
Development of KASP and SSR Markers for PmQ, a Recessive Gene Conferring Powdery Mildew Resistance in Wheat Landrace Qingxinmai

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
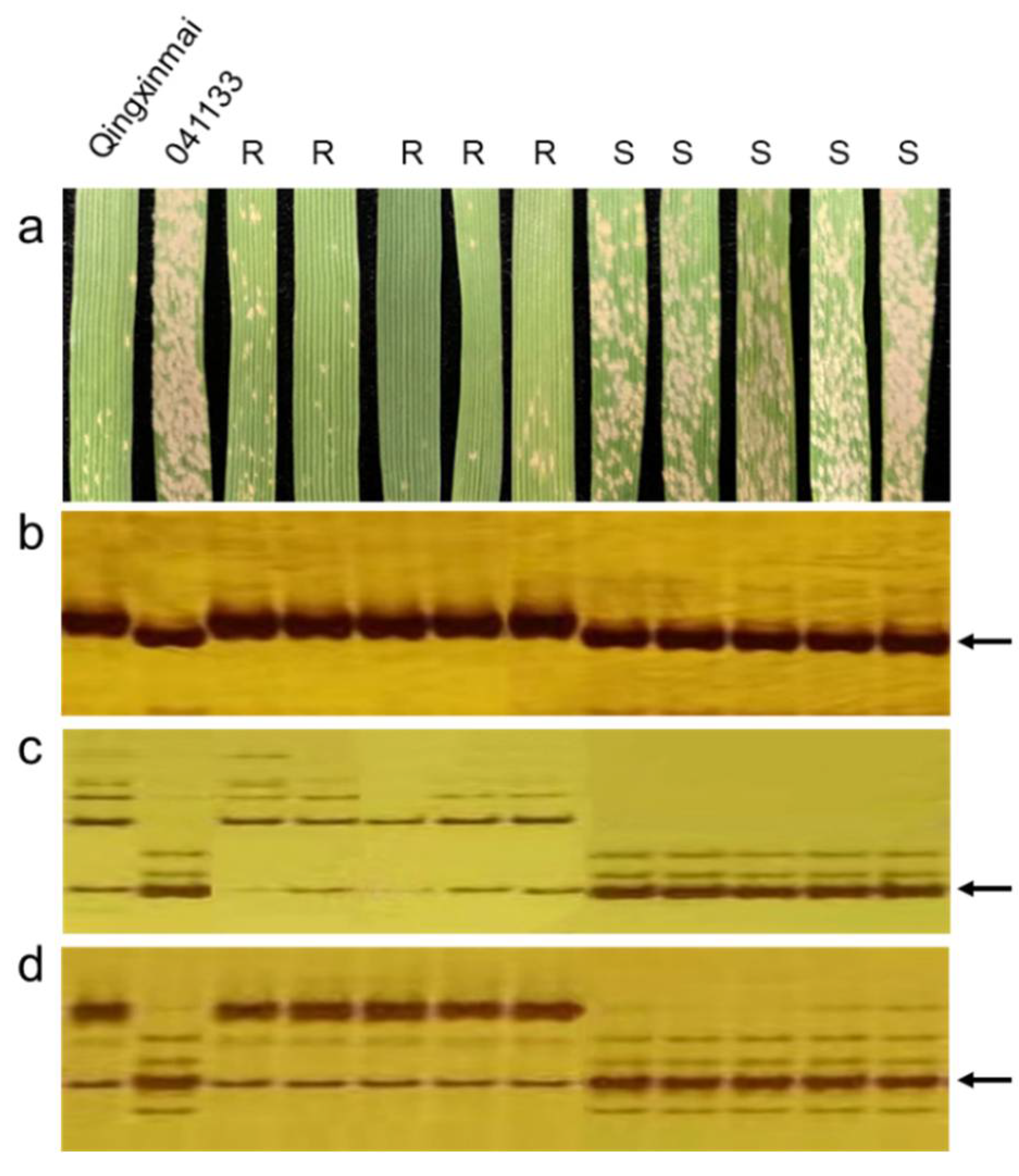
2.2. Assessment of Responses to Powdery Mildew
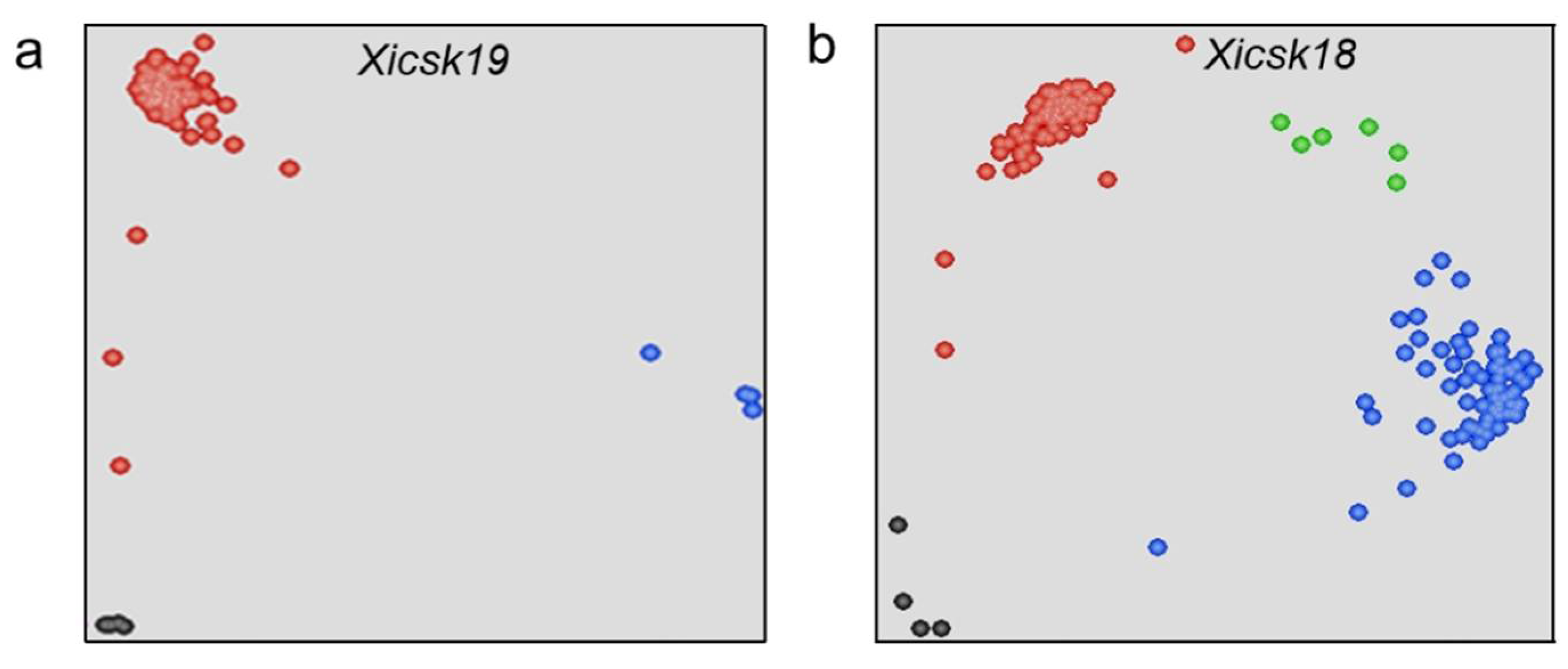
2.3. Development of KASP and SSR Primers
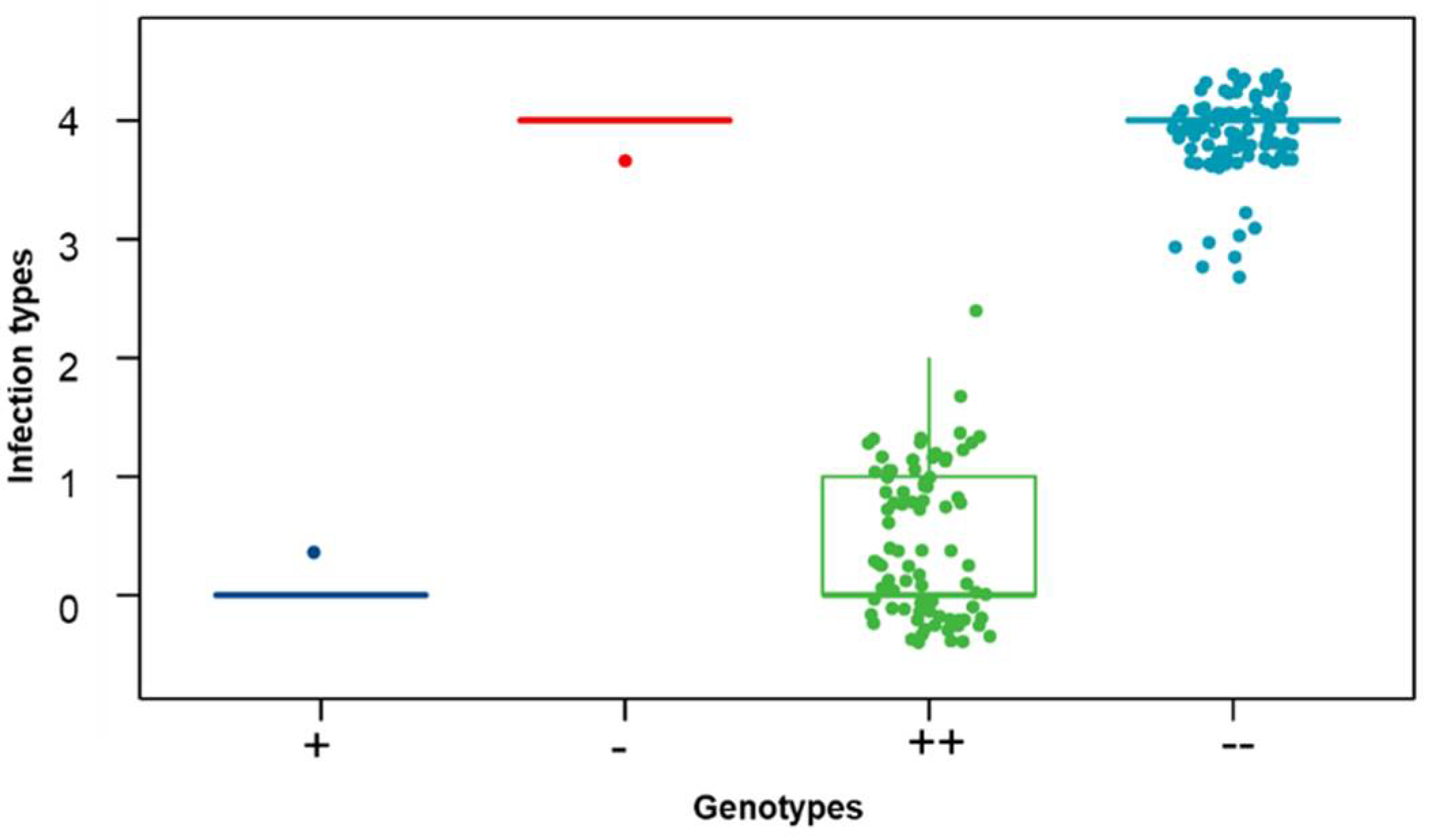
2.4. Identification Polymorphic KASP and SSR Markers
2.5. Genetic Linkage Map Construction and Statistical Analysis
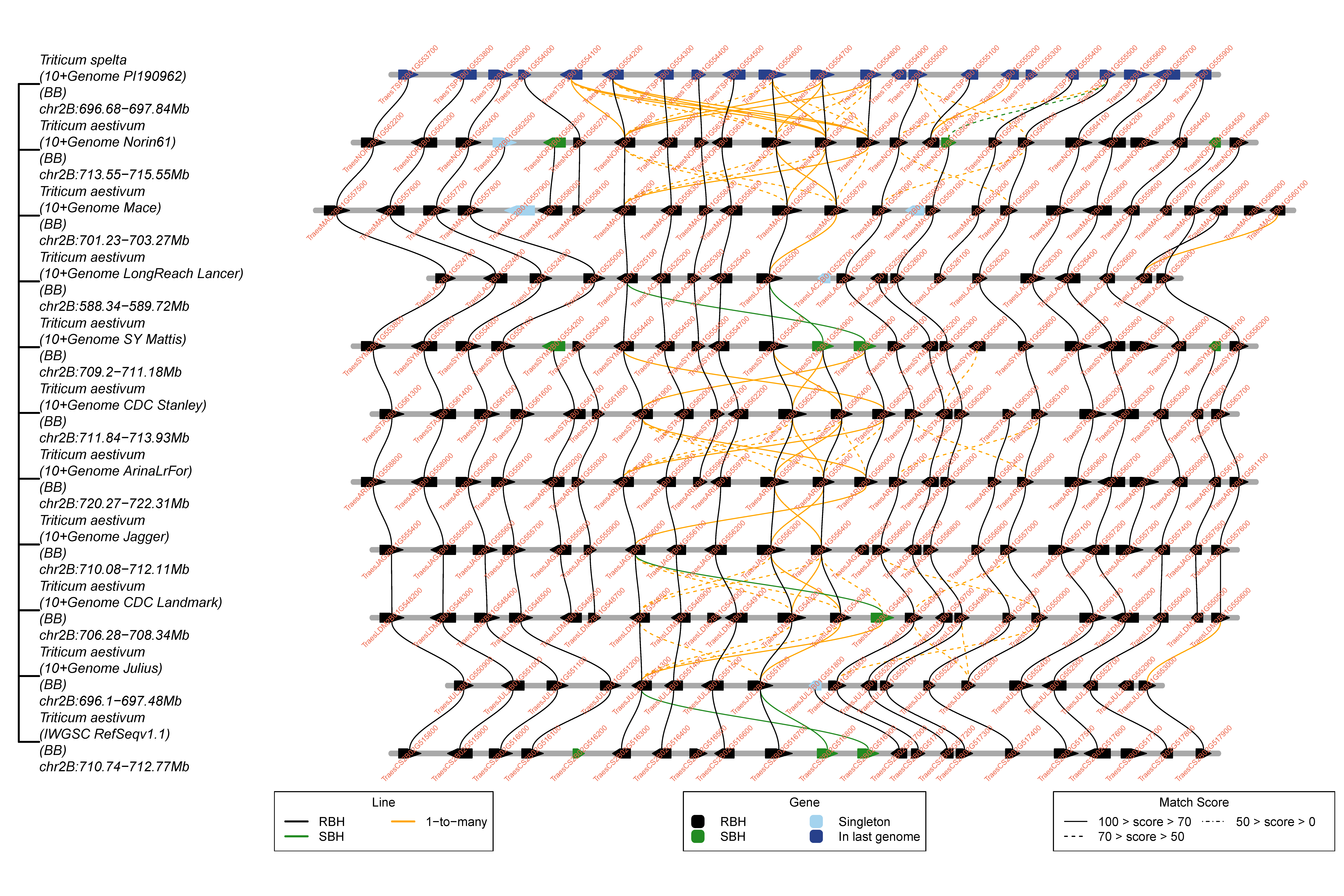
2.6. Analysis of Synteny for the Target Genomic Region
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shiferaw, B.; Smale, M.; Braun, H.-J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that Feed the World 10. Past Successes and Future Challenges to the Role Played by Wheat in Global Food Security. Food Secur. 2013, 5, 291–317. [Google Scholar] [CrossRef]
- Tolmay, V.L. Resistance to Biotic and Abiotic Stress in the Triticeae. Hereditas 2004, 135, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Afzal, F.; Chaudhari, S.K.; Gul, A.; Farooq, A.; Ali, H.; Nisar, S.; Sarfraz, B.; Shehzadi, K.J.; Mujeeb-Kazi, A. Bread Wheat (Triticum Aestivum L.) Under Biotic and Abiotic Stresses: An Overview. In Crop Production and Global Environmental Issues; Hakeem, K., Ed.; Springer: Cham, Switzerland, 2015; pp. 293–317. [Google Scholar]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The Global Burden of Pathogens and Pests on Major Food Crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Wicker, T.; Oberhaensli, S.; Parlange, F.; Buchmann, J.P.; Shatalina, M.; Roffler, S.; Ben-David, R.; Doležel, J.; Šimková, H.; Schulze-Lefert, P.; et al. The Wheat Powdery Mildew Genome Shows the Unique Evolution of An Obligate Biotroph. Nat. Genet. 2013, 45, 1092–1096. [Google Scholar] [CrossRef]
- Sotiropoulos, A.G.; Arango-Isaza, E.; Ban, T.; Barbieri, C.; Bourras, S.; Cowger, C.; Czembor, P.C.; Ben-David, R.; Dinoor, A.; Ellwood, S.R.; et al. Global Genomic Analyses of Wheat Powdery Mildew Reveal Association of Pathogen Spread with Historical Human Migration and Trade. Nat. Commun. 2022, 13, 4315. [Google Scholar] [CrossRef]
- Agrios, G.N. Plant Pathology, 5th ed.; Elsevier Academic Press Inc.: San Diego, CA, USA, 2005. [Google Scholar]
- Cunfer, B.M. Powdery mildew. In Bread Wheat: Improvement and Production; Curtis, B.C., Rajaram, S., Gómez Macpherson, H., Curtis, B.C., Rajaram, S., Gómez Macpherson, H., Eds.; Plant Production and Protection Series; FAO: Rome, Italy, 2002; Volume 30, pp. 317–330. [Google Scholar]
- Draz, I.S.; Esmail, S.M.; Abou-Zeid, M.A.E.-H.; Essa, T.A.E.-M. Powdery mildew susceptibility of spring wheat cultivars as a major constraint on grain yield. Ann. Agric. Sci. 2019, 64, 39–45. [Google Scholar] [CrossRef]
- Cao, X.-R.; Yao, D.-M.; Duan, X.-Y.; Liu, W.; Fan, J.-R.; Ding, K.-J.; Zhou, Y.-L. Effects of Powdery Mildew on 1000-Kernel Weight, Crude Protein Content and Yield of Winter Wheat in Three Consecutive Growing Seasons. J. Integr. Agric. 2014, 13, 1530–1537. [Google Scholar] [CrossRef]
- Gao, H.; Niu, J.; Yang, X.; He, D.; Wang, C. Impacts of Powdery Mildew on Wheat Grain Sugar Metabolism and Starch Accumulation in Developing Grains. Starch-Stärke 2014, 66, 947–958. [Google Scholar] [CrossRef]
- Huo, Z.G.; Chen, L.; Liu, W.C.; Xue, C.Y.; Zhao, S.J.; Zhuang, L.W. Climatic Zonation of Wheat Powdery Mildew in China. Acta Ecol. Sin. 2002, 22, 1873–1881. [Google Scholar]
- Ben-David, R.; Dinoor, A.; Peleg, Z.; Fahima, T. Reciprocal Hosts’ Responses to Powdery Mildew Isolates Originating from Domesticated Wheats and Their Wild Progenitor. Front. Plant Sci. 2018, 9, 75. [Google Scholar] [CrossRef]
- Huang, X.-Q.; Röder, M.S. Molecular Mapping of Powdery Mildew Resistance Genes in Wheat: A Review. Euphytica 2004, 137, 203–223. [Google Scholar] [CrossRef]
- Rasheed, A.; Wen, W.; Gao, F.; Zhai, S.; Jin, H.; Liu, J.; Guo, Q.; Zhang, Y.; Dreisigacker, S.; Xia, X.; et al. Development and Validation of KASP Assays for Genes Underpinning Key Economic Traits in Bread Wheat. Theor. Appl. Genet. 2016, 129, 1843–1860. [Google Scholar] [CrossRef] [PubMed]
- Majeed, U.; Darwish, E.; Rehman, S.U.; Zhang, X. Kompetitive Allele Specific PCR (KASP): A Singleplex Genotyping Platform and Its Application. J. Agric. Sci. 2018, 11, p11. [Google Scholar] [CrossRef]
- Makhoul, M.; Rambla, C.; Voss-Fels, K.P.; Hickey, L.; Snowdon, R.J.; Obermeier, C. Overcoming polyploidy pitfalls: A user guide for effective SNP conversion into KASP markers in wheat. Theor. Appl. Genet. 2020, 133, 2413–2430. [Google Scholar] [CrossRef] [PubMed]
- Marone, D.; Russo, M.; Mores, A.; Ficco, D.; Laidò, G.; Mastrangelo, A.; Borrelli, G. Importance of Landraces in Cereal Breeding for Stress Tolerance. Plants 2021, 10, 1267. [Google Scholar] [CrossRef] [PubMed]
- Bhullar, N.K.; Street, K.; Mackay, M.; Yahiaoui, N.; Keller, B. Unlocking Wheat Genetic Resources for the Molecular Identification of Previously Undescribed Functional Alleles at the Pm3 Resistance Locus. Proc. Natl. Acad. Sci. USA 2009, 106, 9519–9524. [Google Scholar] [CrossRef]
- Xie, J.; Guo, G.; Wang, Y.; Hu, T.; Wang, L.; Li, J.; Qiu, D.; Li, Y.; Wu, Q.; Lu, P.; et al. A Rare Single Nucleotide Variant in Pm5e Confers Powdery Mildew Resistance in Common Wheat. New Phytol. 2020, 228, 1011–1026. [Google Scholar] [CrossRef]
- Lu, P.; Guo, L.; Wang, Z.; Li, B.; Li, J.; Li, Y.; Qiu, D.; Shi, W.; Yang, L.; Wang, N.; et al. A Rare Gain of Function Mutation in A Wheat Tandem Kinase Confers Resistance to Powdery Mildew. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Ma, H.; Kong, Z.; Fu, B.; Li, N.; Zhang, L.; Jia, H.; Ma, Z. Identification and Mapping of a New Powdery Mildew Resistance Gene on Chromosome 6D of Common Wheat. Theor. Appl. Genet. 2011, 123, 1099–1106. [Google Scholar] [CrossRef]
- Xiao, M.; Song, F.; Jiao, J.; Wang, X.; Xu, H.; Li, H. Identification of The Gene Pm47 on Chromosome 7BS Conferring Resistance to Powdery Mildew in the Chinese Wheat Landrace Hongyanglazi. Theor. Appl. Genet. 2013, 126, 1397–1403. [Google Scholar] [CrossRef]
- Tan, C.; Li, G.; Cowger, C.; Carver, B.F.; Xu, X. Characterization of Pm59, a Novel Powdery Mildew Resistance Gene in Afghanistan Wheat Landrace PI 181356. Theor. Appl. Genet. 2018, 131, 1145–1152. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Hu, J.; Song, W.; Qiu, D.; Cui, L.; Wu, P.; Zhang, H.; Liu, H.; Yang, L.; Qu, Y.; et al. Pm61: A Recessive Gene for Resistance to Powdery Mildew in Wheat Landrace Xuxusanyuehuang Identified by Comparative Genomics Analysis. Theor. Appl. Genet. 2018, 131, 2085–2097. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Li, G.; Cowger, C.; Carver, B.F.; Xu, X. Characterization of Pm63, a Powdery Mildew Resistance Gene in Iranian Landrace PI 628024. Theor. Appl. Genet. 2018, 132, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Shi, X.; Hu, J.; Wu, P.; Qiu, D.; Qu, Y.; Xie, J.; Wu, Q.; Zhang, H.; Yang, L.; et al. Identification of a Recessive Gene PmQ Conferring Resistance to Powdery Mildew in Wheat Landrace Qingxinmai Using BSR-Seq Analysis. Plant Dis. 2020, 104, 743–751. [Google Scholar] [CrossRef]
- International Wheat Genome Sequencing Consortium (IWGSC). Shifting the Limits in Wheat Research and Breeding Using a Fully Annotated Reference Genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef]
- Ramirez-Gonzalez, R.H.; Uauy, C.; Caccamo, M. PolyMarker: A Fast Polyploid Primer Design Pipeline: Figure 1. Bioinformatics 2015, 31, 2038–2039. [Google Scholar] [CrossRef]
- Ma, S.; Wang, M.; Wu, J.; Guo, W.; Chen, Y.; Li, G.; Wang, Y.; Shi, W.; Xia, G.; Fu, D.; et al. WheatOmics: A Platform Combining Multiple Omics Data to Accelerate Functional Genomics Studies in Wheat. Mol. Plant 2021, 14, 1965–1968. [Google Scholar] [CrossRef]
- Allen, G.C.; Flores-Vergara, M.A.; Krasynanski, S.; Kumar, S.; Thompson, W. A Modified Protocol for Rapid DNA Isolation from Plant Tissues Using Cetyltrimethylammonium Bromide. Nat. Protoc. 2006, 1, 2320–2325. [Google Scholar] [CrossRef]
- Liu, R.-H.; Meng, J.-L. MapDraw: A Microsoft Excel Macro for Drawing Genetic Linkage Maps Based on Given Genetic Linkage Data. Hereditas 2003, 25, 317–321. [Google Scholar]
- Kosambi, D.D. The Estimation of Map Distances from Recombination Values. Ann. Eugen. 1943, 12, 172–175. [Google Scholar] [CrossRef]
- Lincoln, S.E.; Daly, M.J.; Lander, E.S. Constructing Genetic Linkage Maps with MAPMAKER/EXP Version 3.0: A Tutorial and Reference Mannual, 3rd ed.; Whitehead Institute for Medical Research: Cambridge, MA, USA, 1993. [Google Scholar]
- Avni, R.; Nave, M.; Barad, O.; Baruch, K.; Twardziok, S.O.; Gundlach, H.; Hale, I.; Mascher, M.; Spannagl, M.; Wiebe, K.; et al. Wild Emmer Genome Architecture and Diversity Elucidate Wheat Evolution and Domestication. Science 2017, 357, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Maccaferri, M.; Harris, N.S.; Twardziok, S.O.; Pasam, R.K.; Gundlach, H.; Spannagl, M.; Ormanbekova, D.; Lux, T.; Prade, V.M.; Milner, S.G.; et al. Durum Wheat Genome Highlights Past Domestication Signatures and Future Improvement Targets. Nat. Genet. 2019, 51, 885–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, H.-Q.; Ma, B.; Shi, X.; Liu, H.; Dong, L.; Sun, H.; Cao, Y.; Gao, Q.; Zheng, S.; Li, Y.; et al. Genome Sequence of the Progenitor of Wheat a Subgenome Triticum Urartu. Nature 2018, 557, 424–428. [Google Scholar] [CrossRef] [PubMed]
- Mascher, M.; Gundlach, H.; Himmelbach, A.; Beier, S.; Twardziok, S.O.; Wicker, T.; Radchuk, V.; Dockter, C.; Hedley, P.E.; Russell, J.; et al. A Chromosome Conformation Capture Ordered Sequence of The Barley Genome. Nature 2017, 544, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Walkowiak, S.; Gao, L.; Monat, C.; Haberer, G.; Kassa, M.T.; Brinton, J.; Ramirez-Gonzalez, R.H.; Kolodziej, M.C.; Delorean, E.; Thambugala, D.; et al. Multiple Wheat Genomes Reveal Global Variation in Modern Breeding. Nature 2020, 588, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Xu, Y. Bulk Segregation Analysis in the NGS Era: A Review of Its Teenage Years. Plant J. 2021, 109, 1355–1374. [Google Scholar] [CrossRef] [PubMed]
- Li, J.T.; Liu, Z.J. Discovery and Mapping of Powdery Mildew Resistance Genes in Wheat Landraces in China. Hubei Agric. Sci. 2015, 54, 4113–4116. [Google Scholar]
- Yin, G.-H.; Li, G.-Y.; He, Z.-H.; Liu, J.-J.; Wang, H.; Xia, X.-C. Molecular Mapping of Powdery Mildew Resistance Gene in Wheat Cultivar Jimai 22. Acta Agron. Sin. 2009, 35, 1425–1431. [Google Scholar] [CrossRef]
- Qu, Y.-F.; Wu, P.-P.; Hu, J.-H.; Chen, Y.-X.; Shi, Z.-L.; Qiu, D.; Li, Y.-H.; Zhang, H.-J.; Zhou, Y.; Yang, L.; et al. Molecular Detection of the Powdery Mildew Resistance Genes in Winter Wheats DH51302 and Shimai 26. J. Integr. Agric. 2020, 19, 931–940. [Google Scholar] [CrossRef]
- Zheng, T.C.; Li, X.P.; Yang, G.Y.; Zhang, X.D.; Sun, Y.X.; Dong, D.D. Analysis of Main Characteristics of Breeding Characteristic Level of Yumai 51. Crops 2000, 5, 35–36. [Google Scholar]
- Jia, A.; Ren, Y.; Gao, F.; Yin, G.; Liu, J.; Guo, L.; Zheng, J.; He, Z.; Xia, X. Mapping and Validation of a New QTL For Adult-Plant Resistance to Powdery Mildew in Chinese Elite Bread Wheat Line Zhou8425B. Theor. Appl. Genet. 2018, 131, 1063–1071. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Forward Primer-1 | Forward Primer-2 | Reverse Primer | Marker Type | R/S Allele and SSR Motif | Product Size (bp) | Physical Position (Mb) |
|---|---|---|---|---|---|---|---|
| Xicsk18 | TCCAAGGACTGATGGTACTGC | TCCAAGGACTGATGGTACTGG | TGGTTCAACAGAAGAGGCGT | KASP | G/C | 56 | 712.8 |
| Xicsk19 | GGTTAGCAACATCACCATTCAGC | GGTTAGCAACATCACCATTCAGT | GCCGAGAAAGATACCCATGTTT | KASP | C/T | 103 | 723.5 |
| Xicsk29 | GGCCATCATCTCTTTGTTTCCA | GGCCATCATCTCTTTGTTTCCG | GACAGGAAGGAGTAGCAGTC | KASP | T/C | 80 | 731.9 |
| Xicsqa | CGAGCATATATACTCCGAGCCG | - | AGCATGTGTATTCTTCAGACCGA | SSR | (AT)6 | 273 | 714.9 |
| Xicsqc | AGGAACGGGAAAGATGTCACA | - | CAGGACTGGCAGCACTCT | SSR | (CT)5 | 292 | 712.7 |
| Xicsqd | CATGAGACAAGGTGGTATGCC | - | AACGGCTGAAGTATTCCAGG | SSR | (CT)5 | 294 | 712.7 |
| Xicsqg | GCCACCGTAAATCTTTAGTCCG | - | TCACAAGAATCAAGCCCTACCT | SSR | (TTGTT)5 | 283 | 712.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gebremedhin, H.G.; Li, Y.; Hu, J.; Qiu, D.; Wu, Q.; Zhang, H.; Yang, L.; Zhou, Y.; Zhou, Y.; Liu, Z.; et al. Development of KASP and SSR Markers for PmQ, a Recessive Gene Conferring Powdery Mildew Resistance in Wheat Landrace Qingxinmai. Agriculture 2022, 12, 1344. https://doi.org/10.3390/agriculture12091344
Gebremedhin HG, Li Y, Hu J, Qiu D, Wu Q, Zhang H, Yang L, Zhou Y, Zhou Y, Liu Z, et al. Development of KASP and SSR Markers for PmQ, a Recessive Gene Conferring Powdery Mildew Resistance in Wheat Landrace Qingxinmai. Agriculture. 2022; 12(9):1344. https://doi.org/10.3390/agriculture12091344
Chicago/Turabian StyleGebremedhin, Habteab Goitom, Yahui Li, Jinghuang Hu, Dan Qiu, Qiuhong Wu, Hongjun Zhang, Li Yang, Yang Zhou, Yijun Zhou, Zhiyong Liu, and et al. 2022. "Development of KASP and SSR Markers for PmQ, a Recessive Gene Conferring Powdery Mildew Resistance in Wheat Landrace Qingxinmai" Agriculture 12, no. 9: 1344. https://doi.org/10.3390/agriculture12091344
APA StyleGebremedhin, H. G., Li, Y., Hu, J., Qiu, D., Wu, Q., Zhang, H., Yang, L., Zhou, Y., Zhou, Y., Liu, Z., Zhang, P., & Li, H. (2022). Development of KASP and SSR Markers for PmQ, a Recessive Gene Conferring Powdery Mildew Resistance in Wheat Landrace Qingxinmai. Agriculture, 12(9), 1344. https://doi.org/10.3390/agriculture12091344