
Assessment of the Immune Response of Clinically Infected Calves to Cryptosporidium parvum Infection

 ,
, 
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Calf Enrollment
2.2. Fecal Sample Examination
2.3. Molecular Confirmation
2.4. Acute-Phase Proteins (APPs)
2.5. Proinflammatory Cytokines (CYTs)
2.6. Procalcitonin and Neopterin
2.7. Oxidative Stress Biomarkers
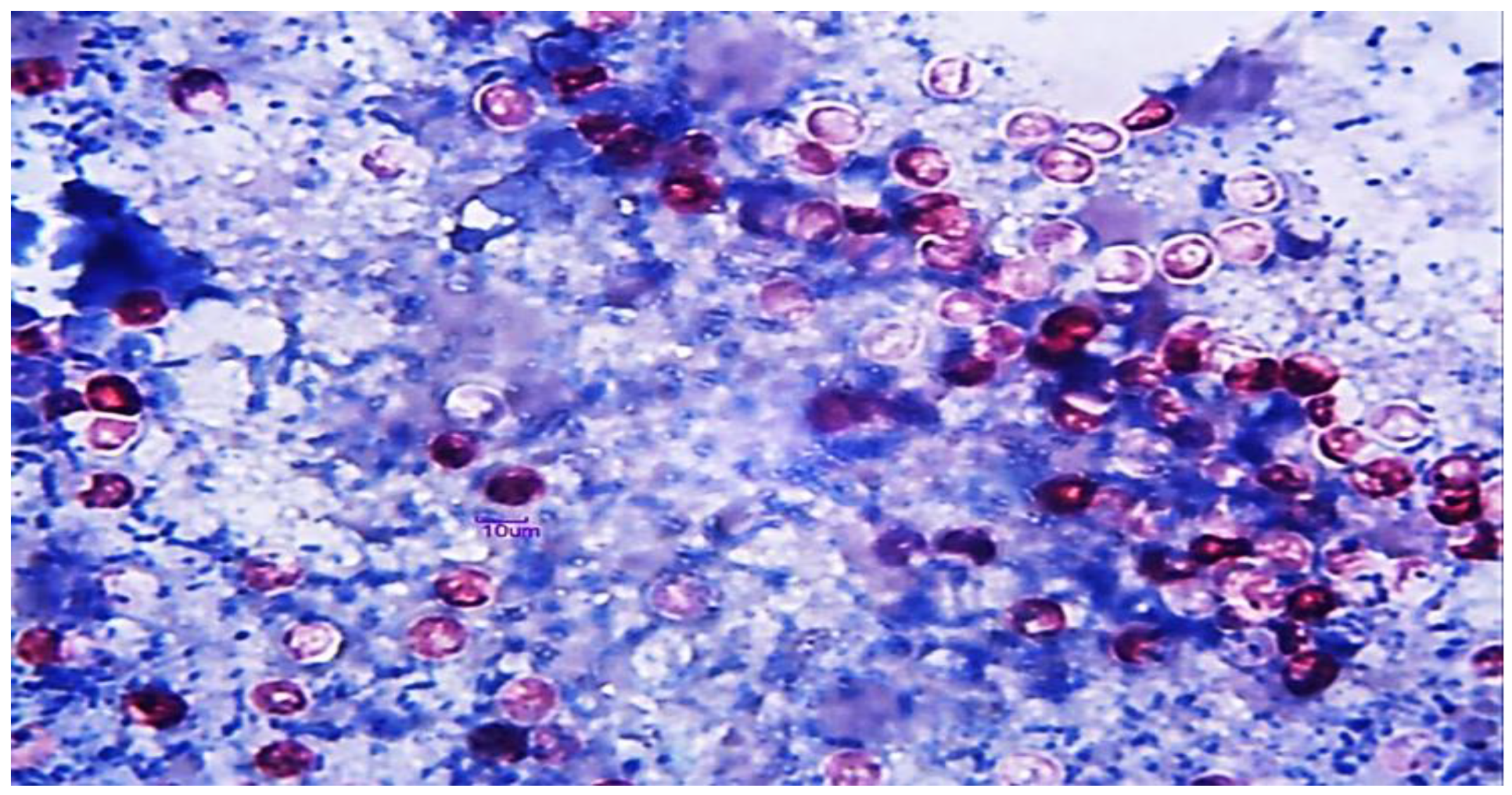
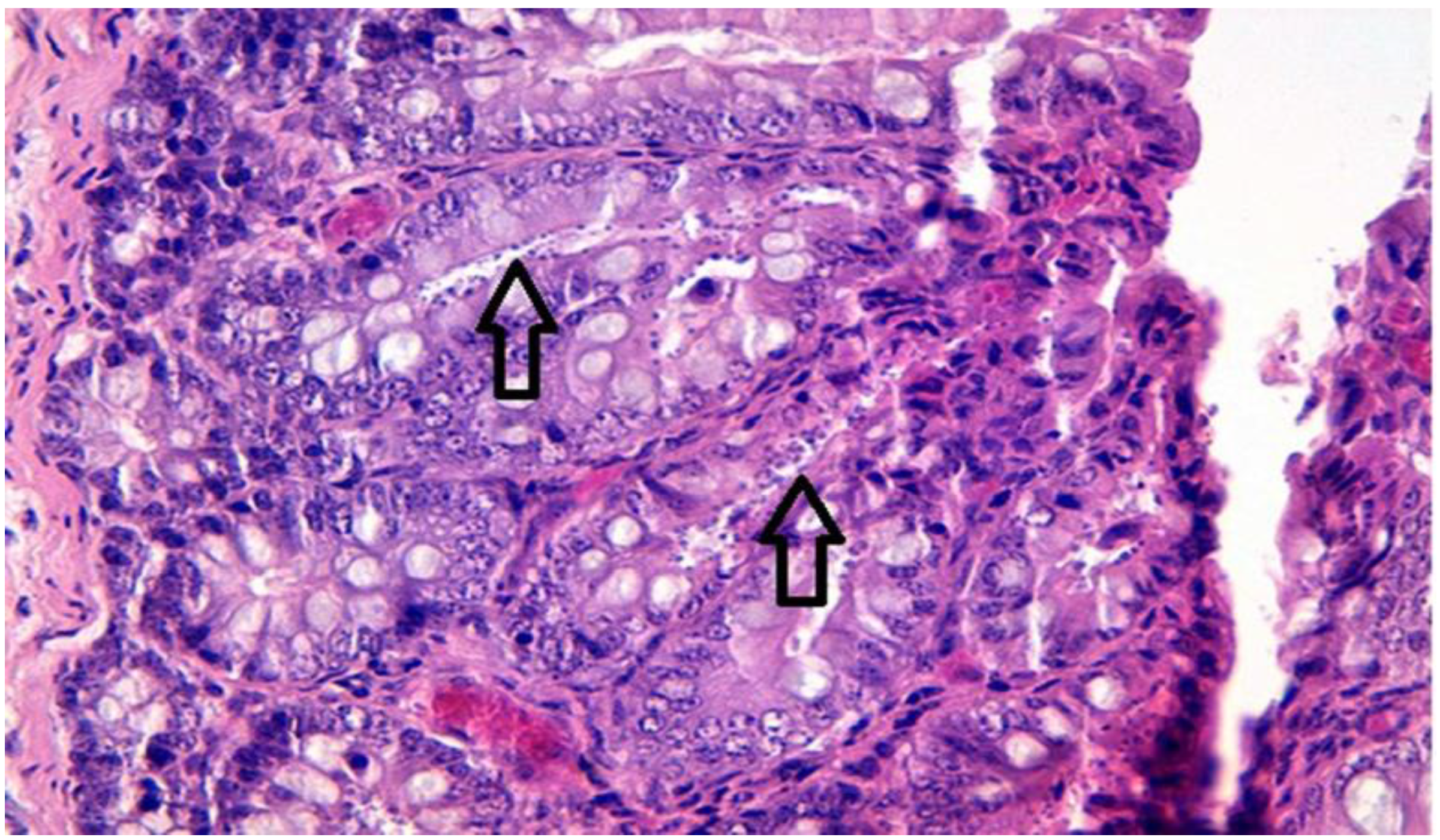
2.8. Histopathology
2.9. Data Analysis
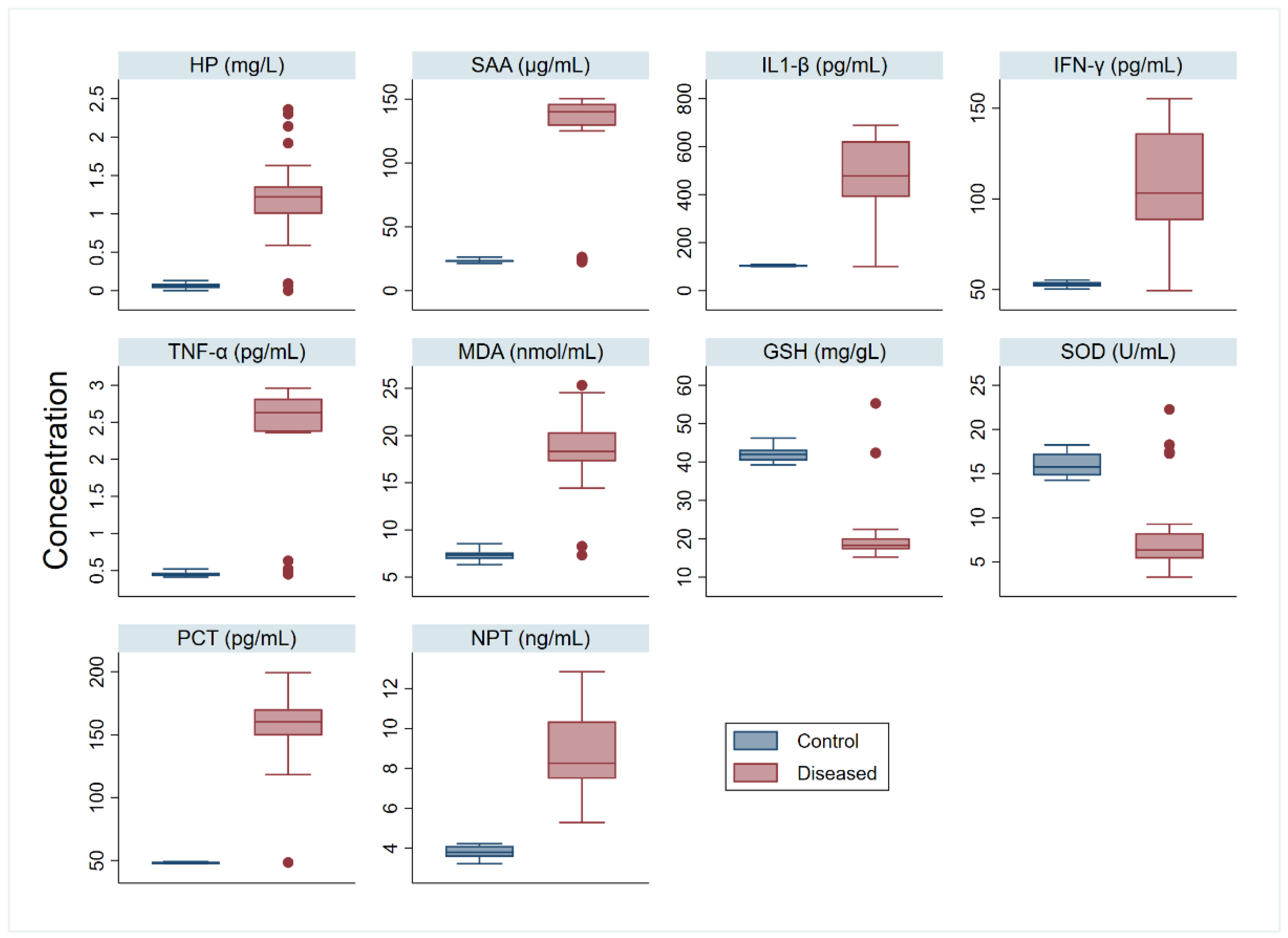
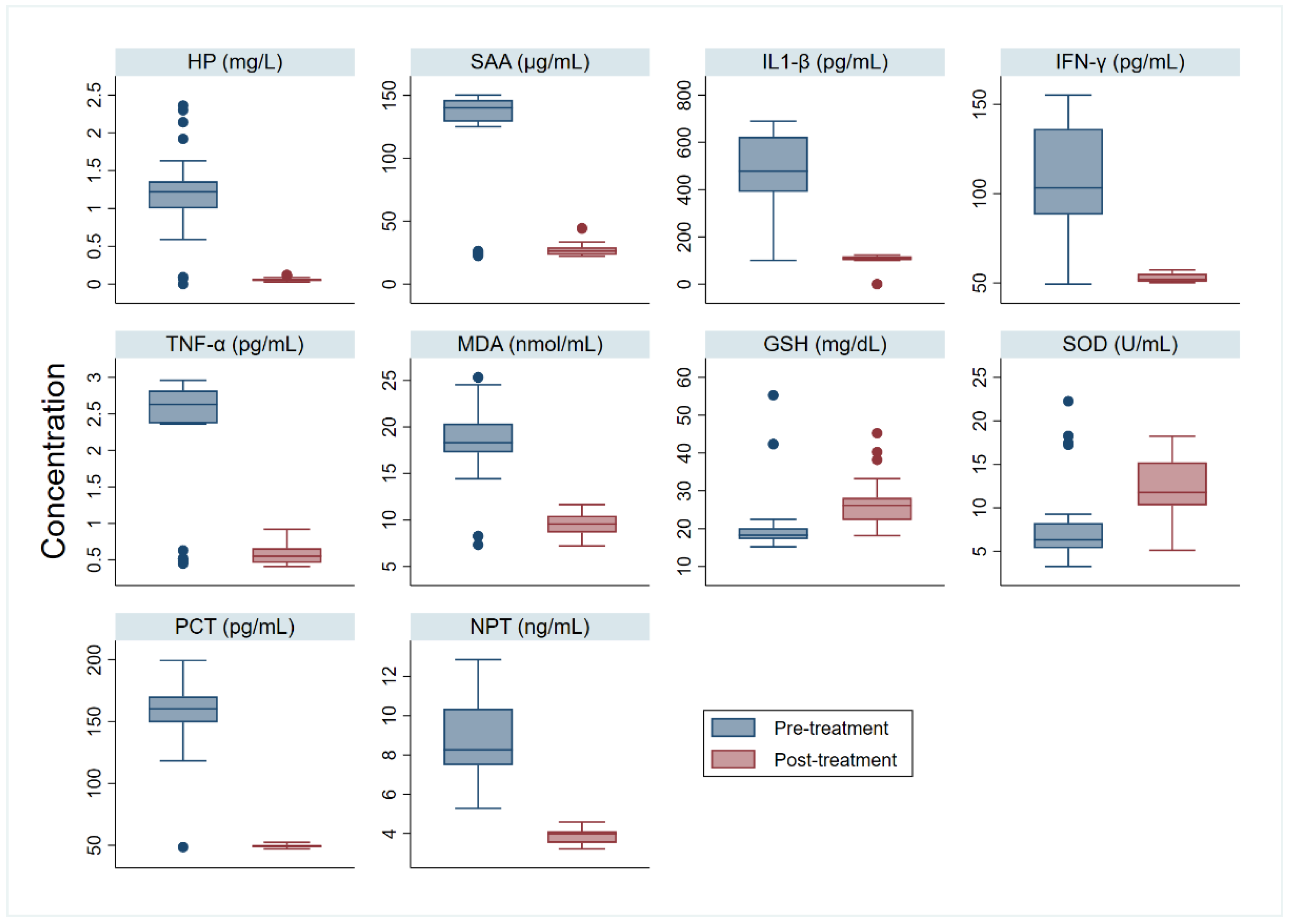
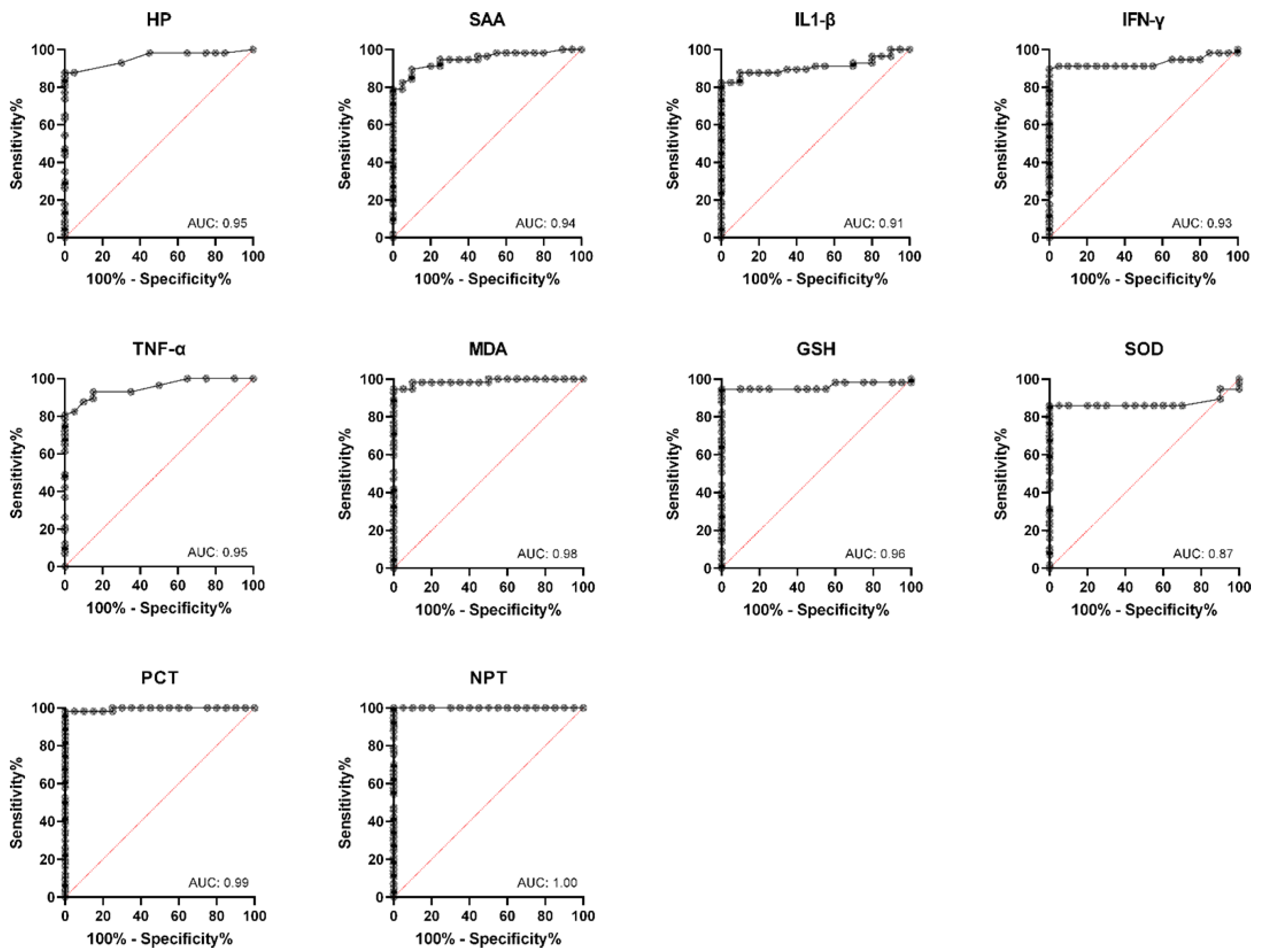
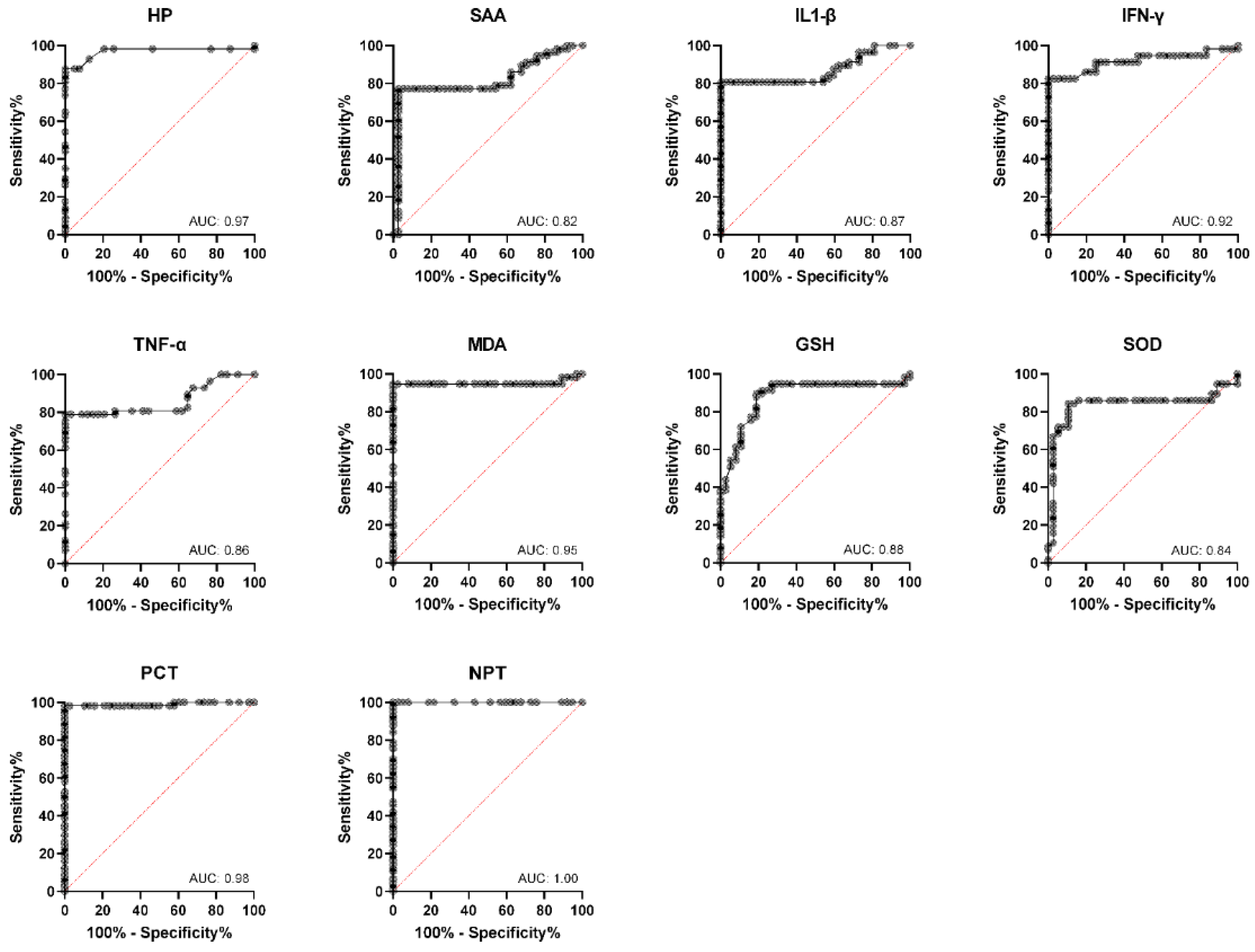
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De la Fuente, R.; Luzon, M.; Ruiz-Santa-Quiteria, J.; Garcıa, A.; Cid, D.; Orden, J.; Garcıa, S.; Sanz, R.; Gomez-Bautista, M. Cryptosporidium and concurrent infections with other major enterophatogens in 1 to 30-day-old diarrheic dairy calves in central Spain. Vet. Parasitol. 1999, 80, 179–185. [Google Scholar] [CrossRef]
- Wyatt, C.R.; Riggs, M.W.; Fayer, R. Cryptosporidiosis in neonatal calves. Vet. Clin. Food Anim. Pract. 2010, 26, 89–103. [Google Scholar] [CrossRef]
- Dixon, B.; Parrington, L.; Cook, A.; Pintar, K.; Pollari, F.; Kelton, D.; Farber, J. The potential for zoonotic transmission of Giardia duodenalis and Cryptosporidium spp. from beef and dairy cattle in Ontario, Canada. Vet. Parasitol. 2011, 175, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, P.; Ajjampur, S.S.R.; Chidambaram, D.; Kattula, D.; Rajan, D.P.; Ward, H.; Kang, G. Investigation of potential zoonotic transmission of cryptosporidiosis in southern India. Am. J. Trop. Med. Hyg. 2011, 85, 657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirvonen, J.; Huszenicza, G.; Kulcsar, M.; Pyörälä, S. Acute-phase response in dairy cows with acute postpartum metritis. Theriogenology 1999, 51, 1071–1083. [Google Scholar] [CrossRef]
- El-Deeb, W.M. Clinicobiochemical investigations of gangrenous mastitis in does: Immunological responses and oxidative stress biomarkers. J. Zhejiang Univ. Sci. B 2013, 14, 33–39. [Google Scholar] [CrossRef] [Green Version]
- El-Deeb, W.; Elmoslemany, A. Cardiac and oxidative stress biomarkers in Trypanosoma evansi infected camels: Diagnostic and prognostic prominence. Parasitology 2015, 142, 767–772. [Google Scholar] [CrossRef]
- El-Bahr, S.; El-Deeb, W. Acute-phase proteins, oxidative stress biomarkers, proinflammatory cytokines, and cardiac troponin in Arabian mares affected with pyometra. Theriogenology 2016, 86, 1132–1136. [Google Scholar] [CrossRef]
- El-Deeb, W.M.; Elmoslemany, A.M. The diagnostic accuracy of acute phase proteins and proinflammatory cytokines in sheep with pneumonic pasteurellosis. PeerJ 2016, 4, e2161. [Google Scholar] [CrossRef] [Green Version]
- El-Deeb, W.M.; Elmoslemany, A.M. Acute phase proteins as biomarkers of Urinary Tract Infection in dairy cows: Diagnostic and prognostic accuracy. Jpn. J. Vet. Res. 2016, 64, 57–66. [Google Scholar]
- El-Deeb, W.; Ghoneim, I.; Fayez, M.; Elsohaby, I.; Alhaider, A.; ElGioushy, M. Acute phase proteins, proinflammatory cytokines and oxidative stress biomarkers in sheep, goats and she-camels with Coxiella burnetii infection-induced abortion. Comp. Immunol. Microbiol. Infect. Dis. 2019, 67, 101352. [Google Scholar] [CrossRef] [PubMed]
- Reinhart, K.; Karzai, W.; Meisner, M. Procalcitonin as a marker of the systemic inflammatory response to infection. Intensive Care Med. 2000, 26, 1193. [Google Scholar] [CrossRef]
- Werner-Felmayer, G.; Werner, E.R.; Fuchs, D.; Hausen, A.; Reibnegger, G.; Wachter, H. Neopterin formation and tryptophan degradation by a human myelomonocytic cell line (THP-1) upon cytokine treatment. Cancer Res. 1990, 50, 2863–2867. [Google Scholar] [PubMed]
- Grinberg, A.; Markovics, A.; Galindez, J.; Lopez-Villalobos, N.; Kosak, A.; Tranquillo, V. Controlling the onset of natural cryptosporidiosis in calves with paromomycin sulphate. Vet. Rec. 2002, 151, 606–608. [Google Scholar] [CrossRef] [PubMed]
- Elitok, B.; Elitok, Ö.M.; Pulat, H. Efficacy of azithromycin dihydrate in treatment of cryptosporidiosis in naturally infected dairy calves. J. Vet. Intern. Med. 2005, 19, 590–593. [Google Scholar] [CrossRef]
- Lallemond, M.; Villeneuve, A.; Belda, J.; Dubreuil, P. Field study of the efficacy of halofuginone and decoquinate in the treatment of cryptosporidiosis in veal calves. Vet. Rec. 2006, 159, 672–676. [Google Scholar] [CrossRef]
- Van Duijkeren, E.; Vulto, A.; van Oldruitenborgh-Oosterbaan, M.S.; Kessels, B.G.; Van Miert, A.S.; Breukink, H.J. Pharmacokinetics of trimethoprim/sulphachlorpyridazine in horses after oral, nasogastric and intravenous administration. J. Vet. Pharm. 1995, 18, 47–53. [Google Scholar] [CrossRef]
- Henriksen, S.A.; Pohlenz, J.F.L. Staining of cryptosporidia by a modified Ziehl-Neelsen technique. Acta Vet. Scand. 1981, 22, 594. [Google Scholar] [CrossRef] [PubMed]
- Fayer, R. General biology. Cryptosporidium Cryptosporidiosis 2008, 2, 1–35. [Google Scholar]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Beutler, E. Improved method for the determination of blood glutathione. J. Lab. Clin. Med. 1963, 61, 882–888. [Google Scholar] [PubMed]
- Nishikimi, M.; Rao, N.A.; Yagi, K. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun. 1972, 46, 849–854. [Google Scholar] [CrossRef]
- Suvarna, S.; Layton, C.; Bancroft, J. The hematoxylins and eosin. In Bancroft’s Theory and Practice of Histological Techniques, 7th ed.; Churchill Livingstone: London, UK, 2013; pp. 172–186. [Google Scholar]
- Urieli-Shoval, S.; Linke, R.P.; Matzner, Y. Expression and function of serum amyloid A, a major acute-phase protein, in normal and disease states. Curr. Opin. Hematol. 2000, 7, 64–69. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Sang, H.; Ye, R.D. Serum amyloid A induces IL-8 secretion through a G protein–coupled receptor, FPRL1/LXA4R. Blood J. Am. Soc. Hematol. 2003, 101, 1572–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murata, H.; Shimada, N.; Yoshioka, M. Current research on acute phase proteins in veterinary diagnosis: An overview. Vet. J. 2004, 168, 28–40. [Google Scholar] [CrossRef]
- Orro, T.; Pohjanvirta, T.; Rikula, U.; Huovilainen, A.; Alasuutari, S.; Sihvonen, L.; Pelkonen, S.; Soveri, T. Acute phase protein changes in calves during an outbreak of respiratory disease caused by bovine respiratory syncytial virus. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Pourjafar, M.; Badiei, K.; Nazifi, S.; Naghib, S. Acute phase response in Holstein dairy calves affected with diarrhoea. Bulg. J. Vet. Med 2011, 14, 142–149. [Google Scholar]
- Hajimohammadi, A.; Nazifi, S.; Ansari-Lari, M.; Khoshmanzar, M.R.; Bigdeli, S.M. Identifying relationships among acute phase proteins (haptoglobin, serum amyloid A, fibrinogen, ceruloplasmin) and clinical findings in dairy calf diarrhea. Comp. Clin. Pathol. 2013, 22, 227–232. [Google Scholar] [CrossRef]
- Dinler, C.; Ulutas, B.; Voyvoda, H.; Ulutas, P.A.; Ural, K.; Karagenc, T. Haptoglobin and serum amyloid-A concentrations and their relationship with oocyst count in neonatal lambs experimentally infected with Cryptosporidium parvum. Vet. Parasitol. 2017, 247, 49–56. [Google Scholar] [CrossRef]
- Hashemnia, M.; Khodakaram-Tafti, A.; Razavi, S.M.; Nazifi, S. Changing patterns of acute phase proteins and inflammatory mediators in experimental caprine coccidiosis. Korean J. Parasitol. 2011, 49, 213. [Google Scholar] [CrossRef]
- Lassen, B.; Bangoura, B.; Lepik, T.; Orro, T. Systemic acute phase proteins response in calves experimentally infected with Eimeria zuernii. Vet. Parasitol. 2015, 212, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Cenesiz, M.; Sağkan Öztürk, A.; Dalğın, D.; Yarım, G.; Çiftçi, G.; Özdemir, R.; Güzel, M.; Kazak, F.; Cenesiz, S. Investigation of acute phase reactants and antioxidant capacity in calves infected with Cryptosporidium parvum. Kafkas Univ. Vet. Fak. Derg. 2017, 23, 481–485. [Google Scholar]
- Mead, J.R.; You, X. Susceptibility differences to Cryptosporidium parvum infection in two strains of gamma interferon knockout mice. J. Parasitol. 1998, 84, 1045–1048. [Google Scholar] [CrossRef] [PubMed]
- Lacroix-Lamandé, S.; Mancassola, R.; Naciri, M.; Laurent, F. Role of gamma interferon in chemokine expression in the ileum of mice and in a murine intestinal epithelial cell line after Cryptosporidium parvum infection. Infect. Immun. 2002, 70, 2090–2099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedi, B.; Mead, J. Cryptosporidium parvum antigens induce mouse and human dendritic cells to generate Th1-enhancing cytokines. Parasite Immunol. 2012, 34, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Perez-Cordon, G.; Yang, G.; Zhou, B.; Nie, W.; Li, S.; Shi, L.; Tzipori, S.; Feng, H. Interaction of Cryptosporidium parvum with mouse dendritic cells leads to their activation and parasite transportation to mesenteric lymph nodes. Pathog. Dis. 2014, 70, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ovington, K.; Alleva, L.; Kerr, E. Cytokines and immunological control of Eimeria spp. Int. J. Parasitol. 1995, 25, 1331–1351. [Google Scholar] [CrossRef]
- Laurent, F.; Kagnoff, M.F.; Savidge, T.C.; Naciri, M.; Eckmann, L. Human Intestinal Epithelial Cells Respond to Cryptosporidium parvum Infection with Increased Prostaglandin H Synthase 2 Expression and Prostaglandin E2and F2α Production. Infect. Immun. 1998, 66, 1787–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCOOL, D.J.; Marcon, M.A.; Forstner, J.F.; Forstner, G.G. The T84 human colonic adenocarcinoma cell line produces mucin in culture and releases it in response to various secretagogues. Biochem. J. 1990, 267, 491–500. [Google Scholar] [CrossRef]
- Thea, D.M.; Pereira, M.E.; Kotler, D.; Sterling, C.R.; Keusch, G.T. Identification and partial purification of a lectin on the surface of the sporozoite of Cryptosporidium parvum. J. Parasitol. 1992, 78, 886–893. [Google Scholar] [CrossRef] [PubMed]
- Robinson, P.; Okhuysen, P.C.; Chappell, C.L.; Lewis, D.E.; Shahab, I.; Janecki, A.; White, A.C., Jr. Expression of tumor necrosis factor alpha and interleukin 1β in jejuna of volunteers after experimental challenge with Cryptosporidium parvum correlates with exposure but not with symptoms. Infect. Immun. 2001, 69, 1172–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assicot, M.; Bohuon, C.; Gendrel, D.; Raymond, J.; Carsin, H.; Guilbaud, J. High serum procalcitonin concentrations in patients with sepsis and infection. Lancet 1993, 341, 515–518. [Google Scholar] [CrossRef]
- Dandona, P.; Nix, D.; Wilson, M.F.; Aljada, A.; Love, J.; Assicot, M.; Bohuon, C. Procalcitonin increase after endotoxin injection in normal subjects. J. Clin. Endocrinol. Metab. 1994, 79, 1605–1608. [Google Scholar] [PubMed]
- Facer, C.A. Malaria antigens stimulate neopterin secretion by PBMC and U937 cells. Microbiol. Immunol. 1995, 39, 207–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abd El-Azeem, A.; Hamdy, G.; Saraya, M.; Fawzy, E.; Anwar, E.; Abdulattif, S. The role of procalcitonin as a guide for the diagnosis, prognosis, and decision of antibiotic therapy for lower respiratory tract infections. Egypt. J. Chest Dis. Tuberc. 2013, 62, 687–695. [Google Scholar] [CrossRef] [Green Version]
- Nanda, S.K.; Dinakaran, A.; Bhat, K.; Kanungo, R. Diagnostic and prognostic role of Procalcitonin in sepsis in a tertiary care hospital. Biomed. Res. (0970-938X) 2016, 27, 79–83. [Google Scholar]
- El-Deeb, W.M.; Elmoslemany, A.M.; Salem, M.A. Cardiac troponin I and immune-inflammatory response in horses with strangles. J. Equine Vet. Sci. 2017, 51, 18–23. [Google Scholar] [CrossRef]
- Rodríguez de Sotillo, D.; Velly, A.; Hadley, M.; Fricton, J. Evidence of oxidative stress in temporomandibular disorders: A pilot study. J. Oral Rehabil. 2011, 38, 722–728. [Google Scholar] [CrossRef]
- El-Deeb, W.M.; Buczinski, S. The diagnostic and prognostic importance of oxidative stress biomarkers and acute phase proteins in urinary tract infection (UTI) in camels. PeerJ 2015, 3, e1363. [Google Scholar] [CrossRef] [Green Version]
- Celi, P. Biomarkers of oxidative stress in ruminant medicine. Immunopharmacol. Immunotoxicol. 2011, 33, 233–240. [Google Scholar] [CrossRef]
- Castillo, C.; Hernandez, J.; Bravo, A.; Lopez-Alonso, M.; Pereira, V.; Benedito, J. Oxidative status during late pregnancy and early lactation in dairy cows. Vet. J. 2005, 169, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Ayala, A.; Muñoz, M.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxidative Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- El-Deeb, W.; Fouda, T.; El-Bahr, S. Clinico-biochemical Investigation of Paratuberculosis of Dromedary Camels in Saudi Arabia: Proinflammatory Cytokines, Acute Phase Proteins and Oxidative Stress Biomarkers. Pak. Vet. J. 2014, 34, 484–488. [Google Scholar]
- Tünger, Ö. Sepsisin tanı ve izlenimde prokalsitonin, CRP ve diğer göstergeler. Klimik Derg. 2007, 20, 1. [Google Scholar]
- El-Deeb, W.M.; Younis, E.E. Clinical and biochemical studies on Theileria annulata in Egyptian buffaloes (Bubalus bubalis) with particular orientation to oxidative stress and ketosis relationship. Vet. Parasitol. 2009, 164, 301–305. [Google Scholar] [CrossRef]
- El-Deeb, W.M.; Iacob, O.C. Serum acute phase proteins in control and Theileria annulata infected water buffaloes (Bubalus bubalis). Vet. Parasitol. 2012, 190, 12–18. [Google Scholar] [CrossRef] [PubMed]
- El-Deeb, W.M.; Tharwat, M. Lipoproteins profile, acute phase proteins, proinflammatory cytokines and oxidative stress biomarkers in sheep with pneumonic pasteurellosis. Comp. Clin. Pathol. 2015, 24, 581–588. [Google Scholar] [CrossRef]
- Gupta, R.K.; Patel, A.K.; Shah, N.; Choudhary, A.K.; Jha, U.K.; Yadav, U.C.; Gupta, P.K.; Pakuwal, U. Oxidative stress and antioxidants in disease and cancer: A review. Asian Pac. J. Cancer Prev. 2014, 15, 4405–4409. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B. Antioxidants in human health and disease. Annu. Rev. Nutr. 1996, 16, 33–50. [Google Scholar] [CrossRef]
- Sies, H. Oxidative stress: Oxidants and antioxidants. Exp. Physiol. Transl. Integr. 1997, 82, 291–295. [Google Scholar] [CrossRef]
- Quílez, J.; Vergara-Castiblanco, C.; Monteagudo, L.; del Cacho, E.; Sánchez-Acedo, C. Host association of Cryptosporidium parvum populations infecting domestic ruminants in Spain. Appl. Environ. Microbiol. 2013, 79, 5363–5371. [Google Scholar] [CrossRef] [Green Version]
- Metwaly, M.S.; Dkhil, M.A.; Al-Quraishy, S.; Al Omar, S.Y. Protective effects of palm pollen aqueous extract against Eimeria papillata induced intestinal damage in mice. Pak. J. Zool. 2015, 47, 971–979. [Google Scholar]
- Abd El-Aziz, T.H.; El-Beih, N.; Soufy, H.; Nasr, S.M.; Khalil, F.A.; Sharaf, M. Effect of Egyptian propolis on lipid profile and oxidative status in comparison with nitazoxanide in immunosuppressed rats infected with Cryptosporidium spp. Glob. Vet. 2014, 13, 17–27. [Google Scholar]
- Wang, C.; Wu, Y.; Qin, J.; Sun, H.; He, H. Induced susceptibility of host is associated with an impaired antioxidant system following infection with Cryptosporidium parvum in Se-deficient mice. PLoS ONE 2009, 4, e4628. [Google Scholar] [CrossRef] [PubMed]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Hamerlinck, F.F.V. Neopterin: A review. Exp. Dermatol. 1999, 8, 167–176. [Google Scholar] [CrossRef]
- Skinner, J.; Brown, R.; Roberts, L. Bovine haptoglobin response in clinically defined field conditions. Vet. Rec. 1991, 128, 147–149. [Google Scholar] [CrossRef] [PubMed]
- Heegaard, P.M.; Godson, D.L.; Toussaint, M.J.; Tjørnehøj, K.; Larsen, L.E.; Viuff, B.; Rønsholt, L. The acute phase response of haptoglobin and serum amyloid A (SAA) in cattle undergoing experimental infection with bovine respiratory syncytial virus. Vet. Immunol. Immunopathol. 2000, 77, 151–159. [Google Scholar] [CrossRef]
- Carter, J.N.; Meredith, G.L.; Montelongo, M.; Gill, D.R.; Krehbiel, C.R.; Payton, M.E.; Confer, A.W. Relationship of vitamin E supplementation and antimicrobial treatment with acute-phase protein responses in cattle affected by naturally acquired respiratory tract disease. Am. J. Vet. Res. 2002, 63, 1111–1117. [Google Scholar] [CrossRef] [PubMed]
- Giamarellos-Bourboulis, E.J.; Mega, A.; Grecka, P.; Scarpa, N.; Koratzanis, G.; Thomopoulos, G.; Giamarellou, H. Procalcitonin: A marker to clearly differentiate systemic inflammatory response syndrome and sepsis in the critically ill patient? Intensive Care Med. 2002, 28, 1351–1356. [Google Scholar] [CrossRef]
- Baydar, T.; Yuksel, O.; Sahin, T.T.; Dikmen, K.; Girgin, G.; Sipahi, H.; Kurukahvecioglu, O.; Bostanci, H.; Sare, M. Neopterin as a prognostic biomarker in intensive care unit patients. J. Crit. Care 2009, 24, 318–321. [Google Scholar] [CrossRef] [PubMed]
- LIUD, S. Prognostic value of procalcitoninin adult patients with sepsis: Asystematic review and meta-analysis. PLoS ONE 2015, 10, e0129450. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Healthy/ Diseased | HP | SAA | IL-1β | IFN-γ | TNF-α | MDA | GSH | SOD | PCT | NPT |
|---|---|---|---|---|---|---|---|---|---|---|---|
| HP | 0.69 | 1.00 | |||||||||
| SAA | 0.68 | 0.45 | 1.00 | ||||||||
| IL-1β | 0.62 | 0.64 | 0.54 | 1.00 | |||||||
| IFN-γ | 0.66 | 0.61 | 0.64 | 0.72 | 1.00 | ||||||
| TNF-α | 0.70 | 0.66 | 0.35 | 0.52 | 0.54 | 1.00 | |||||
| MDA | 0.74 | 0.64 | 0.63 | 0.79 | 0.79 | 0.56 | 1.00 | ||||
| GSH | −0.70 | −0.46 | −0.51 | −0.52 | −0.49 | −0.57 | −0.59 | 1.00 | |||
| SOD | −0.57 | −0.63 | −0.40 | −0.51 | −0.45 | −0.49 | −0.48 | 0.33 | 1.00 | ||
| PCT | 0.75 | 0.39 | 0.49 | 0.38 | 0.42 | 0.50 | 0.46 | −0.55 | −0.23 | 1.00 | |
| NPT | 0.76 | 0.63 | 0.59 | 0.80 | 0.78 | 0.61 | 0.85 | −0.63 | −0.46 | 0.53 | 1.00 |
| Parameters | Pre/ Post-Treatment | HP | SAA | IL-1β | IFN-γ | TNF-α | MDA | GSH | SOD | PCT | NPT |
|---|---|---|---|---|---|---|---|---|---|---|---|
| HP | 0.80 | 1.00 | |||||||||
| SAA | 0.58 | 0.48 | 1.00 | ||||||||
| IL-1β | 0.63 | 0.63 | 0.48 | 1.00 | |||||||
| IFN-γ | 0.70 | 0.67 | 0.56 | 0.68 | 1.00 | ||||||
| TNF-α | 0.61 | 0.63 | 0.23 | 0.46 | 0.57 | 1.00 | |||||
| MDA | 0.76 | 0.69 | 0.53 | 0.70 | 0.76 | 0.56 | 1.00 | ||||
| GSH | −0.64 | −0.53 | −0.42 | −0.51 | −0.48 | −0.51 | −0.65 | 1.00 | |||
| SOD | −0.57 | 0.63 | −0.26 | −0.49 | −0.49 | −0.47 | −0.56 | 0.39 | 1.00 | ||
| PCT | 0.83 | 0.59 | 0.49 | 0.47 | 0.51 | 0.44 | 0.56 | −0.55 | −0.34 | 1.00 | |
| NPT | 0.85 | 0.73 | 0.54 | 0.73 | 0.76 | 0.58 | 0.85 | −0.63 | −0.53 | 0.70 | 1.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Deeb, W.; Iacob, O.; Fayez, M.; Elsohaby, I.; Alhaider, A.; Mkrtchyan, H.V.; Ibrahim, A.; Alhumam, N. Assessment of the Immune Response of Clinically Infected Calves to Cryptosporidium parvum Infection. Agriculture 2022, 12, 1151. https://doi.org/10.3390/agriculture12081151
El-Deeb W, Iacob O, Fayez M, Elsohaby I, Alhaider A, Mkrtchyan HV, Ibrahim A, Alhumam N. Assessment of the Immune Response of Clinically Infected Calves to Cryptosporidium parvum Infection. Agriculture. 2022; 12(8):1151. https://doi.org/10.3390/agriculture12081151
Chicago/Turabian StyleEl-Deeb, Wael, Olimpia Iacob, Mahmoud Fayez, Ibrahim Elsohaby, Abdulrahman Alhaider, Hermine V. Mkrtchyan, Abdelazim Ibrahim, and Naser Alhumam. 2022. "Assessment of the Immune Response of Clinically Infected Calves to Cryptosporidium parvum Infection" Agriculture 12, no. 8: 1151. https://doi.org/10.3390/agriculture12081151
APA StyleEl-Deeb, W., Iacob, O., Fayez, M., Elsohaby, I., Alhaider, A., Mkrtchyan, H. V., Ibrahim, A., & Alhumam, N. (2022). Assessment of the Immune Response of Clinically Infected Calves to Cryptosporidium parvum Infection. Agriculture, 12(8), 1151. https://doi.org/10.3390/agriculture12081151