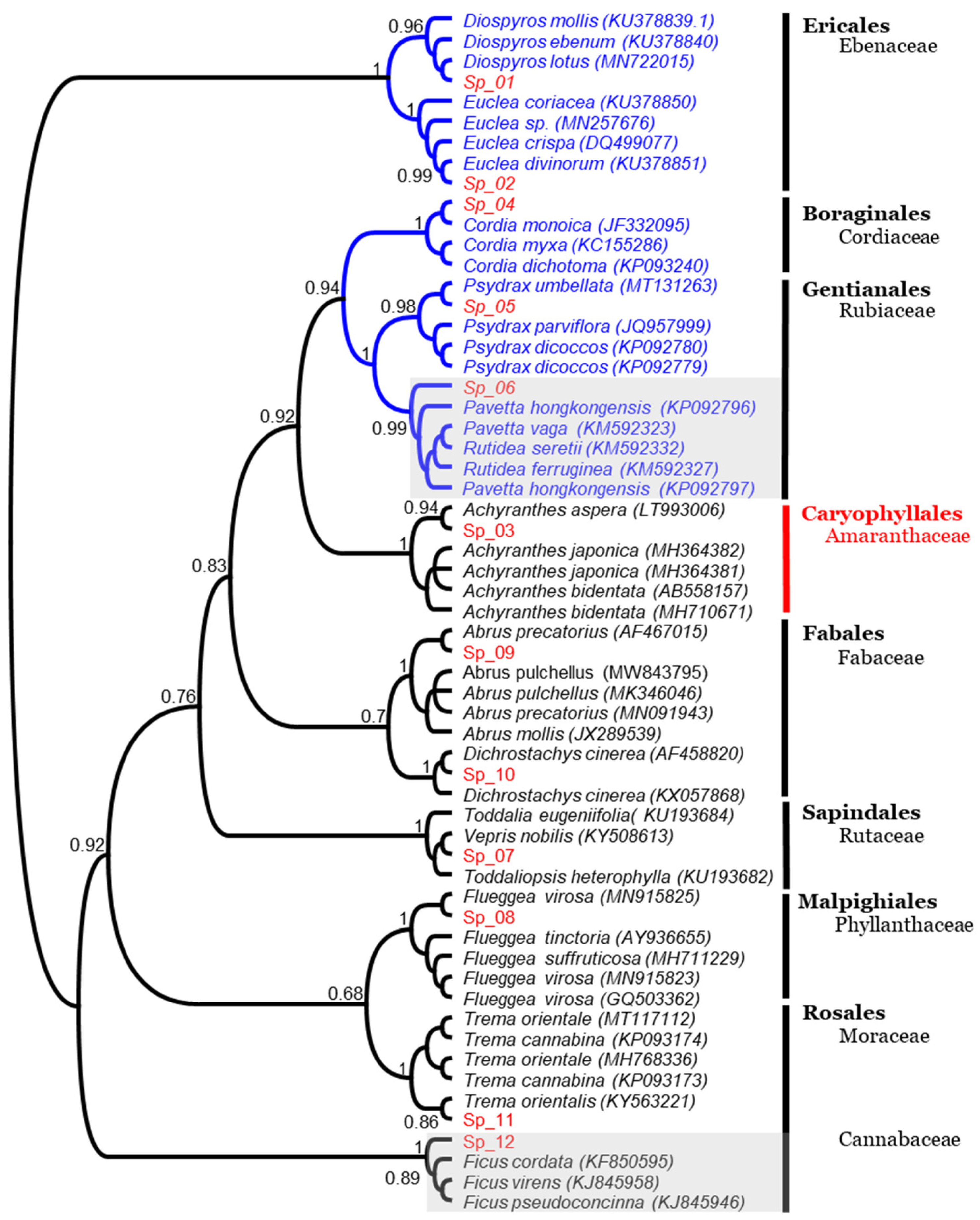
3.2.2. Nuclear ITS Region
3.3. Morpho-Molecular Comparative Analysis
3.4. Species Listing and Rarity Assessment
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CBD COP Decision. Available online: https://www.cbd.int/decision/cop/?id=12268 (accessed on 23 July 2022).
- Amaral, W.; Thomson, L.; Yanchuk, A. Conservation of Genetic Resources in Their Natural Environment. In Forest Genetic Resources Conservation and Management: Overview, Concepts and Some Systematic Approaches; International Plant Genetic Resources Institute: Rome, Italy, 2004; Volume I, Available online: https://www.bioversityinternational.org/fileadmin/_migrated/uploads/tx_news/Forest_genetic_resources_conservation_and_management__overview__concepts_and_some_systematic_approaches__Vol._1_1018.pdf (accessed on 5 June 2022).
- Phillips, O.L.; Vásquez Martínez, R.; Núñez Vargas, P.; Lorenzo Monteagudo, A.; Chuspe Zans, M.-E.; Galiano Sánchez, W.; Peña Cruz, A.; Timaná, M.; Yli-Halla, M.; Rose, S. Efficient Plot-Based Floristic Assessment of Tropical Forests. J. Trop. Ecol. 2003, 19, 629–645. [Google Scholar] [CrossRef] [Green Version]
- Webb, C.O.; Slik, J.W.F.; Triono, T. Biodiversity Inventory, and Informatics in Southeast Asia. Biodivers. Conserv. 2010, 19, 955–972. [Google Scholar] [CrossRef]
- Hartvig, I.; Czako, M.; Kjær, E.D.; Nielsen, L.R.; Theilade, I. The Use of DNA Barcoding in Identification and Conservation of Rosewood (Dalbergia spp.). PLoS ONE 2015, 10, e0138231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aubriot, X.; Lowry, P.P.; Cruaud, C.; Couloux, A.; Haevermans, T. DNA Barcoding in a Biodiversity Hot Spot: Potential Value for the Identification of Malagasy Euphorbia L. Listed in CITES A Ppendices I and II. Mol. Ecol. Resour. 2013, 13, 57–65. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; de Waard, J.R. Biological Identifications through DNA Barcodes. Proc. R. Soc. Lond. B 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- CBOL Plant Working Group; Hollingsworth, P.M.; Forrest, L.L.; Spouge, J.L.; Hajibabaei, M.; Ratnasingham, S.; van der Bank, M.; Chase, M.W.; Cowan, R.S.; Erickson, D.L.; et al. A DNA Barcode for Land Plants. Proc. Natl. Acad. Sci. USA 2009, 106, 12794–12797. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten Species in One: DNA Barcoding Reveals Cryptic Species in the Neotropical Skipper Butterfly Astraptes Fulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef] [Green Version]
- Safhi, F.A.; ALshamrani, S.M.; Jalal, A.S.; Abd El-Moneim, D.; Alyamani, A.A.; Ibrahim, A.A. Genetic Characterization of Some Saudi Arabia’s Accessions from Commiphora gileadensis Using Physio Biochemical Parameters, Molecular Markers, DNA Barcoding Analysis and Relative Gene Expression. Genes 2022, 13, 2099. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Stoeckle, M.Y.; Zemlak, T.S.; Francis, C.M. Identification of Birds through DNA Barcodes. PLoS Biol. 2004, 2, e312. [Google Scholar] [CrossRef] [Green Version]
- Sosa, V.; Mejía-Saules, T.; Cuéllar, M.A.; Vovides, A.P. DNA Barcoding in Endangered Mesoamerican Groups of Plants. Bot. Rev. 2013, 79, 469–482. [Google Scholar] [CrossRef]
- Kress, W.J.; Wurdack, K.J.; Zimmer, E.A.; Weigt, L.A.; Janzen, D.H. Use of DNA Barcodes to Identify Flowering Plants. Proc. Natl. Acad. Sci. USA 2005, 102, 8369–8374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferri, G.; Alù, M.; Corradini, B.; Beduschi, G. Forensic Botany: Species Identification of Botanical Trace Evidence Using a Multigene Barcoding Approach. Int. J. Legal Med. 2009, 123, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Mossa, J.S.; Al-Said, M.S.; Al-Yahya, M.A. Medicinal Plant Diversity in the Flora of Saudi Arabia 1: A Report on Seven Plant Families. Fitoterapia 2004, 75, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Masrahi, Y.S.; Al-Turki, T.A.; Sayed, O.H. Wolffiella Hyalina (Delile) Monod (Lemnaceae)—A New Record for the Flora of Saudi Arabia. Feddes Repert. 2010, 121, 189–193. [Google Scholar] [CrossRef]
- Levin, R.A.; Wagner, W.L.; Hoch, P.C.; Nepokroeff, M.; Pires, J.C.; Zimmer, E.A.; Sytsma, K.J. Family-level Relationships of Onagraceae Based on Chloroplast Rbc L and Ndh F Data. Am. J. Bot. 2003, 90, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Cheng, T.; Xu, C.; Lei, L.; Li, C.; Zhang, Y.; Zhou, S. Barcoding the Kingdom Plantae: New PCR Primers for ITS Regions of Plants with Improved Universality and Specificity. Mol. Ecol. Resour. 2016, 16, 138–149. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Chase, M.W.; Salamin, N.; Wilkinson, M.; Dunwell, J.M.; Kesanakurthi, R.P.; Haidar, N.; Savolainen, V. Land Plants and DNA Barcodes: Short-Term and Long-Term Goals. Philos. Trans. R. Soc. B 2005, 360, 1889–1895. [Google Scholar] [CrossRef]
- El-Atroush, H.; Magdy, M.; Werner, O. DNA Barcoding of Two Endangered Medicinal Plants from Abou Galoom Protectorate. Life Sci. J. 2015, 12, 101–109. [Google Scholar]
- El-Sakaty, S.I.; Magdy, M.; Rizk, S.; Hashim, A.; Abu-Elhamd, M.; Elateek, S. Ex-Situ Conservation of the Micro and Macro Flora of Omayed Biosphere Reserve (OBR): A Survey Report. Egypt. J. Bot. 2022, 62, 149–158. [Google Scholar] [CrossRef]
- Kress, W.J.; Erickson, D.L. DNA Barcodes: Genes, Genomics, and Bioinformatics. Proc. Natl. Acad. Sci. USA 2008, 105, 2761–2762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramana, K.V.; Trivedi, M.H.; Reddy, P.R.K.; Rao, C.V. Evaluation of Anti Ulcer Activity of Cordia Monoica Roxb Stem. Int. J. Pharm. Res. 2014, 2, 300–302. [Google Scholar]
- Olanlokun, J.O.; Bodede, O.; Prinsloo, G.; Olorunsogo, O.O. Comparative Antimalarial, Toxicity and Mito-Protective Effects of Diospyros Mespiliformis Hochst. ex A. DC. and Mondia Whitei (Hook.f.) Skeels on Plasmodium Berghei Infection in Mice. J. Ethnopharmacol. 2021, 268, 113585. [Google Scholar] [CrossRef]
- Luka, J.; Badau, S.J.; Mbaya, A.W.; Gadzama, J.J.; Kumshe, H.A. Acute Toxicity Study and Effect of Prolonged Administration (28 Days) of Crude Ethanolic Root Extract of Diospyros Mespiliformis Hochst (Ebenaceae) on Clinical, Haematological and Biochemical Parameters of Albino Rats. J. Ethnopharmacol. 2014, 153, 268–273. [Google Scholar] [CrossRef]
- Ebbo, A.A.; Sani, D.; Suleiman, M.M.; Ahmad, A.; Hassan, A.Z. Acute and Sub-Chronic Toxicity Evaluation of the Crude Methanolic Extract of Diospyros Mespiliformis Hochst Ex a. Dc (Ebenaceae) and Its Fractions. Toxicol. Rep. 2020, 7, 1138–1144. [Google Scholar] [CrossRef]
- Jahan, M.S.; Chowdhury, N.; Ni, Y. Effect of Different Locations on the Morphological, Chemical, Pulping and Papermaking Properties of Trema Orientalis (Nalita). Bioresour. Technol. 2010, 101, 1892–1898. [Google Scholar] [CrossRef]
- Farzana, M.; Rahman, M.M.; Ferdous, T.; Jahan, M.S. Review on Trema Orientalis as a Potential Bioresource in Tropical Countries. Trees 2022, 36, 1169–1177. [Google Scholar] [CrossRef]
- Olajide, J.E.; Sanni, M.; Achimugu, O.J.; Suleiman, M.S.; Jegede, E.R.; Sheneni, V.D. Effect of Methanol Extract of Trema Orientalis Leaf on Some Biochemical and Histopathological Indices of Wistar Albino Rats with Cadmium-Induced—Hepatotoxicity. Sci. Afr. 2020, 10, e00568. [Google Scholar] [CrossRef]
- Bielecka, M.; Pencakowski, B.; Stafiniak, M.; Jakubowski, K.; Rahimmalek, M.; Gharibi, S.; Matkowski, A.; Ślusarczyk, S. Metabolomics and DNA-Based Authentication of Two Traditional Asian Medicinal and Aromatic Species of Salvia Subg. Perovskia. Cells 2021, 10, 112. [Google Scholar] [CrossRef] [PubMed]
- Duangjai, S.; Samuel, R.; Munzinger, J.; Forest, F.; Wallnöfer, B.; Barfuss, M.H.J.; Fischer, G.; Chase, M.W. A Multi-Locus Plastid Phylogenetic Analysis of the Pantropical Genus Diospyros (Ebenaceae), with an Emphasis on the Radiation and Biogeographic Origins of the New Caledonian Endemic Species. Mol. Phylogenet. Evol. 2009, 52, 602–620. [Google Scholar] [CrossRef] [PubMed]
- Botanic Gardens Conservation International. IUCN Red List of Threatened Species: Ficus Ingens. In IUCN Red List of Threatened Species; Botanic Gardens Conservation International: Richmond, UK, 2018; Available online: https://portals.iucn.org/library/efiles/documents/RL-2011-002.pdf (accessed on 5 June 2022).
- McLachlan, J.S.; Clark, J.S.; Manos, P.S. Molecular Indicators of Tree Migration Capacity under Rapid Climate Change. Ecology 2005, 86, 2088–2098. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
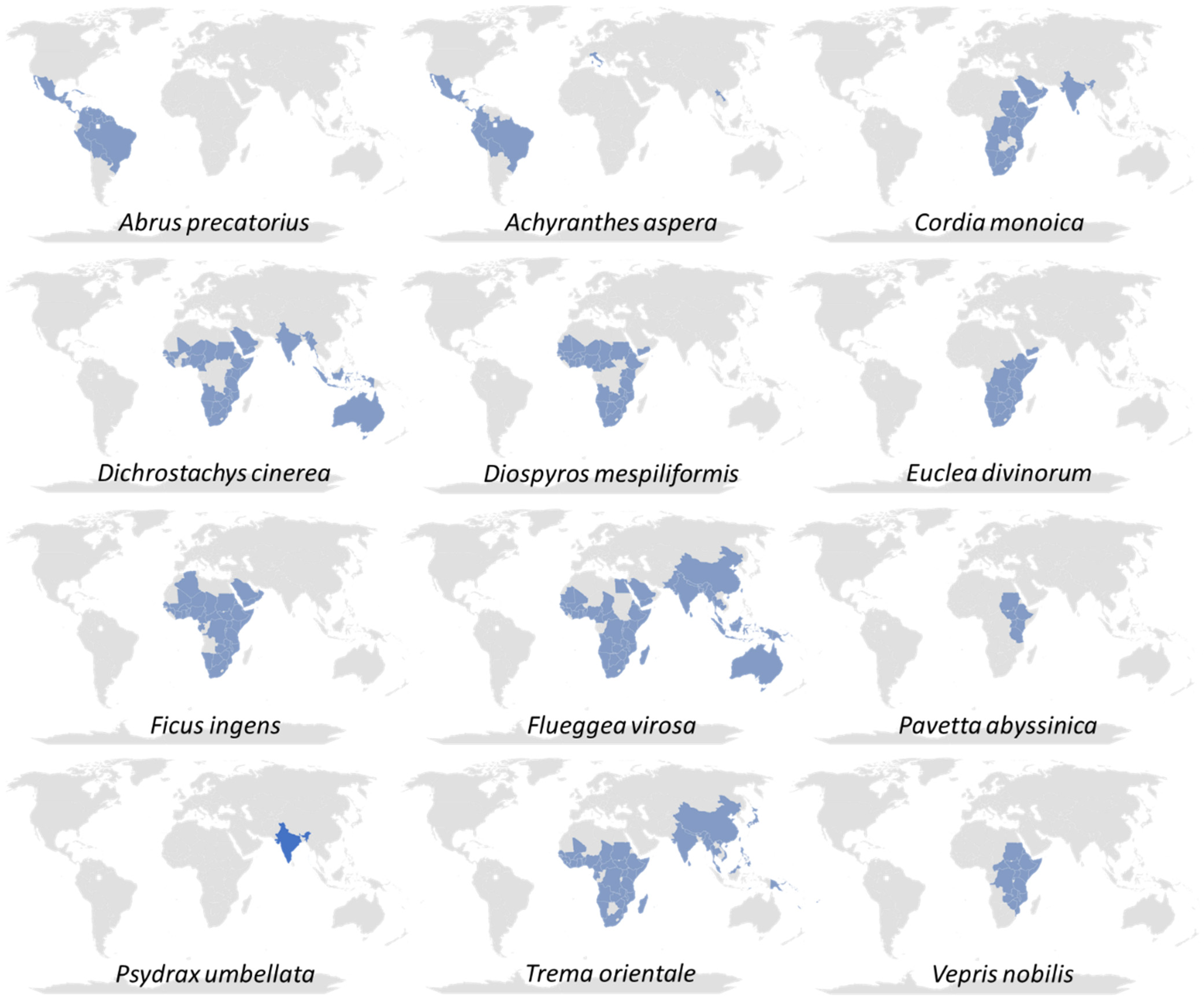{kind=link}
| Species | GC | Organism | Accession | Pairwise |
|---|---|---|---|---|
| Sp_01 | 43.50% | Diospyros mespiliformis | HF568789 | 100.00% |
| Sp_02 | 43.10% | Euclea divinorum | EU980790 | 99.80% |
| 43.10% | Euclea crispa | EU980789 | 99.80% | |
| Sp_03 | 42.80% | Achyranthes longifolia | MZ351434 | 99.50% |
| 42.80% | Achyranthes bidentata | MT955652 | 99.50% | |
| 42.80% | Achyranthes aspera | MW411185 | 99.50% | |
| Sp_04 | 42.50% | Cordia monoica | KR737383 | 100.00% |
| Sp_05 | 43.40% | Psydrax obovata | KY378666 | 99.50% |
| Sp_06 | 44.30% | Pavetta abyssinica | KY378673 | 99.80% |
| Sp_07 | 44.30% | Vepris reflexa | JX573080 | 99.00% |
| Sp_08 | 43.60% | Flueggea virosa | NC_051502 | 100.00% |
| Sp_09 | 42.60% | Abrus precatorius | KF432060 | 99.80% |
| Sp_10 | 43.60% | Dichrostachys cinerea | MN591166 | 98.70% |
| Sp_11 | 43.80% | Trema tomentosum | KY931669 | 99.30% |
| Sp_12 | 42.60% | Ficus squamosa | OK077764 | 99.80% |
| 42.60% | Ficus sarmentosa | NC_061976 | 99.80% | |
| 42.60% | Ficus pumila | NC_058617 | 99.80% | |
| 42.60% | Ficus oligodon | OK078619 | 99.80% |
| Species | GC | Organism | Accession | Pairwise |
|---|---|---|---|---|
| Sp_01 | 47.90% | Diospyros lotus | MN722015 | 100 |
| Sp_02 | 66.00% | Euclea divinorum | KU378851 | 98.60% |
| Sp_03 | 52.60% | Achyranthes aspera | LT993006 | 99.70% |
| Sp_04 | 56.10% | Cordia monoica | MK261116 | 100.00% |
| Sp_05 | 65.10% | Psydrax umbellata | MT131263 | 100.00% |
| Sp_06 | 66.00% | Pavetta hongkongensis | KP092797 | 98.90% |
| Sp_07 | 58.50% | Vepris nobilis | KY508613 | 100.00% |
| Sp_08 | 56.20% | Flueggea virosa | MN915825 | 100.00% |
| Sp_09 | 53.20% | Abrus precatorius | AF467015 | 100.00% |
| Sp_10 | 62.30% | Dichrostachys cinerea | AF458820 | 100.00% |
| Sp_11 | 61.60% | Trema orientalis | KY563221 | 98.70% |
| Sp_12 | 63.50% | Ficus cordata | KF850595 | 97.80% |
| 63.30% | Ficus virens | KJ845958 | 97.70% | |
| 63.60% | Ficus pseudoconcinna | KJ845946 | 97.40% |
| ID * | Morphology | rbcL | PI% | ITS | PI% |
|---|---|---|---|---|---|
| Sp_01 | Diospyros mespiliformis | Diospyros mespiliformis | 100 | Diospyros lotus | 100 |
| Sp_02 | Euclea racemosa | Euclea divinorum | 99.8 | Euclea divinorum | 98.6 |
| Sp_03 | Nuxia oppositifolia | Achyranthes longifolia | 99.5 | Achyranthes aspera | 99.7 |
| Sp_04 | Cordia monoica | Cordia monoica | 100 | Cordia monoica | 100 |
| Sp_05 | Psydrax schimperiana | Psydrax obovata | 99.5 | Psydrax umbellata | 100 |
| Sp_06 | Pavetta gardeniifolia | Pavetta abyssinica | 99.8 | Pavetta hongkongensis | 98.9 |
| Sp_07 | Vepris nobilis | Vepris reflexa | 99 | Vepris nobilis | 100 |
| Sp_08 | Flueggea virosa | Flueggea virosa | 100 | Flueggea virosa | 100 |
| Sp_09 | Abrus precatorius | Abrus precatorius | 99.8 | Abrus precatorius | 99.8 |
| Sp_10 | Dichrostachys cinerea | Dichrostachys cinerea | 98.7 | Dichrostachys cinerea | 100 |
| Sp_11 | Trema orientalis | Trema tomentosum | 99.3 | Trema orientalis | 98.7 |
| Sp_12 | Ficus ingens | Ficus squamosa | 99.8 | Ficus cordata | 97.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Safhi, F.A.; Alshamrani, S.M.; Fiteha, Y.G.; Abd El-Moneim, D. DNA Barcoding of Endangered and Rarely Occurring Plants in Faifa Mountains (Jazan, Saudi Arabia). Agriculture 2022, 12, 1931. https://doi.org/10.3390/agriculture12111931
Safhi FA, Alshamrani SM, Fiteha YG, Abd El-Moneim D. DNA Barcoding of Endangered and Rarely Occurring Plants in Faifa Mountains (Jazan, Saudi Arabia). Agriculture. 2022; 12(11):1931. https://doi.org/10.3390/agriculture12111931
Chicago/Turabian StyleSafhi, Fatmah Ahmed, Salha Mesfer Alshamrani, Yosur Gamal Fiteha, and Diaa Abd El-Moneim. 2022. "DNA Barcoding of Endangered and Rarely Occurring Plants in Faifa Mountains (Jazan, Saudi Arabia)" Agriculture 12, no. 11: 1931. https://doi.org/10.3390/agriculture12111931
APA StyleSafhi, F. A., Alshamrani, S. M., Fiteha, Y. G., & Abd El-Moneim, D. (2022). DNA Barcoding of Endangered and Rarely Occurring Plants in Faifa Mountains (Jazan, Saudi Arabia). Agriculture, 12(11), 1931. https://doi.org/10.3390/agriculture12111931