
QTL Mapping of Leaf Area Index and Chlorophyll Content Based on UAV Remote Sensing in Wheat

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Ground Data Collection
2.2.1. Determination of LAI
2.2.2. Determination of CC
2.3. UAV Image Acquisition and Preprocessing
2.4. Selection of Vegetation Index
2.5. Algorithm Development for LAI and CC Estimation
2.6. Data Analysis
2.7. QTL Mapping of LAI and CC and Candidate Gene Identification
3. Results
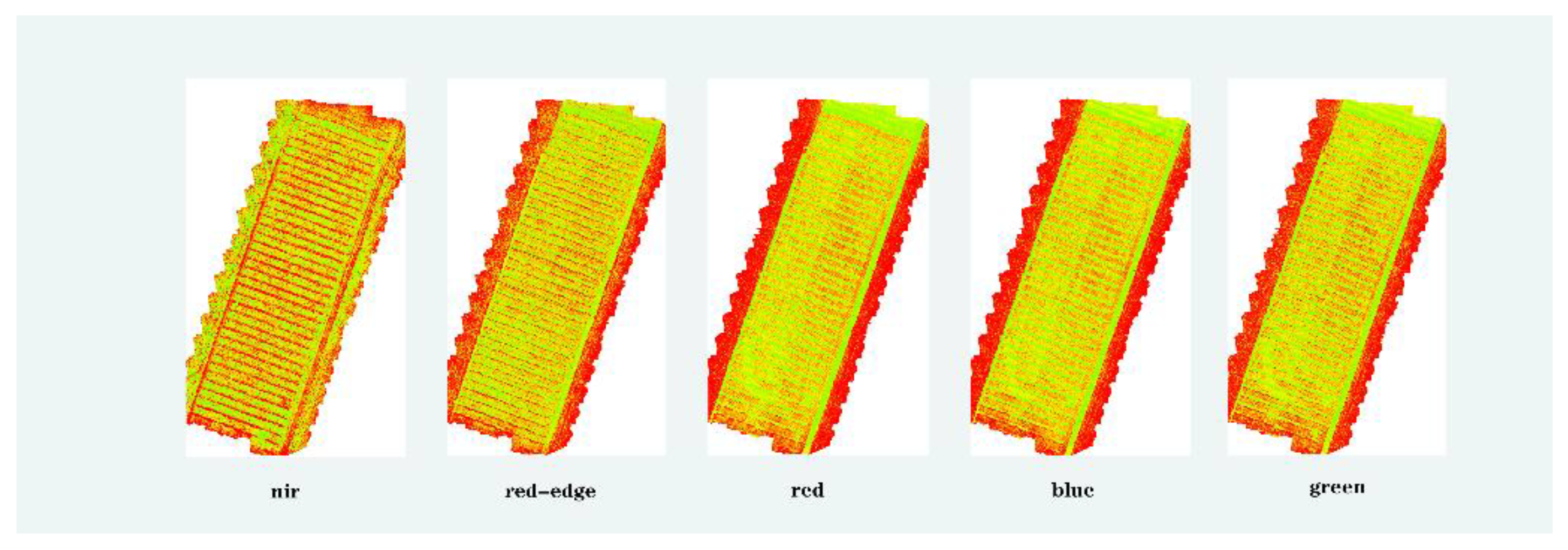
3.1. Multispectral Analysis of Reflectance
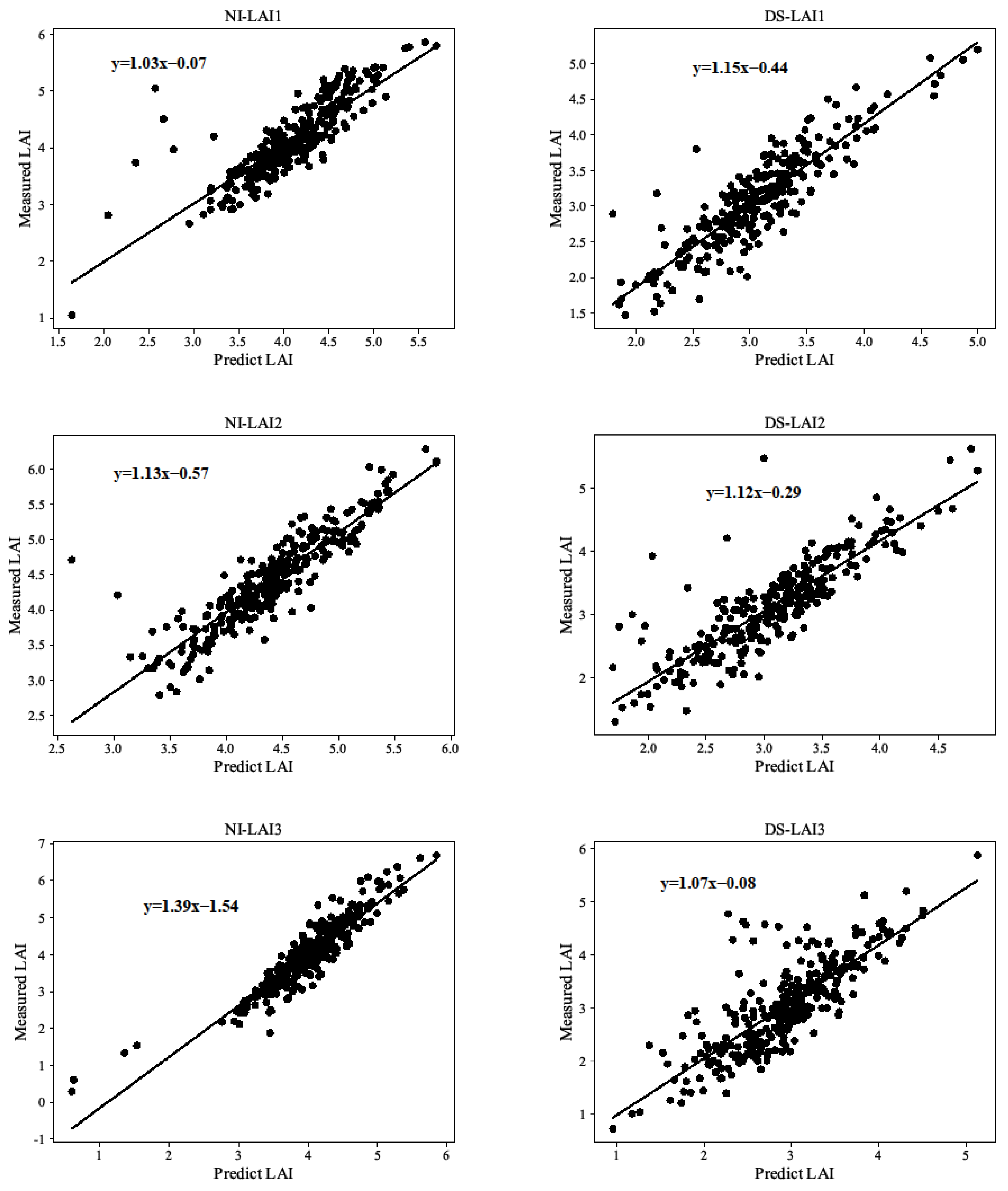
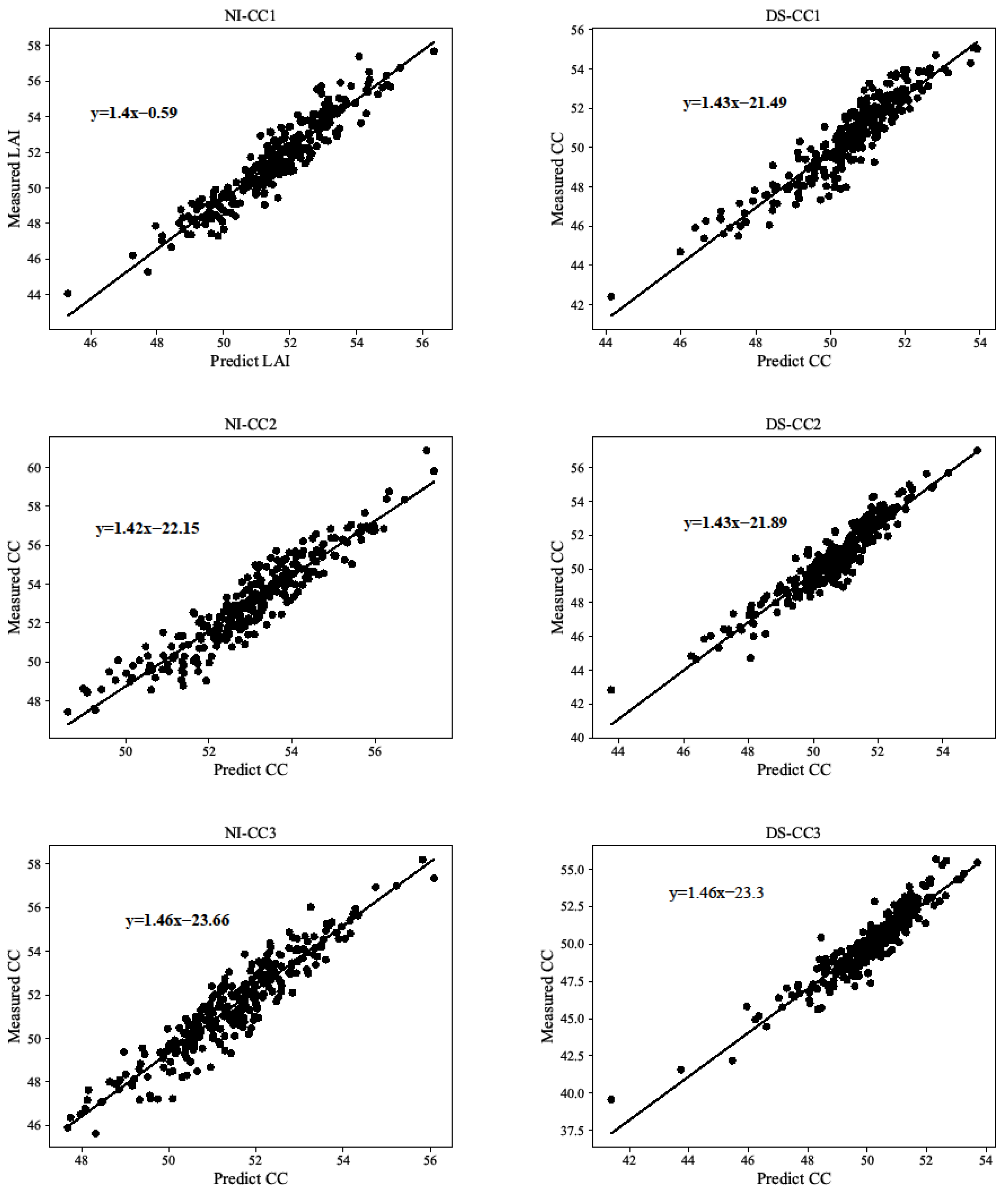
3.2. Estimation of LAI and CC by CART Modeling Verification Analysis
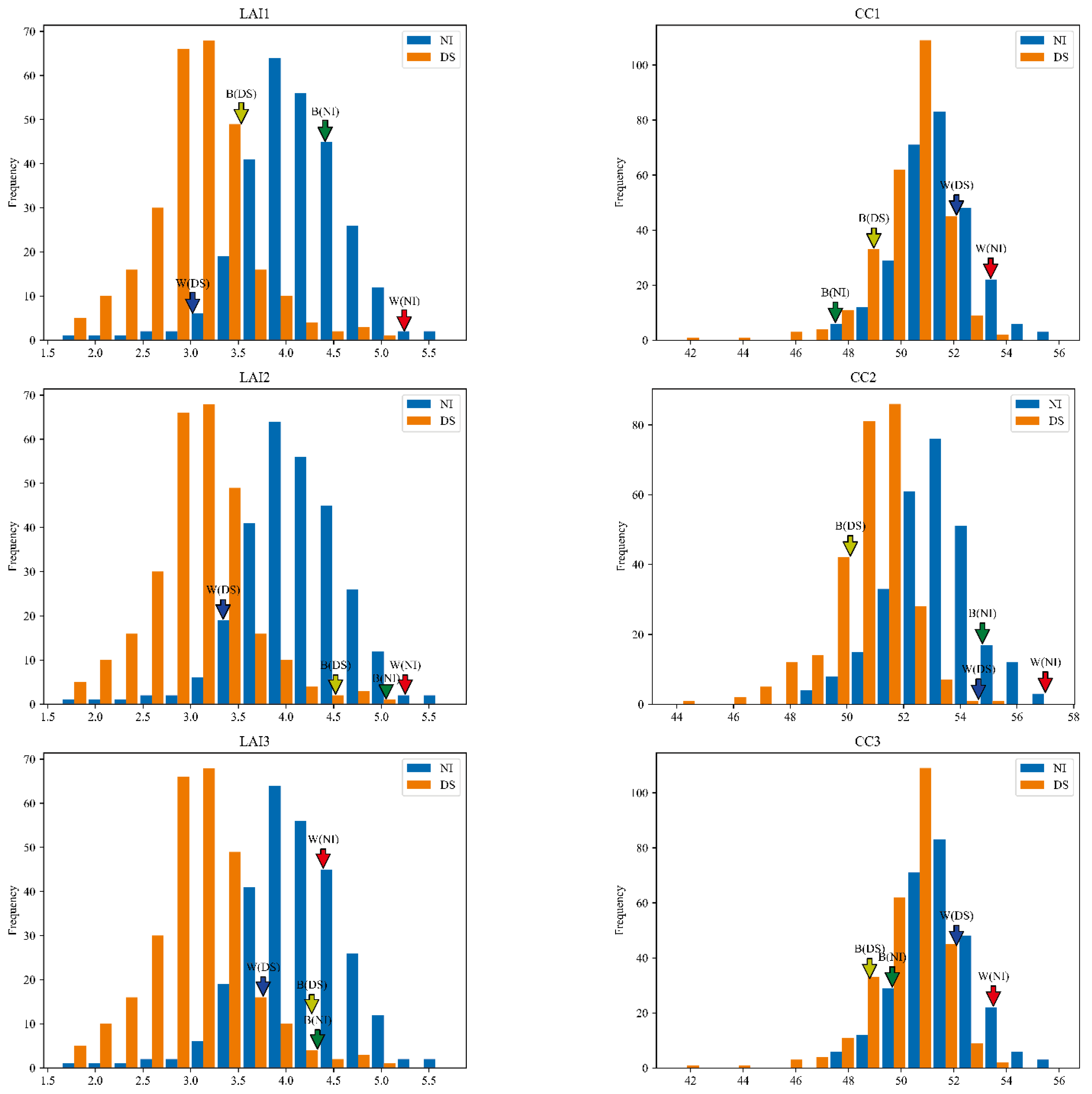
3.3. Genetic Variation of LAI and CC in Wheat RIL Population
3.4. Genetic Variation of Predicted LAI and CC in Wheat RIL Population
3.5. QTL Mapping of LAI and CC
3.6. Candidate Gene Mining and Comparative Analysis of Predicted and Measured Values
4. Discussion
4.1. Spectral Reflectance Difference Analysis
4.2. LAI and CC Modeling Verification
4.3. QTLs and Candidate Genes Association with Predicted and Measured Values of LAI and CC
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shiferaw, B.; Smale, M.; Braun, H.J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Secur. 2013, 5, 291–317. [Google Scholar] [CrossRef] [Green Version]
- Hassan, M.A.; Yang, M.; Rasheed, A.; Jin, X.; Xia, X.; Xiao, Y.; He, Z. Time-series multispectral indices from unmanned aerial vehicle imagery reveal senescence rate in bread wheat. Remote Sens. 2018, 10, 809. [Google Scholar] [CrossRef] [Green Version]
- Zampieri, M.; Ceglar, A.; Dentener, F.; Toreti, A. Wheat yield loss attributable to heat waves, drought and water excess at the global, national and subnational scales. Environ. Res. Lett. 2017, 12, 064008. [Google Scholar] [CrossRef]
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84. [Google Scholar] [CrossRef] [PubMed]
- Sean, M.; Ames, H.D.; Holshouser, D.L. Relationship between leaf area index and yield in double-crop and full-season soybean systems. J. Econ. Entomol. 2002, 95, 945–951. [Google Scholar]
- Yao, X.; Wang, N.; Liu, Y.; Cheng, T.; Tian, Y.; Chen, Q.; Zhu, Y. Estimation of wheat LAI at middle to high levels using unmanned aerial vehicle narrowband multispectral imagery. Remote Sens. 2017, 9, 1304. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Guo, Z.; Huang, C.; Duan, L.; Chen, G.; Jiang, N.; Fang, W.; Feng, H.; Xie, W.; Lian, X.; et al. Combining high-throughput phenotyping and genome-wide association studies to reveal natural genetic variation in rice. Nat. Commun. 2014, 5, 5087. [Google Scholar] [CrossRef]
- Jonckheere, I.; Fleck, S.; Nackaerts, K.; Muys, B.; Coppin, P.; Weiss, M.; Baret, F. Review of methods for in situ leaf area index determination: Part I. Theories, sensors and hemispherical photography. Agric. For. Meteorol. 2004, 121, 19–35. [Google Scholar] [CrossRef]
- Wang, X.Z.; Huang, J.F.; Li, Y.M.; Wang, R.C. Study on hyperspectral and red edge characteristics of rice under different nitrogen supply levels. J. Remote Sens. 2004, 8, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Adão, T.; Hruška, J.; Pádua, L.; Bessa, J.; Peres, E.; Morais, R.; Sousa, J.J. Hyperspectral imaging: A review on UAV-based sensors, data processing and applications for agriculture and forestry. Remote Sens. 2017, 9, 1110. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Yang, G.J.; Li, C.C.; Fang, H.K.; Xu, B.; Wang, L.; Dong, J.H.; Fu, K. Application of an improved method in retrieving leaf area index combined spectral index with PLSR in hyperspectral data generated by unmanned aerial vehicle snapshot camera. Acta Agron. Sin. 2017, 43, 549–557. [Google Scholar] [CrossRef]
- Verger, A.; Vigneau, N.; Chéron, C.; Gilliot, J.M.; Comar, A.; Baret, F. Green area index from an unmanned aerial system over wheat and rapeseed crops. Remote Sens. Environ. 2014, 152, 654–664. [Google Scholar] [CrossRef]
- Jin, Y.H.; Xiong, H.G.; Zhang, F.; Wang, L.F. Comparison of hyperspectral estimation models for chlorophyll content in spring wheat at jointing stage between irrigated and dryland. Agric. Res. Arid. Areas 2014, 32, 106–111. (In Chinese) [Google Scholar]
- Gong, Y.; Yang, K.; Lin, Z.; Fang, S.; Wu, X.; Zhu, R.; Peng, Y. Remote estimation of leaf area index (LAI) with unmanned aerial vehicle (UAV) imaging for different rice cultivars throughout the entire growing season. Plant Methods 2021, 17, 1–16. [Google Scholar] [CrossRef]
- Shibayama, M.; Akiyama, T. Seasonal visible, nir and mid-infrared spectra of rice canopies in relation to LAI and above-ground dry phytomass. Remote Sens. Environ. 1989, 27, 119–127. [Google Scholar] [CrossRef]
- Hunt, E.R.; Hively, W.D.; Fujikawa, S.J.; Linden, D.S.; Daughtry, C.S.; McCarty, G.W. Acquisition of NIR-green-blue digital photographs from unmanned aircraft for crop monitoring. Remote Sens. 2010, 2, 290–305. [Google Scholar] [CrossRef] [Green Version]
- Singhal, G.; Bansod, B.; Mathew, L.; Goswami, J.; Choudhury, B.U.; Raju, P. Estimation of eaf chlorophyll concentration in turmeric (Curcuma longa) using high-resolution unmanned aerial vehicle imagery based on kernel ridge regression. J. Indian Soc. Remote Sens. 2019, 47, 1111–1122. [Google Scholar] [CrossRef]
- Kaivosoja, J.; Pesonen, L.; Kleemola, J.; Pölönen, I.; Salo, H.; Honkavaara, E.; Saari, H.; Mäkynen, J.; Rajala, A. A case study of a precision fertilizer application task generation for wheat based on classified hyperspectral data from UAV combined with farm history data. In Proceedings of the SPIE—The International Society for Optical Engineering, San Diego, CA, USA, 9–12 September 2013; p. 8887. [Google Scholar]
- Condorelli, G.E.; Maccaferri, M.; Newcomb, M.; Andrade-Sanchez, P.; White, J.W.; French, A.N.; Sciara, G.; Ward, R.; Tuberosa, R. Comparative Aaerial and ground based high throughput phenotyping for the genetic dissection of NDVI as a proxy for drought adaptive traits in durum wheat. Front. Plant Sci. 2018, 9, 893. [Google Scholar] [CrossRef]
- Adeel, H.M.; Yang, M.J. Accuracy assessment of plant height using an unmanned aerial vehicle for quantitative genomic analysis in bread wheat. Plant Method 2019, 15, 1–12. [Google Scholar]
- Kumar, S.; Sehgal, S.K.; Kumar, U.; Prasad, P.V.; Joshi, A.K.; Gill, B.S. Genomic characterization of drought tolerance-related traits in spring wheat. Euphytica 2012, 186, 265–276. [Google Scholar] [CrossRef]
- Wang, S.X.; Zhu, Y.L.; Zhang, D.X.; Shao, H.; Liu, P.; Hu, J.B.; Zhang, H.; Zhang, H.P.; Chang, C.; Lu, J.; et al. Genome-wide association study for grain yield and related traits in elite wheat varieties and advanced lines using SNP markers. PLoS ONE 2017, 12, e0188662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Y.; Liu, J.D.; Zhang, J.X.; Dreisigacker, S.; Xia, X.C.; Geng, H.W. QTL mapping of drought tolerance at germination stage in wheat using the 50K SNP array. Plant Genet. Resour. Charact. Util. 2021, 19, 453–460. [Google Scholar] [CrossRef]
- Netto, A.T.; Campostrini, E.; de Oliveira, J.G.; Bressan-Smith, R.E. Photosynthetic pigments, nitrogen, chlorophyll a fluorescence and SPAD-502 readings in coffee leaves. Sci. Hortic. 2005, 104, 199–209. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Breiman, L.; Friedman, J.H.; Olshen, R.A.; Stone, C.J. Classification and Regression Trees (CART). Biometrics 1984, 40, 358. [Google Scholar]
- Saptoro, A.; Tadé, M.O.; Vuthaluru, H. A modified Kennard-Stone algorithm for optimal division of data for developing artificial neural network models. Chem. Prod. Process Model. 2012, 7, 16. [Google Scholar] [CrossRef]
- Zhang, J.; Cheng, T.; Guo, W.; Xu, X.; Qiao, H.; Xie, Y.; Ma, X. Leaf area index estimation model for UAV image hyperspectral data based on wavelength variable selection and machine learning methods. Plant Methods 2021, 17, 1–4. [Google Scholar] [CrossRef]
- Gitelson, A.A. Remote estimation of canopy chlorophyll content in crops. Geophys. Res. Lett. 2005, 32, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Yang, J.; Guo, L.; Hu, P.; Liu, X.; Huang, P.; Wang, C. A bionic point-source polarisation sensor applied to underwater orientation. J. Navig. 2021, 74, 1–16. [Google Scholar] [CrossRef]
- Horler, D.N.; Dockray, M.; Barber, J. The red edge of plant leaf reflectance. Int. J. Remote Sens. 1983, 4, 273–288. [Google Scholar] [CrossRef]
- Berger, B.; Parent, B.; Tester, M. High-throughput shoot imaging to study drought responses. J. Exp. Bot. 2010, 61, 3519–3528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houborg, R.; McCabe, M.F. A hybrid training approach for leaf area index estimation via Cubist and random forests machine-learning. ISPRS J. Photogramm. 2018, 135, 173–188. [Google Scholar] [CrossRef]
- Yang, T.; Gao, X.; Sorooshian, S.; Li, X. Simulating California reservoir operation using the classification and regression-tree algorithm combined with a shuffled cross-validation scheme. Water Resour. Res. 2016, 52, 1626–1651. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Yang, G.; Yu, H.; Xu, B.; Zhao, X.; Dong, J.; Ma, Y. Retrieving winter wheat leaf area index based on unmanned aerial vehicle hyperspectral remote sensing. Trans. Chin. Soc. Agric. Eng. 2016, 32, 113–120. [Google Scholar]
- Jin, X.; Liu, S.; Baret, F.; Hemerlé, M.; Comar, A. Estimates of plant density of wheat crops at emergence from very low altitude UAV imagery. Remote Sens. Environ. 2017, 198, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Hu, P.; Chapman, S.C.; Wang, X.; Potgieter, A.; Duan, T.; Jordan, D.; Guo, Y.; Zheng, B. Estimationof plant height using a high throughput phenotyping platform based on unmanned aerial vehicle and self-calibration: Example for sorghum breeding. Eur. J. Agron. 2018, 95, 24–32. [Google Scholar] [CrossRef]
- Maccaferri, M.; Sanguineti, M.C.; Corneti, S.; Ortega, J.L.; Salem, M.B.; Bort, J.; DeAmbrogio, E.; del Moral, L.F.; Demontis, A.; El-Ahmed, A.; et al. Quantitative trait loci for grain yield and adaptation of durum wheat (Triticum durum Desf.) across a wide range of water availability. Genetics 2008, 178, 489–511. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhao, L.; Dong, M.; Ren, Y.; Zhang, L.; Chen, F. QTL localization of wheat plant height and flag leaf-related traits. J. Wheat Crop. 2019, 39, 761–767. (In Chinese) [Google Scholar]
- Hu, Y.S.; Ren, T.H.; Li, Z.; Tang, Y.Z.; Ren, Z.L.; Yan, B.J. Molecular mapping and genetic analysis of a QTL controlling spike formation rate and tiller number in wheat. Gene 2017, 634, 15. [Google Scholar] [CrossRef]
- Zhao, P.; Xu, F.; Jiang, W.H.; Qi, P.; Li, C.L.; Bai, H.B.; Lv, X.L.; Dong, J.L.; Wang, Z.H. QTL analysis of flag leaf length, width and chlorophyll content in spring wheat. J. Wheat Crop. 2015, 35, 603–608. (In Chinese) [Google Scholar]
- Lyzenga, W.J.; Stone, S.L. Abiotic stress tolerance mediated by protein ubiquitination. J. Exp. Bot. 2012, 63, 599–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kipreo, S.E.T.; Pagano, M. The F -box protein family. Genome Biol. 2000, 1, 1–7. [Google Scholar]
- Sofo, A.; Dichio, B.; Xiloyannis, C.; Masia, A. Lipoxygenase activity and proline accumulation in leaves and roots of olive trees in response to drought stress. Physiol. Plant 2010, 121, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.B.; Liang, Z.S.; Shao, M.A. Osmotic regulation of 10 wheat (Triticum aestivum L.) genotypes at soil water deficits. Colloids Surf. Biointerfaces 2006, 47, 132–139. [Google Scholar]
- Yin, M.; Wang, Y.; Zhang, L.; Li, J.; Quan, W.; Yang, L.; Wang, Q.; Chan, Z. The Arabidopsis Cys2/His2 zinc finger transcription factor ZAT18 is a positive regulator of plant tolerance to drought stress. J. Exp. Bot. 2017, 68, 2991–3005. [Google Scholar] [CrossRef]
- Weiss, D.; Ori, N. Mechanisms of cross talk between gibberellin and other hormones. Plant Physiol. 2007, 144, 1240–1246. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Kim, S.H.; Seo, D.H.; Chung, S.; Kim, S.W.; Lee, J.S.; Kim, W.T.; Lee, J.H. ABA-HYPERSENSITIVE BTB/POZ PROTEIN 1 functions as a negative regulator in ABA-mediated inhibition of germination in Arabidopsis. Plant Mol. Biol. 2016, 90, 303–315. [Google Scholar] [CrossRef]
- Gao, Z.; Chen, Y.F.; Randlett, M.D.; Zhao, X.C.; Findell, J.L.; Kieber, J.J.; Schaller, G.E. Localization of the Raf -like kinase CTR1 to the endoplasmic reticulum of Arabidopsis through participation in ethylene receptor signaling complexes. J. Biol. Chem. 2003, 278, 34725–34732. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water Condition | Bands | Parameters | Heading | Flowering | Filling | Maturity |
|---|---|---|---|---|---|---|
| DI | NIR | Mean | 0.37 | 0.37 | 0.37 | 0.26 |
| SD | 0.03 | 0.03 | 0.03 | 0.03 | ||
| Red-edge | Mean | 0.28 | 0.28 | 0.28 | 0.26 | |
| SD | 0.03 | 0.03 | 0.03 | 0.02 | ||
| Green | Mean | 0.07 | 0.07 | 0.07 | 0.09 | |
| SD | 0.01 | 0.01 | 0.01 | 0.01 | ||
| Red | Mean | 0.05 | 0.05 | 0.05 | 0.10 | |
| SD | 0.01 | 0.01 | 0.01 | 0.02 | ||
| Blue | Mean | 0.04 | 0.04 | 0.04 | 0.04 | |
| SD | 0.01 | 0.01 | 0.01 | 0.01 | ||
| DS | NIR | Mean | 0.33 | 0.32 | 0.30 | 0.28 |
| SD | 0.03 | 0.03 | 0.02 | 0.02 | ||
| Red-edge | Mean | 0.28 | 0.29 | 0.27 | 0.30 | |
| SD | 0.02 | 0.02 | 0.02 | 0.02 | ||
| Green | Mean | 0.08 | 0.09 | 0.10 | 0.11 | |
| SD | 0.01 | 0.02 | 0.02 | 0.01 | ||
| Red | Mean | 0.07 | 0.08 | 0.11 | 0.14 | |
| SD | 0.02 | 0.02 | 0.04 | 0.03 | ||
| Blue | Mean | 0.04 | 0.06 | 0.05 | 0.05 | |
| SD | 0.01 | 0.03 | 0.01 | 0.01 |
| Traits | Growth Stage | Treatment | R2 | RMSE | RE |
|---|---|---|---|---|---|
| LAI | Flowering stage (LAI1) | NI | 0.70 | 0.36 | 0.08 |
| DS | 0.78 | 0.32 | 0.10 | ||
| Filling stage (LAI2) | NI | 0.81 | 0.30 | 0.06 | |
| DS | 0.73 | 0.39 | 0.12 | ||
| Mature stage (LAI3) | NI | 0.84 | 0.44 | 0.11 | |
| DS | 0.64 | 0.52 | 0.18 | ||
| CC | Heading stage (CC1) | NI | 0.88 | 0.99 | 0.01 |
| DS | 0.86 | 0.97 | 0.01 | ||
| Flowering stage (CC2) | NI | 0.88 | 1.02 | 0.01 | |
| DS | 0.88 | 0.94 | 0.01 | ||
| Filling stage (CC3) | NI | 0.88 | 1.00 | 0.01 | |
| DS | 0.87 | 1.05 | 0.02 |
| Traits | Growth Stage | Treatment | Parent | RILs | |||||
|---|---|---|---|---|---|---|---|---|---|
| Worrakatta | Berkut | Mean | SD | Range | CV (%) | h2 (%) | |||
| LAI | Flowering stage | NI | 4.57 | 4.39 | 4.21 | 0.63 | 2.66~5.86 | 15.4 | 70 |
| DS | 4.6 | 3.58 | 3.07 ** | 0.64 | 1.62~5.08 | 21 | 70 | ||
| Filling stage | NI | 5.32 | 4.57 | 4.39 | 0.66 | 2.79~6.29 | 14.9 | 73 | |
| DS | 4.42 | 3.8 | 2.94 ** | 0.65 | 1.31~4.67 | 22.2 | 69 | ||
| Mature stage | NI | 4.57 | 3.95 | 3.85 | 0.77 | 2.24~6.05 | 19.9 | 73 | |
| DS | 4.26 | 3.65 | 2.91 ** | 0.82 | 1.01~5.00 | 28.2 | 75 | ||
| CC | Heading stage | NI | 53.38 | 46.11 | 51.63 | 2.32 | 44.06~57.68 | 4.5 | 81 |
| DS | 53.41 | 48.9 | 50.76 ** | 1.99 | 42.41~55.08 | 3.9 | 75 | ||
| Flowering stage | NI | 57.68 | 48.28 | 53.1 | 2.33 | 47.43~60.86 | 4.4 | 78 | |
| DS | 55.54 | 47.68 | 51.19 ** | 2.16 | 44.52~57.08 | 4.2 | 76 | ||
| Filling stage | NI | 54.82 | 46.48 | 51.31 | 2.11 | 45.62~56.99 | 4.1 | 74 | |
| DS | 54.08 | 47.06 | 50.34 ** | 2.07 | 39.57~55.67 | 4.1 | 74 | ||
| Traits | Growth Stage | Treatment | Parent | RIL | |||||
|---|---|---|---|---|---|---|---|---|---|
| Worrakatta | Berkut | Mean | SD | Range | CV (%) | h2 (%) | |||
| LAI | Flowering stage | NI | 5.24 | 4.41 | 4.1 | 0.47 | 1.63–5.70 | 11.5 | 57 |
| DS | 3.02 | 3.53 | 3.06 ** | 0.44 | 1.85–4.87 | 14.5 | 58 | ||
| Filling stage | NI | 5.25 | 5.05 | 4.4 | 0.48 | 3.31–5.87 | 10.8 | 65 | |
| DS | 3.34 | 4.52 | 3.06 ** | 0.5 | 1.71–4.84 | 16.4 | 58 | ||
| Mature stage | NI | 4.39 | 4.36 | 4.01 | 0.63 | 0.60–5.85 | 15.6 | 93 | |
| DS | 3.76 | 4.27 | 2.92 ** | 0.62 | 0.95–4.51 | 21.4 | 62 | ||
| CC | Heading stage | NI | 53.59 | 47.2 | 51.62 | 1.54 | 45.31–56.33 | 3 | 80 |
| DS | 52.69 | 48.96 | 50.50 ** | 1.36 | 44.13–53.92 | 2.7 | 64 | ||
| Flowering stage | NI | 57.06 | 54.58 | 53 | 1.52 | 48.61–57.41 | 2.9 | 77 | |
| DS | 55.29 | 50.32 | 50.68 ** | 1.39 | 43.76–55.06 | 2.8 | 64 | ||
| Filling stage | NI | 53.5 | 49.66 | 51.44 | 1.41 | 47.67–56.09 | 2.7 | 70 | |
| DS | 50.6 | 48.46 | 50.20 ** | 1.43 | 41.38–53.70 | 2.8 | 67 | ||
| Traits | Growth Stage | Treatment | Predicted Values | Measured Values | Correlation | ||
|---|---|---|---|---|---|---|---|
| Range | Mean | Range | Mean | ||||
| LAI | Flowering stage | NI | 1.65–5.70 | 4.1 | 2.66–5.86 | 4.21 | 0.83 ** |
| DS | 1.80–5.00 | 3.06 | 1.62–5.08 | 3.07 | 0.88 ** | ||
| Filling stage | NI | 2.63–5.87 | 4.4 | 2.79–6.29 | 4.39 | 0.90 ** | |
| DS | 1.69–4.84 | 3.06 | 1.31–4.67 | 2.94 | 0.85 ** | ||
| Mature stage | NI | 0.60–5.85 | 4.01 | 2.24–6.05 | 3.85 | 0.92 ** | |
| DS | 0.95–5.13 | 2.92 | 1.01–5.00 | 2.91 | 0.80 ** | ||
| CC | Heading stage | NI | 45.31–56.33 | 51.62 | 44.06–57.68 | 51.63 | 0.94 ** |
| DS | 44.13–53.92 | 50.5 | 42.41–55.08 | 50.76 | 0.93 ** | ||
| Flowering stage | NI | 48.61–57.41 | 53 | 47.43–60.86 | 53.1 | 0.94 ** | |
| DS | 43.76–55.06 | 50.68 | 44.52–57.08 | 51.19 | 0.94 ** | ||
| Filling stage | NI | 47.67–56.09 | 51.44 | 45.62–56.99 | 51.31 | 0.94 ** | |
| DS | 41.38–53.70 | 50.2 | 39.57–55.67 | 50.34 | 0.93 ** | ||
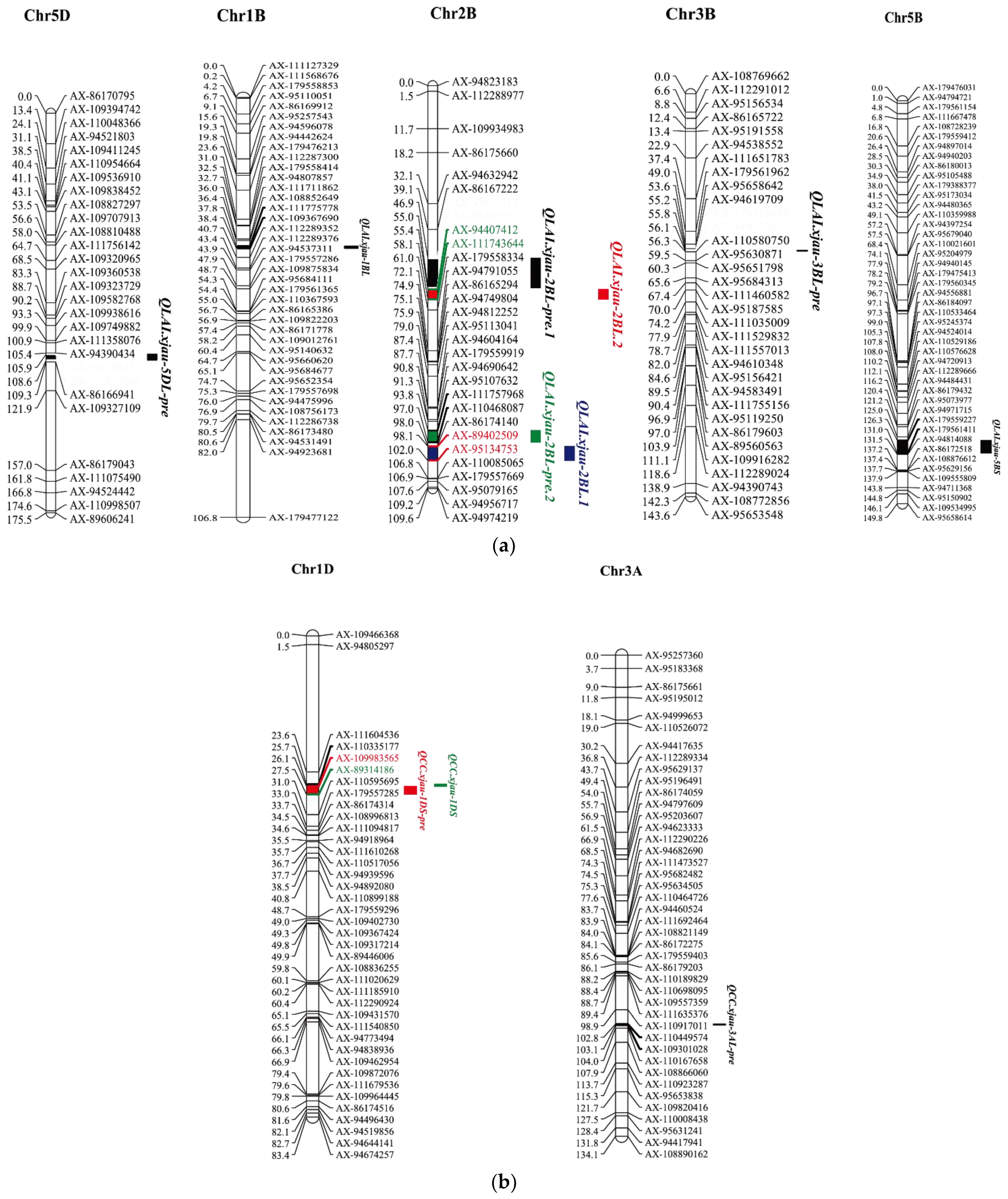
| Traits | Source | Growth Stage | Treatment | QTL Name | Chr. | Marker Interval | Position (cM) | LOD | R2 (%) | Additive |
|---|---|---|---|---|---|---|---|---|---|---|
| LAI | Predicted value | Flowering | NI | QLAI.xjau-2BL-pre.1 | 2BL | AX-89402509–AX-95134753 | 101 | 5.6 | 3.1 | 0.13 |
| Filling | NI | QLAI.xjau-5DL-pre | 5DL | AX-94670615–AX-111652649 | 108 | 5.7 | 3.6 | 0.13 | ||
| Filling | DS | QLAI.xjau-3BL-pre | 3BL | AX-179558689–AX-94903264 | 56 | 5.3 | 3.3 | 0.16 | ||
| Maturity | NI | QLAI.xjau-5DL-pre | 5DL | AX-94670615–AX-111652649 | 108 | 4.1 | 2.5 | 0.13 | ||
| Maturity | DS | QLAI.xjau-2BL-pre.2 | 2BL | AX-95631217–AX-94667571 | 54 | 5.3 | 3.3 | 0.16 | ||
| Measured value | Flowering | NI | QLAI.xjau-2BL.1 | 2BL | AX-111757968–AX-110468087 | 96 | 3.4 | 6.8 | 0.17 | |
| Flowering | NI | QLAI.xjau-5BS | 5BS | AX-179559227–AX-179561411 | 127 | 4.2 | 8.2 | 0.18 | ||
| Maturity | NI | QLAI.xjau-1BL | 1BL | AX-111775778–AX-109367690 | 38 | 2.8 | 7.6 | −0.19 | ||
| Maturity | DS | QLAI.xjau-2BL.2 | 2BL | AX-94407412–AX-111743644 | 56 | 4.6 | 13.8 | 0.3 | ||
| CC | Predicted value | Heading | NI | QCC.xjau-1DS-pre | 1DS | AX-109983565–AX-89314186 | 27 | 4.1 | 2.5 | 0.31 |
| Heading | NI | QCC.xjau-3AL-pre | 3AL | AX-110449574–AX-109301028 | 103 | 3.9 | 2.5 | -0.3 | ||
| Measured value | Heading | NI | QCC.xjau-1DS | 1DS | AX-110335177–AX-109983565 | 26 | 2.5 | 5.8 | 0.52 | |
| Flowering | NI | QCC.xjau-1DS | 1DS | AX-110335177–AX-109983565 | 26 | 3.2 | 5.3 | 0.57 |
| Source | Traits | QTL Name | Marker Interval | Physical Location (Mb) | Gene | Function Description |
|---|---|---|---|---|---|---|
| Predicted value | LAI | QLAI.xjau-2BL-pre.1 | AX-89402509– AX-95134753 | 797.209401–797.210138 | TraesCS2B01G623600 | F-box family protein |
| LAI | QLAI.xjau-2BL-pre.2 | AX-95631217– AX-94667571 | 555.852677–555.853828 | TraesCS2B01G391300 | F-box family protein | |
| LAI | QLAI.xjau-3BL-pre | AX-179558689– AX-94903264 | 81.813957–81.818009 | TraesCS3B01G115100 | Kelch-like protein | |
| LAI | QLAI.xjau-5DL-pre | AX-94670615– AX-111652649 | 473.052014–473.053148 | TraesCS5D01G408900 | Peroxidase | |
| LAI | QLAI.xjau-5DL-pre | AX-94670615– AX-111652649 | 474.813752–474.814753 | TraesCS5D01G411400 | C2H2-type zinc finger protein | |
| CC | QCC.xjau-1DS-pre | AX-109983565– AX-89314186 | 382.257141–382.258109 | TraesCS1D01G284100 | GATA transcription factor | |
| CC | QCC.xjau-3AL-pre | AX-110449574– AX-109301028 | 711.402978–711.407172 | TraesCS3A01G480600 | BTB/POZ domain- containing protein | |
| Measured value | LAI | QLAI.xjau-1BL | AX-111775778– AX-109367690 | 652.781869–652.786461 | TraesCS1B01G427400 | Wuschel homeobox protein |
| LAI | QLAI.xjau-2BL.1 | AX-111757968– AX-110468087 | 794.07325–794.082431 | TraesCS2B01G617700 | MYB-related transcription factor | |
| LAI | QLAI.xjau-2BL.2 | AX-94407412– AX-111743644 | 691.187478–691.1904 | TraesCS2B01G493800 | BTB/POZ domain-containing protein | |
| LAI | QLAI.xjau-2BL.2 | AX-94407412– AX-111743644 | 712.002368–712.004414 | TraesCS2B01G517100 | F-box family protein | |
| LAI | QLAI.xjau-5BS | AX-179559227– AX-179561411 | 19.455584–19.456986 | TraesCS5B01G019700 | F-box family protein | |
| CC | QCC.xjau-1DS | AX-110335177– AX-109983565 | 382.257141–382.258109 | TraesCS1D01G284100 | GATA transcription factor | |
| CC | QCC.xjau-1DS | AX-110335177– AX-109983565 | 392.69691–392.697515 | TraesCS1D01G293600 | Abscisic acid receptor |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Gao, X.; Cheng, Y.; Ren, Y.; Zhang, Z.; Wang, R.; Cao, J.; Geng, H. QTL Mapping of Leaf Area Index and Chlorophyll Content Based on UAV Remote Sensing in Wheat. Agriculture 2022, 12, 595. https://doi.org/10.3390/agriculture12050595
Wang W, Gao X, Cheng Y, Ren Y, Zhang Z, Wang R, Cao J, Geng H. QTL Mapping of Leaf Area Index and Chlorophyll Content Based on UAV Remote Sensing in Wheat. Agriculture. 2022; 12(5):595. https://doi.org/10.3390/agriculture12050595
Chicago/Turabian StyleWang, Wei, Xue Gao, Yukun Cheng, Yi Ren, Zhihui Zhang, Rui Wang, Junmei Cao, and Hongwei Geng. 2022. "QTL Mapping of Leaf Area Index and Chlorophyll Content Based on UAV Remote Sensing in Wheat" Agriculture 12, no. 5: 595. https://doi.org/10.3390/agriculture12050595
APA StyleWang, W., Gao, X., Cheng, Y., Ren, Y., Zhang, Z., Wang, R., Cao, J., & Geng, H. (2022). QTL Mapping of Leaf Area Index and Chlorophyll Content Based on UAV Remote Sensing in Wheat. Agriculture, 12(5), 595. https://doi.org/10.3390/agriculture12050595