
The Metarhizium anisopliae Strains Expressing dsRNA of the NlCHSA Enhance Virulence to the Brown Planthopper Nilaparvata lugens



Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungus Strains and Insect Culture
2.2. Vector Construction and Fungal Transformation
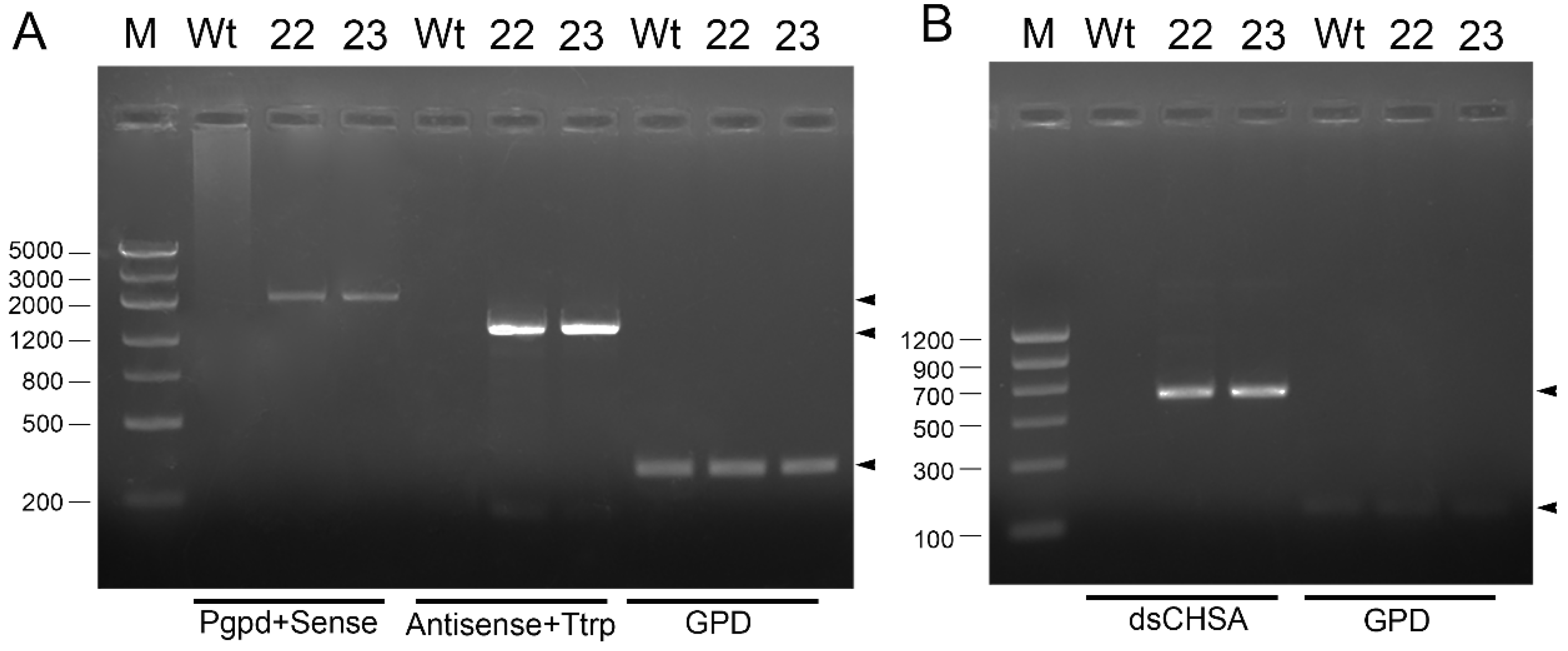
2.3. Verification of Transformants
2.4. Insect Bioassays
2.5. qPCR
2.6. Statistical Analysis
3. Results
3.1. Transgenic M. anisopliae Strains Expressed dsRNA of the NlCHSA
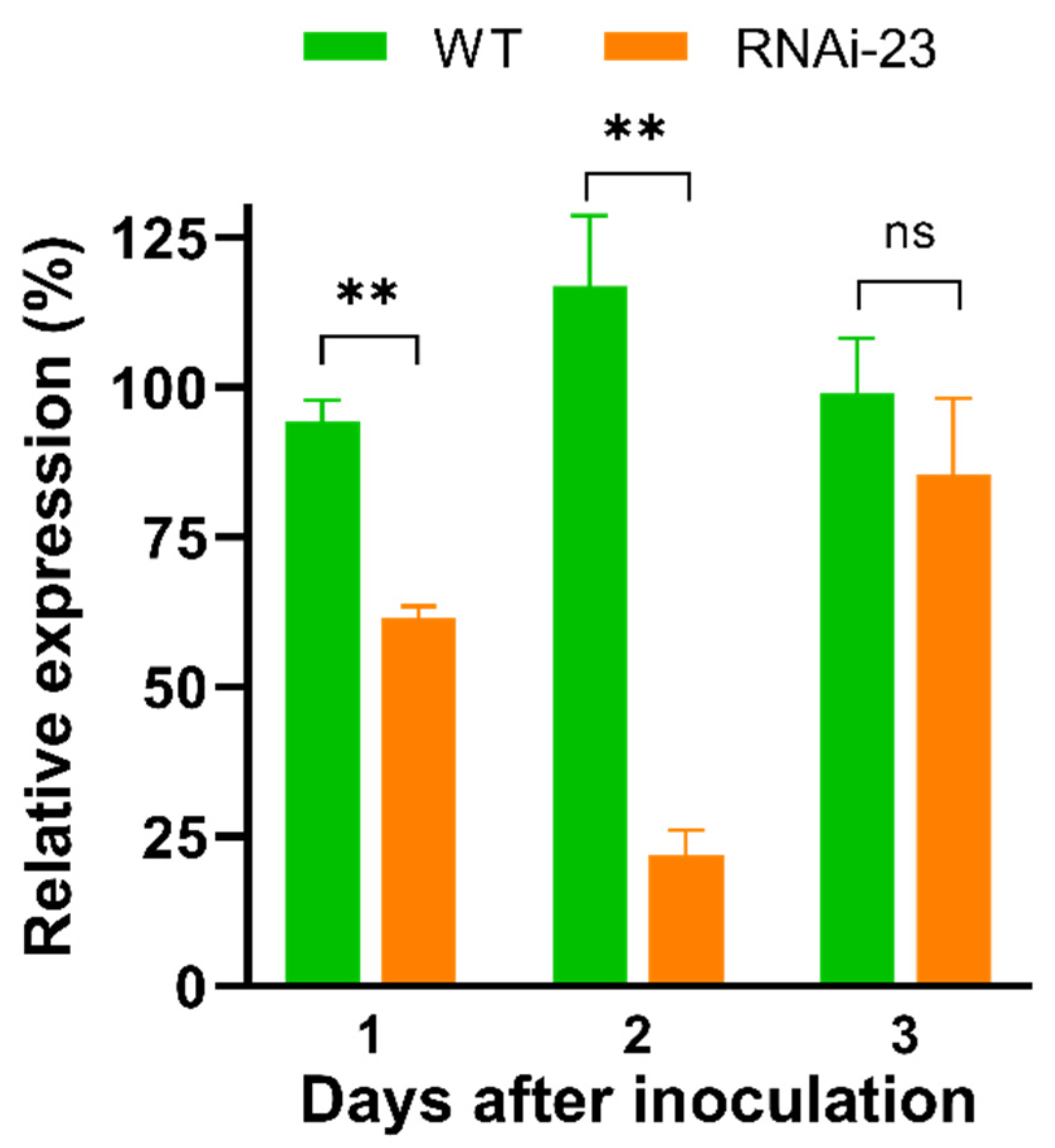
3.2. Transgenic M. anisopliae Strain Decreases NlCHSA Expression in N. lugens
3.3. Transgenic M. anisopliae Strains Enhance Their Virulence to N. lugens
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wang, Y.; Chen, J.; Zhu, Y.C.; Ma, C.; Huang, Y.; Shen, J. Susceptibility to neonicotinoids and risk of resistance development in the brown planthopper, Nilaparvata lugens (Stal) (Homoptera: Delphacidae). Pest Manag. Sci. 2008, 64, 1278–1284. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wu, H.; Chen, H.; Liu, Y.; He, J.; Kang, H.; Sun, Z.; Pan, G.; Wang, Q.; Hu, J.; et al. A gene cluster encoding lectin receptor kinases confers broad-spectrum and durable insect resistance in rice. Nat. Biotechnol. 2015, 33, 301–305. [Google Scholar] [CrossRef]
- Wu, J.; Ge, L.; Liu, F.; Song, Q.; Stanley, D. Pesticide-induced planthopper population resurgence in rice cropping systems. Annu. Rev. Entomol. 2020, 65, 409–429. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.W.; St Leger, R.J. Metarhizium spp., cosmopolitan insect-pathogenic fungi: Mycological aspects. Adv. Appl. Microbiol. 2004, 54, 1–70. [Google Scholar]
- Zimmermann, G. The entomopathogenic fungus Metarhizium anisopliae and its potential as a biocontrol agent. Pestic. Sci. 1993, 37, 375–379. [Google Scholar] [CrossRef]
- Tang, J.F.; Liu, X.Y.; Ding, Y.C.; Jiang, W.J.; Xie, J.Q. Evaluation of Metarhizium anisopliae for rice planthopper control and its synergy with selected insecticides. Crop Prot. 2019, 121, 132–138. [Google Scholar] [CrossRef]
- Jain, R.G.; Fletcher, S.J.; Manzie, N.; Robinson, K.E.; Li, P.; Lu, E.; Brosnan, C.A.; Xu, Z.P.; Mitter, N. Foliar application of clay-delivered RNA interference for whitefly control. Nat. Plants 2022, 8, 535–548. [Google Scholar] [CrossRef]
- Head, G.P.; Carroll, M.W.; Evans, S.P.; Rule, D.M.; Willse, A.R.; Clark, T.L.; Storer, N.P.; Flannagan, R.D.; Samuel, L.W.; Meinke, L.J. Evaluation of SmartStax and SmartStax PRO maize against western corn rootworm and northern corn rootworm: Efficacy and resistance management. Pest Manag. Sci. 2017, 73, 1883–1899. [Google Scholar] [CrossRef]
- Chen, X.; Li, L.; Hu, Q.; Zhang, B.; Wu, W.; Jin, F.; Jiang, J. Expression of dsRNA in recombinant Isaria fumosorosea strain targets the TLR7 gene in Bemisia tabaci. BMC Biotechnol. 2015, 15, 64. [Google Scholar] [CrossRef]
- Arakane, Y.; Muthukrishnan, S.; Kramer, K.J.; Specht, C.A.; Tomoyasu, Y.; Lorenzen, M.D.; Kanost, M.; Beeman, R.W. The Tribolium chitin synthase genes TcCHS1 and TcCHS2 are specialized for synthesis of epidermal cuticle and midgut peritrophic matrix. Insect Mol. Biol. 2005, 14, 453–463. [Google Scholar] [CrossRef]
- Wang, Y.; Fan, H.W.; Huang, H.J.; Xue, J.; Wu, W.J.; Bao, Y.Y.; Xu, H.J.; Zhu, Z.R.; Cheng, J.A.; Zhang, C.X. Chitin synthase 1 gene and its two alternative splicing variants from two sap-sucking insects, Nilaparvata lugens and Laodelphax striatellus (Hemiptera: Delphacidae). Insect Biochem. Mol. Biol. 2012, 42, 637–646. [Google Scholar] [CrossRef]
- Li, T.; Chen, J.; Fan, X.; Chen, W.; Zhang, W. MicroRNA and dsRNA targeting chitin synthase A reveal a great potential for pest management of the hemipteran insect Nilaparvata lugens. Pest Manag. Sci. 2017, 73, 1529–1537. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, D.; Yao, Q.; Zhang, J.; Dong, X.; Tian, H.; Chen, J.; Zhang, W. Feeding-based RNA interference of a trehalose phosphate synthase gene in the brown planthopper, Nilaparvata lugens. Insect Mol. Biol. 2010, 19, 777–786. [Google Scholar] [CrossRef]
- Hu, J.; Xia, Y. Increased virulence in the locust-specific fungal pathogen Metarhizium acridum expressing dsRNAs targeting the host F1F0-ATPase subunit genes. Pest Manag. Sci. 2019, 75, 180–186. [Google Scholar] [CrossRef]
- Fang, W.G.; Zhang, Y.; Yang, X.; Zheng, X.; Duan, H.; Li, Y.; Pei, Y. Agrobacterium tumefaciens-mediated transformation of Beauveria bassiana using an herbicide resistance gene as a selection marker. J. Invertebr. Pathol. 2004, 85, 18–24. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, J.; Zhu, K.Y. Chitosan/double-stranded RNA nanoparticle-mediated RNA interference to silence chitin synthase genes through larval feeding in the African malaria mosquito (Anopheles gambiae). Insect Mol. Biol. 2010, 19, 683–693. [Google Scholar] [CrossRef]
- Ganbaatar, O.; Cao, B.; Zhang, Y.; Bao, D.; Bao, W.; Wuriyanghan, H. Knockdown of Mythimna separata chitinase genes via bacterial expression and oral delivery of RNAi effectors. BMC Biotechnol. 2017, 17, 9. [Google Scholar] [CrossRef]
- Shi, J.F.; Mu, L.L.; Chen, X.; Guo, W.C.; Li, G.Q. RNA interference of chitin synthase genes inhibits chitin biosynthesis and affects larval performance in Leptinotarsa decemlineata (Say). Int. J. Biol. Sci. 2016, 12, 1319–1331. [Google Scholar] [CrossRef]
- Tian, H.; Peng, H.; Yao, Q.; Chen, H.; Xie, Q.; Tang, B.; Zhang, W. Developmental control of a lepidopteran pest Spodoptera exigua by ingestion of bacteria expressing dsRNA of a non-midgut gene. PLoS ONE 2009, 4, e6225. [Google Scholar] [CrossRef]
- Zhao, Y.; Sui, X.; Xu, L.; Liu, G.; Lu, L.; You, M.; Xie, C.; Li, B.; Ni, Z.; Liang, R. Plant-mediated RNAi of grain aphid CHS1 gene confers common wheat resistance against aphids. Pest Manag. Sci. 2018, 74, 2754–2760. [Google Scholar] [CrossRef]
- Ye, C.; Jiang, Y.D.; An, X.; Yang, L.; Shang, F.; Niu, J.; Wang, J.J. Effects of RNAi-based silencing of chitin synthase gene on moulting and fecundity in pea aphids (Acyrthosiphon pisum). Sci. Rep. 2019, 9, 3694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.; Singh, N.D.; Li, L.; Zhang, X.; Daniell, H. Engineered chloroplast dsRNA silences cytochrome p450 monooxygenase, V-ATPase and chitin synthase genes in the insect gut and disrupts Helicoverpa armigera larval development and pupation. Plant Biotechnol. J. 2015, 13, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Rajula, J.; Karthi, S.; Mumba, S.; Pittarate, S.; Thungrabeab, M.; Krutmuang, P. Chapter 4—Current status and future prospects of entomopathogenic fungi: A potential source of biopesticides. In Recent Advancement in Microbial Biotechnology; De Mandal, S., Passari, A.K., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 71–98. [Google Scholar]
- Whitten, M.M.A.; Facey, P.D.; Sol, R.D.; Fernández-Martínez, L.T.; Evans, M.C.; Mitchell, J.J.; Bodger, O.G.; Dyson, P.J. Symbiont-mediated RNA interference in insects. Proc. R. Soc. B 2016, 283, 20160042. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′–3′) |
|---|---|
| Primers used in construction of vectors and transformant verification | |
| GPD-F | GTAAAACGACGGCCAGTGCCAGTTGCCCCGGTACAGGTGG |
| GPD-R | GCCTTCTCAGCCTTGTCTTCTTTGCGTGTGTGTATATGGA |
| CHSA-F | TCCATATACACACACGCAAAGAAGACAAGGCTGAGAAGGC |
| CHSA-R | GAGTCGTGTCAGCACAGCGAGCACATGCTTGAATTGGTTAGGGTTAACGGAATCGTTTGCGGAAGGC |
| R-CHSA-F | TTGCATGCCTGCAGGTCGACTGGATCCTCGCTGTGCTGACACGACTCACCGGAATCGTTTGCGGAAGGC |
| R-CHSA-R | CAGTAACGTTAAGTGGATCGAAGACAAGGCTGAGAAGG |
| TrpC-F | CCTTCTCAGCCTTGTCTTCGATCCACTTAACGTTACTG |
| TrpC-R | TCAATGTCATCTTCTGTCGACGAGTGGAGATGTGGAGTGG |
| Intron-F | CCCTAACCAATTCAAGCAT |
| Intron-R | ATGCTTGAATTGGTTAGGG |
| MaGPD-F | CATCGATACTGAATACGC |
| MaGPD-R | AGTGAAGAACTTGACCTT |
| Primers used in RT-qPCR | |
| QNlCHSA-F | AAACTTTGGAGGCAATGGA |
| QNlCHSA-R | GCTGCTCGACGAAGTGGTG |
| qNlβ_actin-F | TGCGTGACATCAAGGAGAAGC |
| qNlβ_actin-R | CCATACCCAAGAAGGAAGGCT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Cui, H.; Hong, M.; Xia, Y.; Zhang, W. The Metarhizium anisopliae Strains Expressing dsRNA of the NlCHSA Enhance Virulence to the Brown Planthopper Nilaparvata lugens. Agriculture 2022, 12, 1393. https://doi.org/10.3390/agriculture12091393
Hu J, Cui H, Hong M, Xia Y, Zhang W. The Metarhizium anisopliae Strains Expressing dsRNA of the NlCHSA Enhance Virulence to the Brown Planthopper Nilaparvata lugens. Agriculture. 2022; 12(9):1393. https://doi.org/10.3390/agriculture12091393
Chicago/Turabian StyleHu, Jun, He Cui, Mingsheng Hong, Yuxian Xia, and Wenqing Zhang. 2022. "The Metarhizium anisopliae Strains Expressing dsRNA of the NlCHSA Enhance Virulence to the Brown Planthopper Nilaparvata lugens" Agriculture 12, no. 9: 1393. https://doi.org/10.3390/agriculture12091393
APA StyleHu, J., Cui, H., Hong, M., Xia, Y., & Zhang, W. (2022). The Metarhizium anisopliae Strains Expressing dsRNA of the NlCHSA Enhance Virulence to the Brown Planthopper Nilaparvata lugens. Agriculture, 12(9), 1393. https://doi.org/10.3390/agriculture12091393