
Genome-Wide Association Study Uncovers Loci and Candidate Genes Underlying Phytosterol Variation in Sesame (Sesamum indicum L.)

 ,
,  ,
, 
 and
and 
Abstract
:1. Introduction

2. Materials and Methods
2.1. Plant Materials
2.2. Phytosterol Content Analysis
2.3. Statistical Analysis
2.4. Genome-Wide Association Analysis
2.5. Identification of Candidate Gene and Haplotype Analysis
2.6. Gene Expression Analysis of Candidate Genes
3. Results
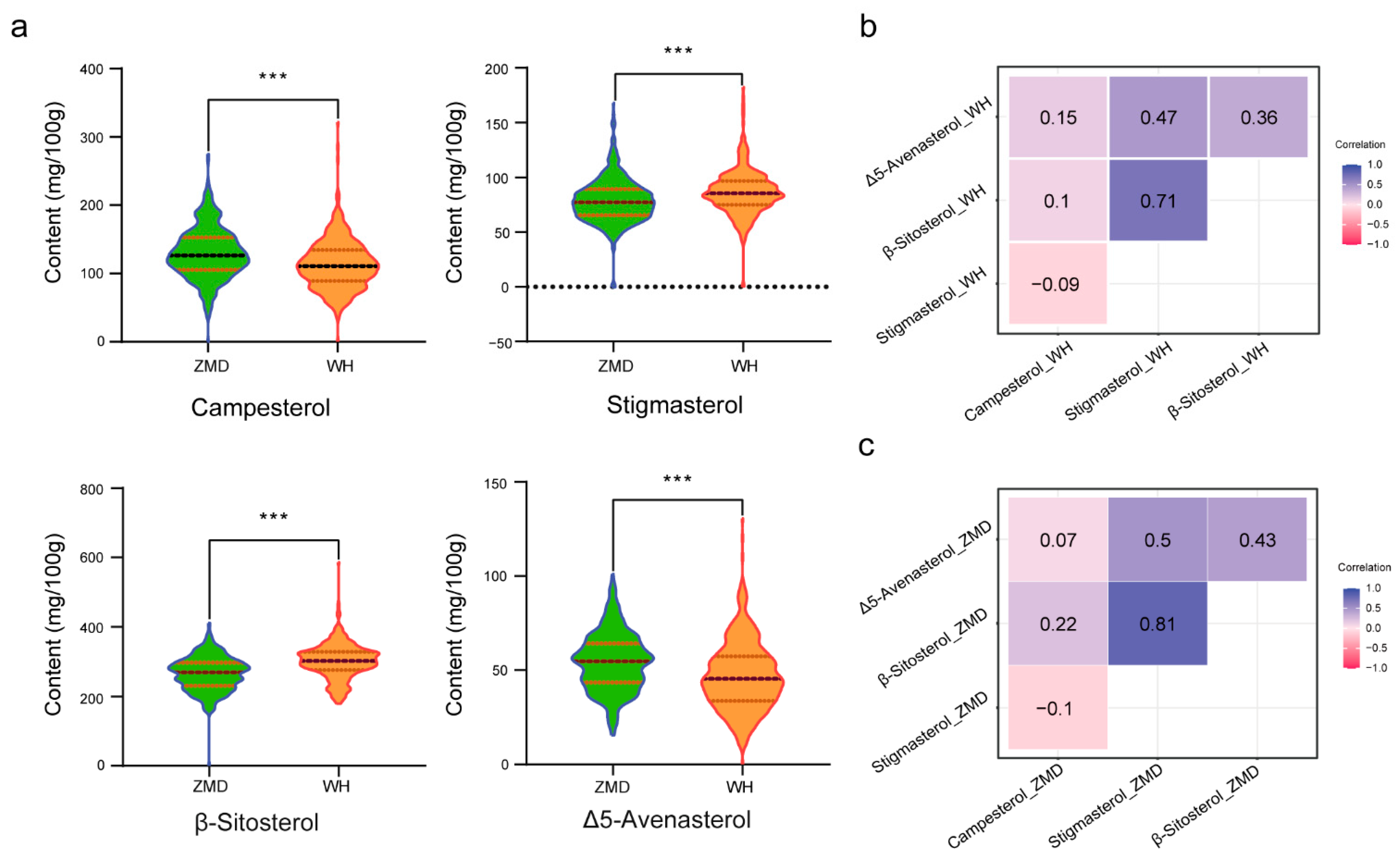
3.1. Phenotypic Variation and Correlation Analysis of Primary Phytosterols in Sesame
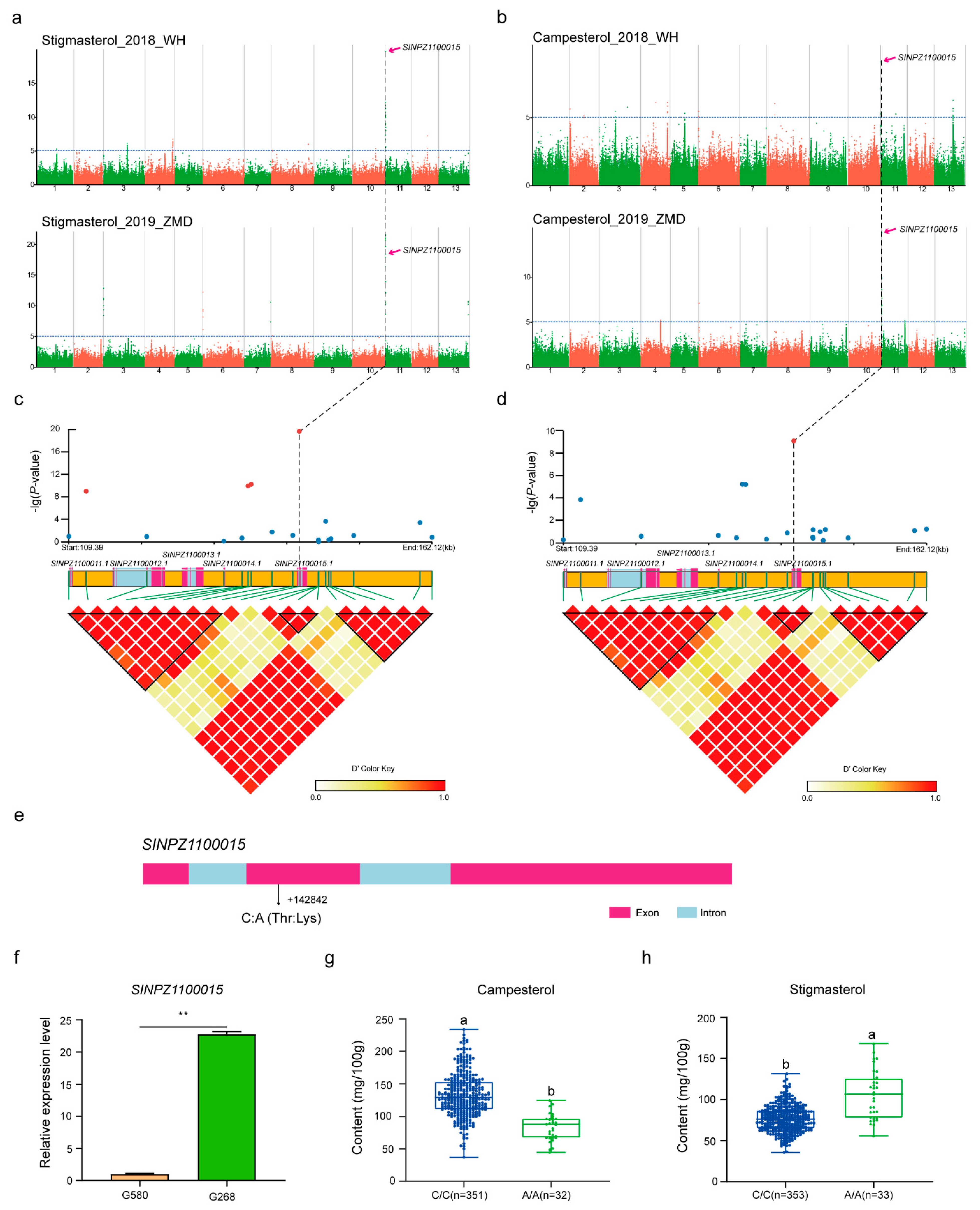
3.2. GWAS for Four Phytosterol Contents in Sesame
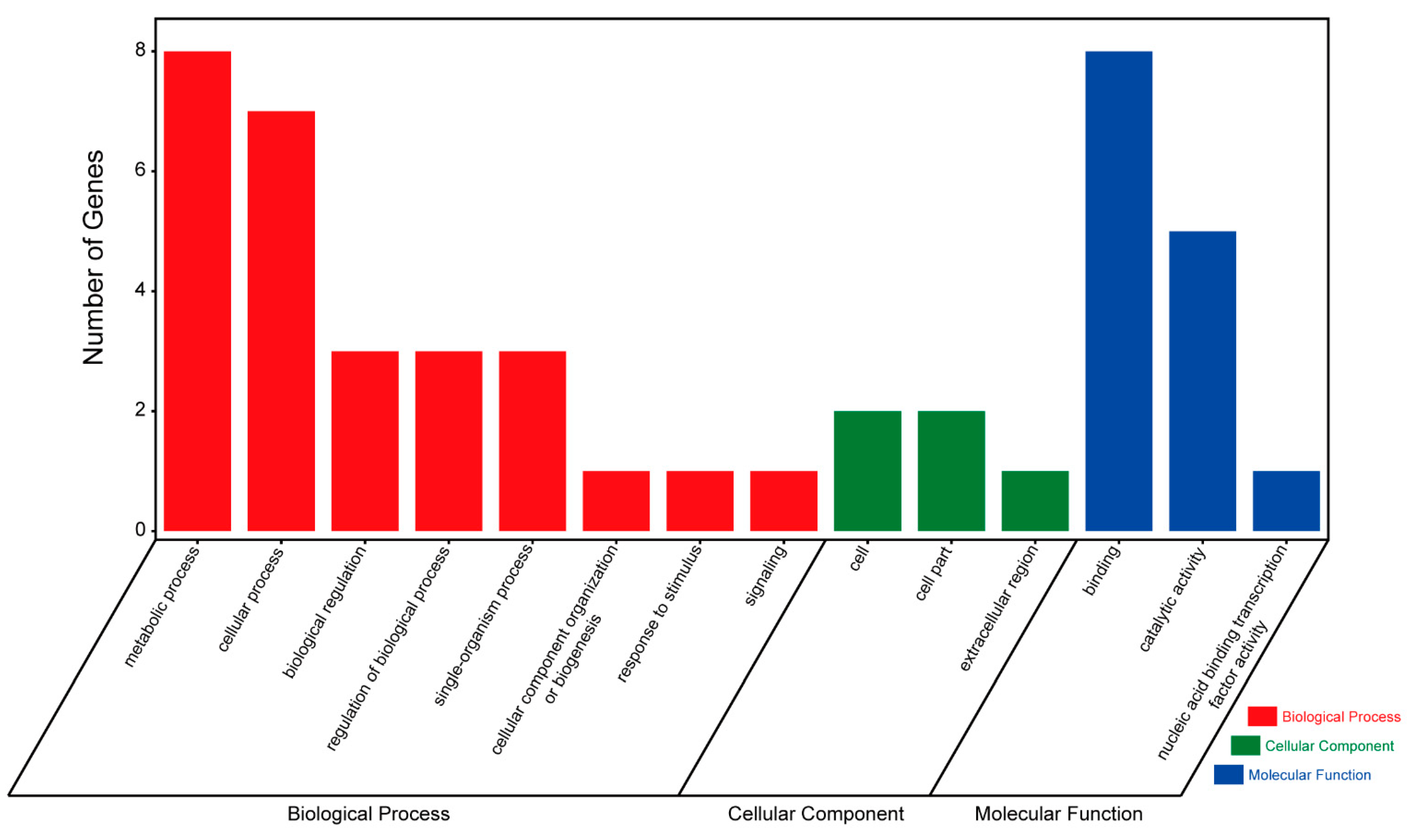
3.3. Candidate Genes Associated with Phytosterol Contents
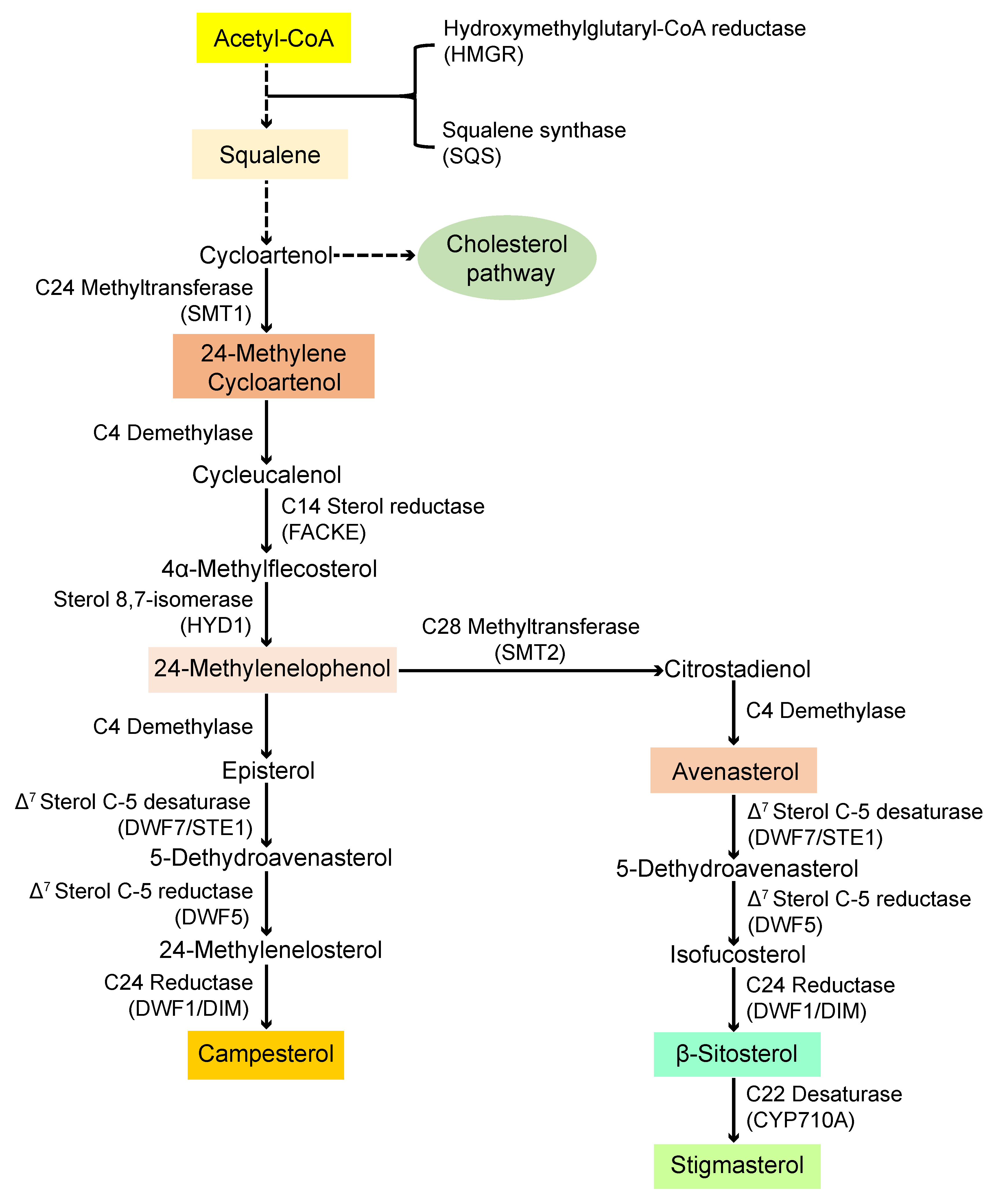
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Neelakandan, A.K.; Chamala, S.; Valliyodan, B.; Nes, W.D.; Nguyen, H.T. Metabolic engineering of soybean affords improved phytosterol seed traits. Plant Biotechnol. J. 2012, 10, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Tan, M.A.; An, S.S.A. Phytosterols: Potential metabolic modulators in neurodegenerative diseases. Int. J. Mol. Sci. 2021, 22, 12255. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Belwal, T.; Li, L.; Limwachiranon, J.; Liu, X.; Luo, Z. Phytosterols and their derivatives: Potential health-promoting uses against lipid metabolism and associated diseases, mechanism, and safety issues. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1243–1267. [Google Scholar] [CrossRef] [PubMed]
- Racette, S.B.; Lin, X.; Ma, L.; Ostlund, R.E. Natural dietary phytosterols. J. AOAC Int. 2015, 98, 679–684. [Google Scholar] [CrossRef] [PubMed]
- De Smet, E.; Mensink, R.P.; Plat, J. Effects of plant sterols and stanols on intestinal cholesterol metabolism: Suggested mechanisms from past to present. Mol. Nutr. Food Res. 2012, 56, 1058–1072. [Google Scholar] [CrossRef]
- Ramprasath, V.R.; Awad, A.B. Role of phytosterols in cancer prevention and treatment. J. AOAC Int. 2015, 98, 735–738. [Google Scholar] [CrossRef]
- Shahzad, N.; Khan, W.; Shadab; Ali, A.; Saluja, S.S.; Sharma, S.; Al-Allaf, F.A.; Abduljaleel, Z.; Ibrahim, I.A.A.; Abdel-Wahab, A.F.; et al. Phytosterols as a natural anticancer agent: Current status and future perspective. Biomed. Pharmacother. 2017, 88, 786–794. [Google Scholar] [CrossRef]
- Sohn, S.I.; Rathinapriya, P.; Balaji, S.; Balan, D.J.; Swetha, T.K.; Durgadevi, R.; Alagulakshmi, S.; Singaraj, P.; Pandian, S. Phytosterols in seaweeds: An overview on biosynthesis to biomedical applications. Int. J. Mol. Sci. 2021, 22, 12691. [Google Scholar] [CrossRef]
- Kaur, R.; Myrie, S.B. Association of dietary phytosterols with cardiovascular disease biomarkers in humans. Lipids 2020, 55, 569–584. [Google Scholar] [CrossRef]
- Kopylov, A.T.; Malsagova, K.A.; Stepanov, A.A.; Kaysheva, A.L. Diversity of plant sterols metabolism: The impact on human health, sport, and accumulation of contaminating sterols. Nutrients 2021, 13, 1623. [Google Scholar] [CrossRef]
- Zhang, X.; Lin, K.; Li, Y. Highlights to phytosterols accumulation and equilibrium in plants: Biosynthetic pathway and feedback regulation. Plant Physiol. Biochem. 2020, 155, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Valitova, J.N.; Sulkarnayeva, A.G.; Minibayeva, F.V. Plant sterols: Diversity, biosynthesis, and physiological functions. Biochem. 2016, 81, 819–834. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.S.S.; Ali, K.; Dahuja, A.; Tyagi, A. Role of phytosterols in drought stress tolerance in rice. Plant Physiol. Biochem. 2015, 96, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, M.A. Plant sterols and the membrane environment. Trends Plant Sci. 1998, 3, 170–175. [Google Scholar] [CrossRef]
- Wedow, J.M.; Burroughs, C.H.; Rios Acosta, L.; Leakey, A.D.B.; Ainsworth, E.A. Age-dependent increase in α-tocopherol and phytosterols in maize leaves exposed to elevated ozone pollution. Plant Direct 2021, 5, e00307. [Google Scholar] [CrossRef] [PubMed]
- Clouse, S.D. Arabidopsis mutants reveal multiple roles for sterols in plant development. Plant Cell 2002, 14, 1995–2000. [Google Scholar] [CrossRef] [Green Version]
- Schrick, K.; Mayer, U.; Martin, G.; Bellini, C.; Kuhnt, C.; Schmidt, J.; Jürgens, G. Interactions between sterol biosynthesis genes in embryonic development of Arabidopsis. Plant J. 2002, 31, 61–73. [Google Scholar] [CrossRef]
- Yu, L.; Fan, J.; Zhou, C.; Xu, C. Sterols are required for the coordinated assembly of lipid droplets in developing seeds. Nat. Commun. 2021, 12, 5598. [Google Scholar] [CrossRef]
- Mondal, N.; Bhat, K.V.; Srivastava, P.S.; Sen, S.K. Effects of domestication bottleneck and selection on fatty acid desaturases in Indian sesame germplasm. Plant Genet. Resour. Characterisation Util. 2016, 14, 81–90. [Google Scholar] [CrossRef]
- Wei, X.; Liu, K.; Zhang, Y.; Feng, Q.; Wang, L.; Zhao, Y.; Li, D.; Zhao, Q.; Zhu, X.; Zhu, X.; et al. Genetic discovery for oil production and quality in sesame. Nat. Commun. 2015, 6, 8609. [Google Scholar] [CrossRef] [Green Version]
- Ke, T.; Dong, C.; Mao, H.; Zhao, Y.; Chen, H.; Liu, H.; Dong, X.; Tong, C.; Liu, S. Analysis of expression sequence tags from a full-length-enriched cDNA library of developing sesame seeds (Sesamum indicum). BMC Plant Biol. 2011, 11, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Q.; Xu, F.; Min, M.H.; Chu, S.H.; Kim, K.W.; Park, Y.J. Genome-wide association study of vitamin E using genotyping by sequencing in sesame (Sesamum indicum). Genes Genom. 2019, 41, 1085–1093. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Zhou, R.; Dossou, S.S.K.; Song, S.; Wang, L. Fine mapping of a major pleiotropic QTL associated with sesamin and sesamolin variation in sesame (Sesamum indicum L.). Plants 2021, 10, 1343. [Google Scholar] [CrossRef] [PubMed]
- Harker, M.; Hellyer, A.; Clayton, J.C.; Duvoix, A.; Lanot, A.; Safford, R. Co-ordinate regulation of sterol biosynthesis enzyme activity during accumulation of sterols in developing rape and tobacco seed. Planta 2003, 216, 707–715. [Google Scholar] [CrossRef]
- Zhao, X.; Shen, J.; Chang, K.J.; Kim, S.H. Analysis of fatty acids and phytosterols in ethanol extracts of Nelumbo nucifera seeds and rhizomes by GC-MS. J. Agric. Food Chem. 2013, 61, 6841–6847. [Google Scholar] [CrossRef]
- Liu, H.; Li, X.; Zhang, Q.; Yuan, P.; Liu, L.; King, G.J.; Ding, G.; Wang, S.; Cai, H.; Wang, C.; et al. Integrating a genome-wide association study with transcriptomic data to predict candidate genes and favourable haplotypes influencing Brassica napus seed phytate. DNA Res. 2021, 28, dsab011. [Google Scholar] [CrossRef]
- Dossa, K.; Li, D.; Zhou, R.; Yu, J.; Wang, L.; Zhang, Y.; You, J.; Liu, A.; Mmadi, M.A.; Fonceka, D.; et al. The genetic basis of drought tolerance in the high oil crop Sesamum indicum. Plant Biotechnol. J. 2019, 17, 1788–1803. [Google Scholar] [CrossRef] [Green Version]
- Turner, S. qqman: An R package for visualizing GWAS results using Q-Q and manhattan plots. J. Open Source Softw. 2018, 3, 731. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.S.; He, W.M.; Ji, J.J.; Zhang, C.; Guo, Y.; Yang, T.L. LDBlockShow: A fast and convenient tool for visualizing linkage disequilibrium and haplotype blocks based on variant call format files. Brief. Bioinform. 2021, 22, bbaa227. [Google Scholar] [CrossRef]
- Li, X.; Shi, Z.; Qie, Q.; Gao, J.; Wang, X.; Han, Y. CandiHap: A toolkit for haplotype analysis for sequence of samples and fast identification of candidate causal gene(s) in genome-wide association study. bioRxiv 2020. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, Y.; Li, D.; Dossa, K.; Wang, M.L.; Zhou, R.; Yu, J.; Zhang, X. Gene expression profiles that shape high and low oil content sesames. BMC Genet. 2019, 20, 45. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Dossou, S.S.K.; Wei, X.; Zhang, Y.; Li, D.; Yu, J.; Zhang, X. Transcriptome dynamics during black and white sesame (Sesamum indicum L.) seed development and identification of candidate genes associated with black pigmentation. Genes 2020, 11, 1399. [Google Scholar] [CrossRef] [PubMed]
- Mitsuda, N.; Iwase, A.; Yamamoto, H.; Yoshida, M.; Seki, M.; Shinozaki, K.; Ohme-Takagi, M. NAC transcription factors, NST1 and NST3, are key regulators of the formation of secondary walls in woody tissues of Arabidopsis. Plant Cell 2007, 19, 270–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, R.; Ye, Z.H. The arabidopsis NAC transcription factor NST2 functions together with SND1 and NST1 to regulate secondary wall biosynthesis in fibers of inflorescence stems. Plant Signal. Behav. 2015, 10, 37–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, R.; Richardson, E.A.; Ye, Z.H. Two NAC domain transcription factors, SND1 and NST1, function redundantly in regulation of secondary wall synthesis in fibers of Arabidopsis. Planta 2007, 225, 1603–1611. [Google Scholar] [CrossRef] [PubMed]
- Ryan, E.; Galvin, K.; O’Connor, T.P.; Maguire, A.R.; O’Brien, N.M. Phytosterol, squalene, tocopherol content and fatty acid profile of selected seeds, grains, and legumes. Plant Foods Hum. Nutr. 2007, 62, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Hu, H.; Yu, Y.; Zhao, J.; Liu, L.; Zhao, S.; Xie, J.; Li, C.; Shen, M. Simultaneous determination of tocopherols, phytosterols, and squalene in vegetable oils by high performance liquid chromatography-tandem mass spectrometry. Food Anal. Methods 2021, 14, 1567–1576. [Google Scholar] [CrossRef]
- Bouali, I.; Trabelsi, H.; Herchi, W.; Martine, L.; Albouchi, A.; Bouzaien, G.; Sifi, S.; Boukhchina, S.; Berdeaux, O. Analysis of pecan nut (Carya illinoinensis) unsaponifiable fraction. Effect of ripening stage on phytosterols and phytostanols composition. Food Chem. 2014, 164, 309–316. [Google Scholar] [CrossRef]
- Cegielska-Taras, T.; Nogala-Kałucka, M.; Szala, L.; Siger, A. Study of variation of tocochromanol and phytosterol contents in black and yellow seeds of Brassica napus L. doubled haploid populations. Acta Sci. Pol. Technol. Aliment. 2016, 15, 321–332. [Google Scholar] [CrossRef] [Green Version]
- Haddadi, P.; Ebrahimi, A.; Langlade, N.B.; Yazdi-samadi, B.; Berger, M.; Calmon, A.; Naghavi, M.R.; Vincourt, P.; Sarrafi, A. Genetic dissection of tocopherol and phytosterol in recombinant inbred lines of sunflower through quantitative trait locus analysis and the candidate gene approach. Mol. Breed. 2012, 29, 717–729. [Google Scholar] [CrossRef]
- Nurmi, T.; Nyström, L.; Edelmann, M.; Lampi, A.M.; Piironen, V. Phytosterols in wheat genotypes in the health grain diversity screen. J. Agric. Food Chem. 2008, 56, 9710–9715. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.S.S.; Mawlong, I.; Ali, K.; Tyagi, A. Regulation of phytosterol biosynthetic pathway during drought stress in rice. Plant Physiol. Biochem. 2018, 129, 11–20. [Google Scholar] [CrossRef]
- Schaeffer, A.; Bronner, R.; Benveniste, P.; Schaller, H. The ratio of campesterol to sitosterol that modulates growth in Arabidopsis is controlled by STEROL METHYLTRANSFERASE 2;1. Plant J. 2001, 25, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Zauber, H.; Burgos, A.; Garapati, P.; Schulze, W.X. Plasma membrane lipid-protein interactions affect signaling processes in sterol-biosynthesis mutants in Arabidopsis thaliana. Front. Plant Sci. 2014, 5, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berhe, M.; Dossa, K.; You, J.; Mboup, P.A.; Diallo, I.N.; Diouf, D.; Zhang, X.; Wang, L. Genome-wide association study and its applications in the non-model crop Sesamum indicum. BMC Plant Biol. 2021, 21, 283. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Liu, Y.; Liu, Y.; Cui, X.; Sun, Z.; Du, Z.; Wu, K.; Jiang, X.; Mei, H.; Zheng, Y. Genome-wide association study of seed coat color in sesame (Sesamum indicum L.). PLoS ONE 2021, 16, e0251526. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Yuan, Y.; Spiekerman, J.J.; Guley, J.T.; Egbosiuba, J.C.; Ye, Z.H. Functional characterization of NAC and MYB transcription factors involved in regulation of biomass production in switchgrass (Panicum virgatum). PLoS ONE 2015, 10, e0251526. [Google Scholar] [CrossRef]
- Niu, Q.; Tan, K.; Zang, Z.; Xiao, Z.; Chen, K.; Hu, M.; Luo, M. Modification of phytosterol composition influences cotton fiber cell elongation and secondary cell wall deposition. BMC Plant Biol. 2019, 19, 208. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Environment | Min (mg/g) | Max (mg/g) | Mean (mg/g) | SD | CV (%) |
|---|---|---|---|---|---|---|
| Campesterol | 2018_WH | 0 | 2.847 | 1.129 | 0.352 | 31.14 |
| 2019_ZMD | 0 | 2.751 | 1.298 | 0.375 | 28.85 | |
| Stigmasterol | 2018_WH | 0.415 | 1.826 | 0.867 | 0.193 | 22.24 |
| 2019_ZMD | 0.356 | 1.682 | 0.793 | 0.193 | 24.38 | |
| β-Sitosterol | 2018_WH | 1.794 | 4.656 | 3.003 | 0.477 | 15.87 |
| 2019_ZMD | 1.6 | 4.114 | 2.662 | 0.468 | 17.58 | |
| Δ5-Avenasterol | 2018_WH | 0 | 1.307 | 0.467 | 0.181 | 38.72 |
| 2019_ZMD | 0.155 | 1.012 | 0.544 | 0.153 | 28.04 | |
| Total phytosterol | 2018_WH | 2.694 | 8.206 | 5.496 | 0.798 | 14.6 |
| 2019_ZMD | 2.986 | 8.388 | 5.297 | 0.853 | 16.11 |
| Environment | Trait | Locus | Chr | Position | Ref | Alt | p-Value | PVE (%) |
|---|---|---|---|---|---|---|---|---|
| WH | Campesterol | Chr2-370158 | 2 | 370158 | C | T | 5.603 | 8.94 |
| Chr3-18989758 | 3 | 18989758 | C | G | 5.744 | 6.47 | ||
| Chr4-10138424 | 4 | 10138424 | C | T | 6.095 | 9.60 | ||
| Chr5-9706516 | 5 | 9706516 | C | T | 5.312 | 7.76 | ||
| Chr6-39270 | 6 | 39270 | A | C | 5.424 | 1.63 | ||
| Chr8-5486783 | 8 | 5486783 | C | A | 6.005 | 2.86 | ||
| Chr11-142842 | 11 | 142842 | C | A | 9.079 | 7.95 | ||
| Chr13-12554381 | 13 | 12554381 | G | A | 6.257 | 9.03 | ||
| Stigmasterol | Chr-1-13179445 | 1 | 13179445 | G | A | 5.207 | 4.53 | |
| Chr3-15930743 | 3 | 15930743 | G | A | 6.098 | 5.22 | ||
| Chr4-18569755 | 4 | 18569755 | G | A | 6.668 | 11.40 | ||
| Chr8-25004528 | 8 | 25004528 | C | A | 5.954 | 3.03 | ||
| Chr10-15465843 | 10 | 15465843 | T | C | 5.282 | 6.67 | ||
| Chr11-142842 | 11 | 142842 | C | A | 19.675 | 28.24 | ||
| Chr12-10292195 | 12 | 10292195 | C | T | 7.182 | 9.44 | ||
| β-Sitosterol | Chr4-4016241 | 4 | 4016241 | T | C | 5.033 | 5.98 | |
| Chr8-3623152 | 8 | 3623152 | G | A | 5.076 | 3.33 | ||
| Chr11-1227222 | 11 | 1227222 | T | C | 5.321 | 8.35 | ||
| Chr13-14801278 | 13 | 14801278 | A | G | 5.124 | 4.80 | ||
| Δ5-Avenasterol | Chr2-7188241 | 2 | 7188241 | C | A | 5.607 | 2.61 | |
| Chr11-5738482 | 11 | 5738482 | G | A | 10.211 | 6.42 | ||
| Chr12-10290279 | 12 | 10290279 | G | A | 5.115 | 7.19 | ||
| ZMD | Campesterol | Chr4-13279017 | 4 | 13279017 | C | T | 5.182 | 1.37 |
| Chr6-39270 | 6 | 39270 | A | C | 7.070 | 8.20 | ||
| Chr11-142842 | 11 | 142842 | C | A | 14.887 | 17.87 | ||
| Stigmasterol | Chr3-23741 | 3 | 23741 | G | T | 12.830 | 22.01 | |
| Chr6-39270 | 6 | 39270 | A | C | 12.154 | 19.11 | ||
| Chr11-142842 | 11 | 142842 | C | A | 18.351 | 22.11 | ||
| Chr7-17668358 | 7 | 17668358 | G | A | 10.523 | 19.41 | ||
| Chr11-111895 | 11 | 111895 | G | A | 21.507 | 31.02 | ||
| Chr13-19899439 | 13 | 19899439 | A | G | 10.635 | 19.54 | ||
| β-Sitosterol | Chr3-14800933 | 3 | 14800933 | G | A | 5.301 | 4.41 | |
| Δ5-Avenasterol | Chr9-6530662 | 9 | 6530662 | C | T | 5.238 | 6.75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Zhou, Q.; Dossou, S.S.K.; Zhou, R.; Zhao, Y.; Zhou, W.; Zhang, Y.; Li, D.; You, J.; Wang, L. Genome-Wide Association Study Uncovers Loci and Candidate Genes Underlying Phytosterol Variation in Sesame (Sesamum indicum L.). Agriculture 2022, 12, 392. https://doi.org/10.3390/agriculture12030392
Wang Z, Zhou Q, Dossou SSK, Zhou R, Zhao Y, Zhou W, Zhang Y, Li D, You J, Wang L. Genome-Wide Association Study Uncovers Loci and Candidate Genes Underlying Phytosterol Variation in Sesame (Sesamum indicum L.). Agriculture. 2022; 12(3):392. https://doi.org/10.3390/agriculture12030392
Chicago/Turabian StyleWang, Zhijian, Qi Zhou, Senouwa Segla Koffi Dossou, Rong Zhou, Yingzhong Zhao, Wangyi Zhou, Yanxin Zhang, Donghua Li, Jun You, and Linhai Wang. 2022. "Genome-Wide Association Study Uncovers Loci and Candidate Genes Underlying Phytosterol Variation in Sesame (Sesamum indicum L.)" Agriculture 12, no. 3: 392. https://doi.org/10.3390/agriculture12030392
APA StyleWang, Z., Zhou, Q., Dossou, S. S. K., Zhou, R., Zhao, Y., Zhou, W., Zhang, Y., Li, D., You, J., & Wang, L. (2022). Genome-Wide Association Study Uncovers Loci and Candidate Genes Underlying Phytosterol Variation in Sesame (Sesamum indicum L.). Agriculture, 12(3), 392. https://doi.org/10.3390/agriculture12030392