


Physiological Alterations and Nondestructive Test Methods of Crop Seed Vigor: A Comprehensive Review

Abstract
:1. Introduction
2. Physiological Factors Affecting Seed Vigor
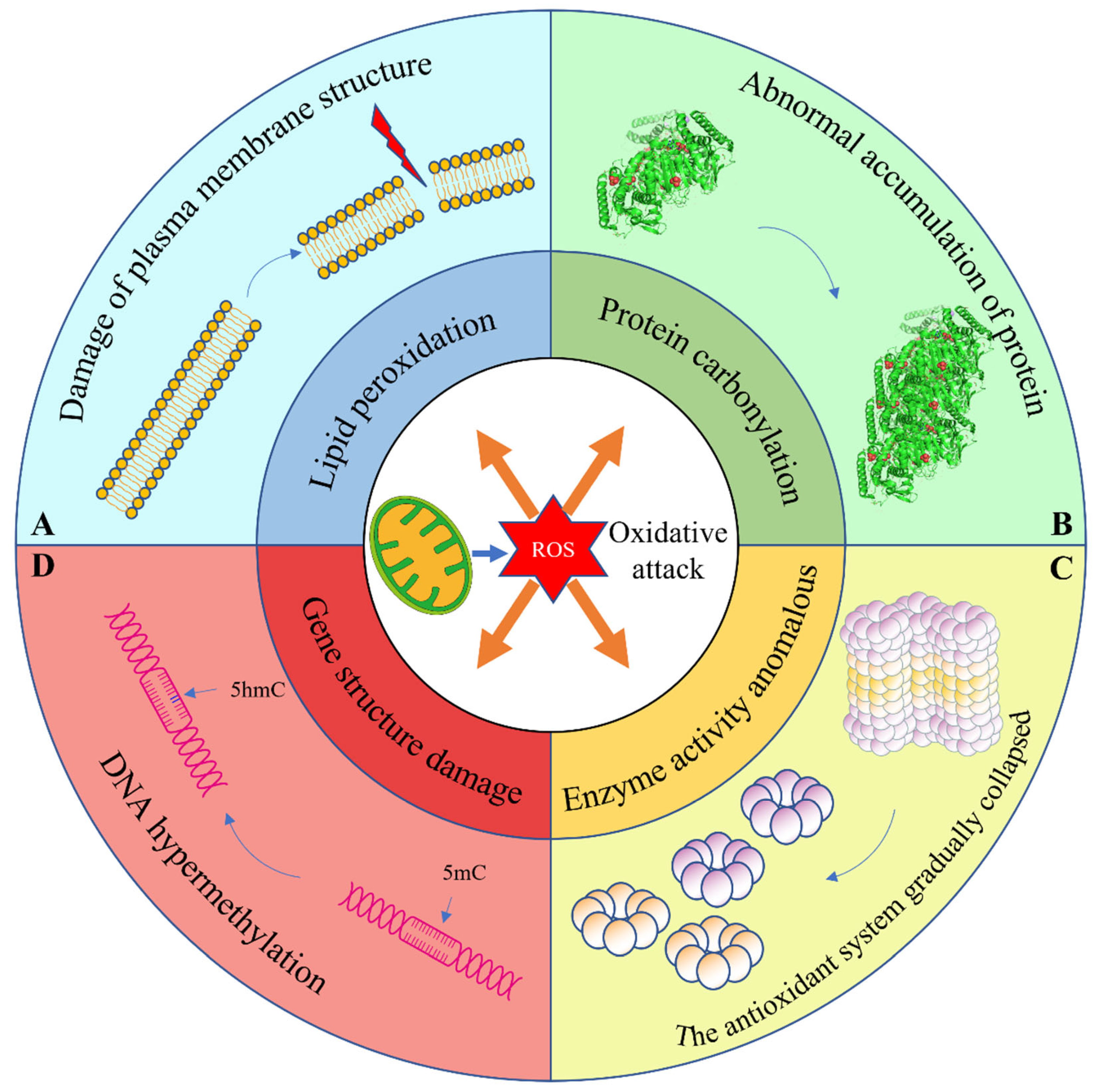
2.1. Production of Reactive Oxygen Species (ROS)
2.2. Lipid Peroxidation
2.3. Abnormal Protein Modification
2.4. Anomalous Enzyme Activity
2.5. Gene Damage
2.6. Destruction of Organelle Structure
3. The Relationship between Spectrum and Chemical
4. Current Testing Methods for Seed Vigor
4.1. Vibrational Spectroscopic Techniques
4.2. Biological Speckle Laser
4.3. Other Novel Means
5. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Nomenclature
| ROS | Reactive Oxygen Species |
| HSI | Hyperspectral Imaging Technology |
| TTC | 2,3,5-Triphenyltetrazolium Chloride |
| MDA | Malondialdehyde |
| HNE | 4-Hydroxy-Non-2-Enal |
| NA | Natural Aging |
| AAa | Artificial Aging |
| AAb | Accelerated Aging |
| CD | Conjugated Diene |
| FFA | Free Fatty Acid |
| LOOH | Lipid Hydroperoxide |
| TL | Total Lipid |
| POL | Product Of Lipid |
| CAT | Catalase |
| SOD | Superoxide Dismutase |
| APX | Ascorbic Acid Peroxidase |
| GR | Glutathione Reductase |
| DHAR | Dehydroascorbic Acid Reductase |
| MDHAR | Monodehydroascorbate Reductase |
| GPX | Glutathione Peroxidase |
| POX | Peroxidase |
| PGI | Phosphohexose Isomerase |
| MDH | Malate Dehydrogenase |
| qRT-PCR | Quantitative Real-Time Polymerase Chain Reaction |
| TCL | Thermochemilumenescence |
| HPLC-ESI-SIM | High Performance Liquid Chromatography-Electrospray Ionization-Selective Ion Monitoring. |
| DEG | Differentially Expressed Genes |
| NIR | Near Infrared Spectroscopy |
| VIS-NIR | Visible Near-Infrared Spectroscopy |
| SWIR | Short Wave Infrared Spectroscopy |
| ISTA | International Rules for Seed Testing by the International Seed Testing Association |
| TPF | 1,3,5-triphenylformazan |
| PLS-DA | Partial Least Squares Analysis |
| SIMCA | Similarity Classification |
| LS-SVM | Minimum Support Vector Machine |
| DL | Deep Learning Technology |
| K-NN | K-Nearest Neighbors |
| FI-NIR | Fourier Transform Infrared |
| CARS | Competitive Adaptive Reweighted Sampling |
| PLS-OLDA | Partial Least Squares-Orthogonal Linear Discriminant Analysis |
| SVM | Support Vector Machine |
| BPR | Biomimetic Pattern Recognition |
| MD | Mahalanobis Distance |
| LDA | Linear Discriminant Analysis |
| ATR-FITR | Attenuated Total Reflectance-Fourier Transform Infrared Spectroscopy |
| ELM | Extreme Learning Machine |
| DCNN | Deep Convolutional Neural Networks |
| DT | Decision Tree |
| VIP | Variable Importance of Projection |
| CNN | Convolutional Neural Networks |
| PLS-R | Partial Least Squares Regression |
| SPA | Successive Projection Algorithm |
| RC | Regression Coefficients |
| LR | Logistic Regression |
| DF | Deep Forests |
| RF | Random Forest |
| COM-IM | Moment of Inertia of Co-occurrence Matrix |
| GD | Generalized Differences |
| WE | Wavelet Entropy |
| Gr | Granularity |
| STD | Standard Deviation |
| MSF-BA | Modified structural Function-Biospeckle Activity |
| FTHSP | Full-Field Time History of Speckle Pattern |
| AVD | Absolute Value of Difference |
| TL | Transfer Learning |
| MCLA | 2-methyl-6-(p-methoxyphenyl)-3,7-dihydroimidazo [1,2α] pyrazin-3-one |
| ELS | Electrical Impedance Spectroscopy |
| GC-IMS | Gas Chromatography-Ion Mobility Spectrometry. |
| IDS | Incubation, Drying and Separation |
References
- Zhang, T.; Fan, S.; Xiang, Y.; Zhang, S.; Wang, J.; Sun, Q. Non-destructive analysis of germination percentage, germination energy and simple vigour index on wheat seeds during storage by Vis/NIR and SWIR hyperspectral imaging. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2020, 239, 118488. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M.; Gorain, J.; Pal, A.K.; Saha, B.; Chakraborti, P.; Avinash, B.; Pyngrope, D.M.; Banerjee, S.; Sarkar, S. Study of seed morphology and influence of ageing and storage conditions on germination and seedling vigour of non-Basmati aromatic rice. J. Stored Prod. Res. 2021, 93, 101863. [Google Scholar] [CrossRef]
- Yin, G.; Xin, X.; Song, C.; Chen, X.; Zhang, J.; Wu, S.; Li, R.; Liu, X.; Lu, X. Activity levels and expression of antioxidant enzymes in the ascorbate–glutathione cycle in artificially aged rice seed. Plant Physiol. Biochem. 2014, 80, 1–9. [Google Scholar] [CrossRef]
- Bailly, C. Active oxygen species and antioxidants in seed biology. Seed Sci. Res. 2004, 14, 93–107. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. REACTIVE OXYGEN SPECIES: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratajczak, E.; Małecka, A.; Bagniewska-Zadworna, A.; Kalemba, E.M. The production, localization and spreading of reactive oxygen species contributes to the low vitality of long-term stored common beech (Fagus sylvatica L.) seeds. J. Plant Physiol. 2015, 174, 147–156. [Google Scholar] [CrossRef]
- Xu, M.; He, D.; Teng, H.; Chen, L.; Song, H.; Huang, Q. Physiological and proteomic analyses of coix seed aging during storage. Food Chem. 2018, 260, 82–89. [Google Scholar] [CrossRef]
- Zhou, X.; Zhuang, Z.; Wang, W.; He, L.; Wu, H.; Cao, Y.; Pan, F.; Zhao, J.; Hu, Z.; Sekhar, C.; et al. OGG1 is essential in oxidative stress induced DNA demethylation. Cell. Signal. 2016, 28, 1163–1171. [Google Scholar] [CrossRef]
- Fan, Y.; Ma, S.; Wu, T. Individual wheat kernels vigor assessment based on NIR spectroscopy coupled with machine learning methodologies. Infrared Phys. Technol. 2020, 105, 103213. [Google Scholar] [CrossRef]
- Tian, X.; Zhang, C.; Li, J.; Fan, S.; Yang, Y.; Huang, W. Detection of early decay on citrus using LW-NIR hyperspectral reflectance imaging coupled with two-band ratio and improved watershed segmentation algorithm. Food Chem. 2021, 360, 130077. [Google Scholar] [CrossRef]
- Yang, Y.; Zhao, C.; Huang, W.; Tian, X.; Fan, S.; Wang, Q.; Li, J. Optimization and compensation of models on tomato soluble solids content assessment with online Vis/NIRS diffuse transmission system. Infrared Phys. Technol. 2022, 121, 104050. [Google Scholar] [CrossRef]
- Yang, Y.; Zhuang, H.; Yoon, S.-C.; Wang, W.; Jiang, H.; Jia, B. Rapid classification of intact chicken breast fillets by predicting principal component score of quality traits with visible/near-Infrared spectroscopy. Food Chem. 2018, 244, 184–189. [Google Scholar] [CrossRef]
- Long, Y.; Huang, W.; Wang, Q.; Fan, S.; Tian, X. Integration of textural and spectral features of Raman hyperspectral imaging for quantitative determination of a single maize kernel mildew coupled with chemometrics. Food Chem. 2022, 372, 131246. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Liang, X.; Huang, W.; Zhang, V.; Pang, Q.; He, X.; Li, L.; Zhang, C. Real-time defects detection for apple sorting using NIR cameras with pruning-based YOLOV4 network. Comput. Electron. Agric. 2022, 193, 106715. [Google Scholar] [CrossRef]
- Li, L.; Huang, W.; Wang, Z.; Liu, S.; He, X.; Fan, S. Calibration transfer between developed portable Vis/NIR devices for detection of soluble solids contents in apple. Postharvest Biol. Technol. 2022, 183, 111720. [Google Scholar] [CrossRef]
- Fan, S.; Li, J.; Zhang, Y.; Tian, X.; Wang, Q.; He, X.; Zhang, C.; Huang, W. On line detection of defective apples using computer vision system combined with deep learning methods. J. Food Eng. 2020, 286, 110102. [Google Scholar] [CrossRef]
- Ambrose, A.; Kandpal, L.M.; Kim, M.S.; Lee, W.-H.; Cho, B.-K. High speed measurement of corn seed viability using hyperspectral imaging. Infrared Phys. Technol. 2016, 75, 173–179. [Google Scholar] [CrossRef]
- Pang, L.; Wang, L.; Yuan, P.; Yan, L.; Xiao, J. Rapid seed viability prediction of Sophora japonica by improved successive projection algorithm and hyperspectral imaging. Infrared Phys. Technol. 2022, 123, 104143. [Google Scholar] [CrossRef]
- De Medeiros, A.D.; Bernardes, R.C.; da Silva, L.J.; de Freitas, B.A.L.; dos Santos Dias, D.C.F.; da Silva, C.B. Deep learning-based approach using X-ray images for classifying Crambe abyssinica seed quality. Ind. Crops Prod. 2021, 164, 113378. [Google Scholar] [CrossRef]
- Feng, L.; Hou, T.; Wang, B.; Zhang, B. Assessment of rice seed vigour using selected frequencies of electrical impedance spectroscopy. Biosyst. Eng. 2021, 209, 53–63. [Google Scholar] [CrossRef]
- De Melo, S.M.B.; Alves-de-Oliveira, D.F.; Souza, N.C.; Tavares-Silva, W.K.; de Macêdo, C.E.C.; Voigt, E.L. Oxidative status of Moringa oleifera Lam. seeds during storage. S. Afr. J. Bot. 2020, 129, 429–434. [Google Scholar] [CrossRef]
- Hu, D.; Ma, G.; Wang, Q.; Yao, J.; Wang, Y.; Pritchard, H.; Wang, X. Spatial and temporal nature of reactive oxygen species production and programmed cell death in elm (Ulmus pumila L.) seeds during controlled deterioration. Plant Cell Environ. 2012, 35, 2045–2059. [Google Scholar] [CrossRef]
- Oenel, A.; Fekete, A.; Krischke, M.; Faul, S.C.; Gresser, G.; Havaux, M.; Mueller, M.J.; Berger, S. Enzymatic and Non-Enzymatic Mechanisms Contribute to Lipid Oxidation During Seed Aging. Plant Cell Physiol. 2017, 58, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Rajjou, L.; Lovigny, Y.; Groot, S.P.C.; Belghazi, M.; Job, C.; Job, D. Proteome-Wide Characterization of Seed Aging in Arabidopsis: A Comparison between Artificial and Natural Aging Protocols. Plant Physiol. 2008, 148, 620–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehner, A.; Mamadou, N.; Poels, P.; Côme, D.; Bailly, C.; Corbineau, F. Changes in soluble carbohydrates, lipid peroxidation and antioxidant enzyme activities in the embryo during ageing in wheat grains. J. Cereal Sci. 2008, 47, 555–565. [Google Scholar] [CrossRef]
- Roberts, E. Predicting the storage life of seeds. Seed Sci. Technol. 1973, 1, 499–514. [Google Scholar]
- Ebone, L.A.; Caverzan, A.; Chavarria, G. Physiologic alterations in orthodox seeds due to deterioration processes. Plant Physiol. Biochem. 2019, 145, 34–42. [Google Scholar] [CrossRef]
- Huang, J.; Niazi, A.K.; Young, D.; Rosado, L.A.; Vertommen, D.; Bodra, N.; Abdelgawwad, M.R.; Vignols, F.; Wei, B.; Wahni, K.; et al. Self-protection of cytosolic malate dehydrogenase against oxidative stress in Arabidopsis. J. Exp. Bot. 2017, 69, 3491–3505. [Google Scholar] [CrossRef] [Green Version]
- El-Maarouf-Bouteau, H.; Bailly, C. Oxidative signaling in seed germination and dormancy. Plant Signal. Behav. 2008, 3, 175–182. [Google Scholar] [CrossRef] [Green Version]
- Adetunji, A.E.; Sershen; Varghese, B.; Pammenter, N. Effects of exogenous application of five antioxidants on vigour, viability, oxidative metabolism and germination enzymes in aged cabbage and lettuce seeds. S. Afr. J. Bot. 2021, 137, 85–97. [Google Scholar] [CrossRef]
- Miernyk, J.A.; Johnston, M.L.; Huber, S.C.; Tovar-Méndez, A.; Hoyos, E.; Randall, D.D. Oxidation of an Adjacent Methionine Residue Inhibits Regulatory Seryl-Phosphorylation of Pyruvate Dehydrogenase. Proteom. Insights 2009, 2, PRI.S2799. [Google Scholar] [CrossRef] [Green Version]
- Chandra, J.; Sershen, N.; Varghese, B.; Sahu, K. The Potential of ROS Inhibitors and Hydrated Storage in Improving the Storability of Recalcitrant Madhuca Latifolia Seeds. Seed Sci. Technol. 2019, 47, 33–45. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Y.; Xue, H.; Pritchard, H.W.; Wang, X. Changes in the mitochondrial protein profile due to ROS eruption during ageing of elm (Ulmus pumila L.) seeds. Plant Physiol. Biochem. 2017, 114, 72–87. [Google Scholar] [CrossRef]
- Su, D.; Wang, X.; Zhang, W.; Li, P.; Tang, B. Fluorescence imaging for visualizing the bioactive molecules of lipid peroxidation within biological systems. TrAC Trends Anal. Chem. 2022, 146, 116484. [Google Scholar] [CrossRef]
- Agmon, E.; Solon, J.; Bassereau, P.; Stockwell, B.R. Modeling the effects of lipid peroxidation during ferroptosis on membrane properties. Sci. Rep. 2018, 8, 5155. [Google Scholar] [CrossRef] [Green Version]
- Wong-ekkabut, J.; Xu, Z.; Triampo, W.; Tang, I.M.; Peter Tieleman, D.; Monticelli, L. Effect of Lipid Peroxidation on the Properties of Lipid Bilayers: A Molecular Dynamics Study. Biophys. J. 2007, 93, 4225–4236. [Google Scholar] [CrossRef] [Green Version]
- Leprince, O.; Atherton, N.; Deltour, R.; Hendry, C. The Involvement of Respiration in Free Radical Processes during Loss of Desiccation Tolerance in Germinating Zea mays L. (An Electron Paramagnetic Resonance Study). Plant Physiol. 1994, 104, 1333–1339. [Google Scholar] [CrossRef] [Green Version]
- Sahu, B.; Sahu, A.K.; Sahu, A.; Naithani, S.C. Gene expression of late embryogenesis abundant proteins, small heat shock proteins and peroxiredoxin and oxidation of lipid and protein during loss and re-establishment of desiccation tolerance in Pisum sativum seeds. S. Afr. J. Bot. 2018, 119, 28–36. [Google Scholar] [CrossRef]
- Zhang, T.; Ayed, C.; Fisk, I.D.; Pan, T.; Wang, J.; Yang, N.; Sun, Q. Evaluation of volatile metabolites as potential markers to predict naturally-aged seed vigour by coupling rapid analytical profiling techniques with chemometrics. Food Chem. 2022, 367, 130760. [Google Scholar] [CrossRef]
- Andrade, G.C.; Medeiros Coelho, C.M.; Uarrota, V.G. Modelling the vigour of maize seeds submitted to artificial accelerated ageing based on ATR-FTIR data and chemometric tools (PCA, HCA and PLS-DA). Heliyon 2020, 6, e03477. [Google Scholar] [CrossRef]
- Zhao, Y.; Tang, J.; Song, C.; Qi, S.; Lin, Q.; Cui, Y.; Ling, J.; Duan, Y. Nitric oxide alleviates chilling injury by regulating the metabolism of lipid and cell wall in cold-storage peach fruit. Plant Physiol. Biochem. 2021, 169, 63–69. [Google Scholar] [CrossRef]
- Fotouo-M, H.; Vorster, J.; du Toit, E.S.; Robbertse, P.J. The effect of natural long-term packaging methods on antioxidant components and malondialdehyde content and seed viability Moringa oleifera oilseed. S. Afr. J. Bot. 2020, 129, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Zacheo, G.; Cappello, M.S.; Gallo, A.; Santino, A.; Cappello, A.R. Changes Associated with Post-harvest Ageing in Almond Seeds. LWT Food Sci. Technol. 2000, 33, 415–423. [Google Scholar] [CrossRef]
- Hsu, C.C.; Chen, C.L.; Chen, J.J.; Sung, J.M. Accelerated aging-enhanced lipid peroxidation in bitter gourd seeds and effects of priming and hot water soaking treatments. Sci. Hortic. 2003, 98, 201–212. [Google Scholar] [CrossRef]
- Sahu, B.; Sahu, A.K.; Thomas, V.; Naithani, S.C. Reactive oxygen species, lipid peroxidation, protein oxidation and antioxidative enzymes in dehydrating Karanj (Pongamia pinnata) seeds during storage. S. Afr. J. Bot. 2017, 112, 383–390. [Google Scholar] [CrossRef]
- Kammoun, M.; Essid, M.F.; Ksouri, F.; Rokka, V.-M.; Charfeddine, M.; Gargouri-Bouzid, R.; Nouri-Ellouz, O. Assessment of physiological age and antioxidant status of new somatic hybrid potato seeds during extended cold storage. J. Plant Physiol. 2020, 254, 153279. [Google Scholar] [CrossRef]
- Nisar, F.; Gul, B.; Khan, M.A.; Hameed, A. Heteromorphic seeds of coastal halophytes Arthrocnemum macrostachyum and A. indicum display differential patterns of hydrogen peroxide accumulation, lipid peroxidation and antioxidant activities under increasing salinity. Plant Physiol. Biochem. 2019, 144, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Parkhey, S.; Naithani, S.C.; Keshavkant, S. ROS production and lipid catabolism in desiccating Shorea robusta seeds during aging. Plant Physiol. Biochem. 2012, 57, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Veselova, T.V.; Veselovsky, V.A.; Obroucheva, N.V. Deterioration mechanisms in air-dry pea seeds during early aging. Plant Physiol. Biochem. 2015, 87, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Yin, G.; Xin, X.; Wu, S.; Wei, X.; Lu, X. Levels of Crotonaldehyde and 4-hydroxy-(E)-2-nonenal and Expression of Genes Encoding Carbonyl-Scavenging Enzyme at Critical Node During Rice Seed Aging. Rice Sci. 2018, 25, 152–160. [Google Scholar]
- Xia, F.; Wang, X.; Li, M.; Mao, P. Mitochondrial structural and antioxidant system responses to aging in oat (Avena sativa L.) seeds with different moisture contents. Plant Physiol. Biochem. 2015, 94, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Suresh, A.; Shah, N.; Kotecha, M.; Robin, P. Evaluation of biochemical and physiological changes in seeds of Jatropha curcas L. Under natural aging, accelerated aging and saturated salt accelerated aging. Sci. Hortic. 2019, 255, 21–29. [Google Scholar] [CrossRef]
- Nigam, M.; Mishra, A.P.; Salehi, B.; Kumar, M.; Sahrifi-Rad, M.; Coviello, E.; Iriti, M.; Sharifi-Rad, J. Accelerated ageing induces physiological and biochemical changes in tomato seeds involving MAPK pathways. Sci. Hortic. 2019, 248, 20–28. [Google Scholar] [CrossRef]
- Jiang, F.L.; Bo, L.P.; Xu, J.J.; Wu, Z. Changes in respiration and structure of non-heading Chinese cabbage seeds during gradual artificial aging. Sci. Hortic. 2018, 238, 14–22. [Google Scholar] [CrossRef]
- Chaengsakul, C.; Onwimol, D.; Kongsil, P.; Suwannarat, S. Ethanol production and mitochondrial-related gene expression of maize (Zea mays) seed during storage. J. Integr. Agric. 2019, 18, 2435–2445. [Google Scholar] [CrossRef]
- Cheng, H.; Ma, X.; Jia, S.; Li, M.; Mao, P. Transcriptomic analysis reveals the changes of energy production and AsA-GSH cycle in oat embryos during seed ageing. Plant Physiol. Biochem. 2020, 153, 40–52. [Google Scholar] [CrossRef]
- Yao, Z.; Liu, L.; Gao, F.; Rampitsch, C.; Reinecke, D.M.; Ozga, J.A.; Ayele, B.T. Developmental and seed aging mediated regulation of antioxidative genes and differential expression of proteins during pre- and post-germinative phases in pea. J. Plant Physiol. 2012, 169, 1477–1488. [Google Scholar] [CrossRef]
- Sharma, S.N.; Maheshwari, A.; Sharma, C.; Shukla, N. Gene expression patterns regulating the seed metabolism in relation to deterioration/ageing of primed mung bean (Vigna radiata L.) seeds. Plant Physiol. Biochem. 2018, 124, 40–49. [Google Scholar] [CrossRef]
- Gao, G.; Hu, J.; Zhang, X.; Zhang, F.; Li, M.; Wu, X. Transcriptome analysis reveals genes expression pattern of seed response to heat stress in Brassica napus L. Oil Crop Sci. 2021, 6, 87–96. [Google Scholar] [CrossRef]
- Liu, S.; Liu, W.; Lai, J.; Liu, Q.; Zhang, W.; Chen, Z.; Gao, J.; Song, S.; Liu, J.; Xiao, Y. OsGLYI3, a glyoxalase gene expressed in rice seed, contributes to seed longevity and salt stress tolerance. Plant Physiol. Biochem. 2022, 183, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Zavadskiy, S.; Sologova, S.; Moldogazieva, N. Oxidative distress in aging and age-related diseases: Spatiotemporal dysregulation of protein oxidation and degradation. Biochimie 2022, 195, 114–134. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Wu, X.; Wu, W. Effects of oxidative modification by malondialdehyde on the in vitro digestion properties of rice bran protein. J. Cereal Sci. 2021, 97, 103158. [Google Scholar] [CrossRef]
- Møller, I.M.; Rogowska-Wrzesinska, A.; Rao, R.S.P. Protein carbonylation and metal-catalyzed protein oxidation in a cellular perspective. J. Proteom. 2011, 74, 2228–2242. [Google Scholar] [CrossRef]
- Bertini, I.; Gray, H.B.; Stiefel, E.I.; Valentine, J.S. Dioxygen reactivity and toxicity. Biol. Inorg. Chem. Struct. React. 2007, 31–41. [Google Scholar]
- Srivalli, S.; Khanna-Chopra, R. Delayed wheat flag leaf senescence due to removal of spikelets is associated with increased activities of leaf antioxidant enzymes, reduced glutathione/oxidized glutathione ratio and oxidative damage to mitochondrial proteins. Plant Physiol. Biochem. 2009, 47, 663–670. [Google Scholar] [CrossRef]
- McDonald, M. Seed deterioration: Physiology, repair and assessment. Seed Sci. Technol. 1999, 27, 177–237. [Google Scholar]
- Wang, Y.; Cui, Y.; Hu, G.; Wang, X.; Chen, H.; Shi, Q.; Xiang, J.; Zhang, Y.; Zhu, D.; Zhang, Y. Reduced bioactive gibberellin content in rice seeds under low temperature leads to decreased sugar consumption and low seed germination rates. Plant Physiol. Biochem. 2018, 133, 1–10. [Google Scholar] [CrossRef]
- Van Loon, B.; Markkanen, E.; Hübscher, U. Oxygen as a friend and enemy: How to combat the mutational potential of 8-oxo-guanine. DNA Repair 2010, 9, 604–616. [Google Scholar] [CrossRef]
- Lindahl, T.; Barnes, D.E. Repair of endogenous DNA damage. Cold Spring Harb. Symp. Quant. Biol. 2000, 65, 127–133. [Google Scholar] [CrossRef]
- Dawlaty, M.M.; Breiling, A.; Le, T.; Barrasa, M.I.; Raddatz, G.; Gao, Q.; Powell, B.E.; Cheng, A.W.; Faull, K.F.; Lyko, F.; et al. Loss of Tet Enzymes Compromises Proper Differentiation of Embryonic Stem Cells. Dev. Cell 2014, 29, 102–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menezo, Y.J.R.; Silvestris, E.; Dale, B.; Elder, K. Oxidative stress and alterations in DNA methylation: Two sides of the same coin in reproduction. Reprod. BioMed. Online 2016, 33, 668–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiran, K.R.; Deepika, V.B.; Swathy, P.S.; Prasad, K.; Kabekkodu, S.P.; Murali, T.S.; Satyamoorthy, K.; Muthusamy, A. ROS-dependent DNA damage and repair during germination of NaCl primed seeds. J. Photochem. Photobiol. B Biol. 2020, 213, 112050. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Yang, X.; Wang, X.; Li, Q.; Guo, J.; Ma, T.; Zhao, C.; Tang, Y.; Qiao, L.; Wang, J.; et al. The sweetpotato β-amylase gene IbBAM1.1 enhances drought and salt stress resistance by regulating ROS homeostasis and osmotic balance. Plant Physiol. Biochem. 2021, 168, 167–176. [Google Scholar] [CrossRef]
- Khan, A.R.; Azhar, W.; Wu, J.; Ulhassan, Z.; Salam, A.; Zaidi, S.H.R.; Yang, S.; Song, G.; Gan, Y. Ethylene participates in zinc oxide nanoparticles induced biochemical, molecular and ultrastructural changes in rice seedlings. Ecotoxicol. Environ. Saf. 2021, 226, 112844. [Google Scholar] [CrossRef]
- Salam, A.; Khan, A.R.; Liu, L.; Yang, S.; Azhar, W.; Ulhassan, Z.; Zeeshan, M.; Wu, J.; Fan, X.; Gan, Y. Seed priming with zinc oxide nanoparticles downplayed ultrastructural damage and improved photosynthetic apparatus in maize under cobalt stress. J. Hazard. Mater. 2022, 423, 127021. [Google Scholar] [CrossRef]
- Spanò, C.; Muccifora, S.; Ruffini Castiglione, M.; Bellani, L.; Bottega, S.; Giorgetti, L. Polystyrene nanoplastics affect seed germination, cell biology and physiology of rice seedlings in-short term treatments: Evidence of their internalization and translocation. Plant Physiol. Biochem. 2022, 172, 158–166. [Google Scholar] [CrossRef]
- Kareem, H.A.; Hassan, M.U.; Zain, M.; Irshad, A.; Shakoor, N.; Saleem, S.; Niu, J.; Skalicky, M.; Chen, Z.; Guo, Z.; et al. Nanosized zinc oxide (n-ZnO) particles pretreatment to alfalfa seedlings alleviate heat-induced morpho-physiological and ultrastructural damages. Environ. Pollut. 2022, 303, 119069. [Google Scholar] [CrossRef]
- Speight, J.G. Lange’s Handbook of Chemistry; McGraw-Hill: New York, NY, USA, 2004. [Google Scholar]
- Wu, N.; Weng, S.; Chen, J.; Xiao, Q.; Zhang, C.; He, Y. Deep convolution neural network with weighted loss to detect rice seeds vigor based on hyperspectral imaging under the sample-imbalanced condition. Comput. Electron. Agric. 2022, 196, 106850. [Google Scholar] [CrossRef]
- Wang, Y.; Peng, Y.; Qiao, X.; Zhuang, Q. Discriminant analysis and comparison of corn seed vigor based on multiband spectrum. Comput. Electron. Agric. 2021, 190, 106444. [Google Scholar] [CrossRef]
- Jia, S.; Yang, L.; An, D.; Liu, Z.; Yan, Y.; Li, S.; Zhang, X.; Zhu, D.; Gu, J. Feasibility of analyzing frost-damaged and non-viable maize kernels based on near infrared spectroscopy and chemometrics. J. Cereal Sci. 2016, 69, 145–150. [Google Scholar] [CrossRef]
- Ambrose, A.; Lohumi, S.; Lee, W.-H.; Cho, B.K. Comparative nondestructive measurement of corn seed viability using Fourier transform near-infrared (FT-NIR) and Raman spectroscopy. Sens. Actuators B Chem. 2016, 224, 500–506. [Google Scholar] [CrossRef]
- Wakholi, C.; Kandpal, L.M.; Lee, H.; Bae, H.; Park, E.; Kim, M.S.; Mo, C.; Lee, W.-H.; Cho, B.-K. Rapid assessment of corn seed viability using short wave infrared line-scan hyperspectral imaging and chemometrics. Sens. Actuators B Chem. 2018, 255, 498–507. [Google Scholar] [CrossRef]
- Jin, B.; Qi, H.; Jia, L.; Tang, Q.; Gao, L.; Li, Z.; Zhao, G. Determination of viability and vigor of naturally-aged rice seeds using hyperspectral imaging with machine learning. Infrared Phys. Technol. 2022, 122, 104097. [Google Scholar] [CrossRef]
- Wang, Z.; Tian, X.; Fan, S.; Zhang, C.; Li, J. Maturity determination of single maize seed by using near-infrared hyperspectral imaging coupled with comparative analysis of multiple classification models. Infrared Phys. Technol. 2021, 112, 103596. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, H.; Rao, Z.; Ji, H. Hyperspectral imaging technology combined with deep forest model to identify frost-damaged rice seeds. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2020, 229, 117973. [Google Scholar] [CrossRef]
- Seo, Y.-W.; Ahn, C.; Lee, H.; Park, E.; Mo, C.; Cho, B.-K. Non-Destructive Sorting Techniques for Viable Pepper (Capsicum annuum L.) Seeds Using Fourier Transform Near-Infrared and Raman Spectroscopy. J. Biosyst. Eng. 2016, 41, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Matthews, S.; Bradnock, T. The detection of seed samples of wrinkle-seeded peas (Pisum sativum L.) of potentially low planting value. Proc. Int. Seed Test. Assoc. 1976, 32, 553–563. [Google Scholar]
- Yang, L.; Wen, B. Seed Quality. In Encyclopedia of Applied Plant Sciences, 2nd ed.; Thomas, B., Murray, B.G., Murphy, D.J., Eds.; Academic Press: Oxford, UK, 2017; pp. 553–563. [Google Scholar]
- Gupta, N.; Chinnappa, M.; Singh, P.M.; Kumar, R.; Sagar, V. Determination of the physio-biochemical changes occurring during seed development, maturation, and desiccation tolerance in Moringa oleifera Lam. S. Afr. J. Bot. 2022, 144, 430–436. [Google Scholar] [CrossRef]
- Afzal, I.; Jaffar, I.; Zahid, S.; Rehman, H.U.; Basra, S.M.A. Physiological and biochemical changes during hermetic storage of Moringa oleifera seeds. S. Afr. J. Bot. 2020, 129, 435–441. [Google Scholar] [CrossRef]
- Tanaka, J.; Kiyoshi, K.; Kadokura, T.; Suzuki, K.-i.; Nakayama, S. Elucidation of the enzyme involved in 2,3,5-triphenyl tetrazolium chloride (TTC) staining activity and the relationship between TTC staining activity and fermentation profiles in Saccharomyces cerevisiae. J. Biosci. Bioeng. 2021, 131, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Finney, E.; Norris, K. Determination of Moisture in Corn Kernels by Near-Infrared Transmittance Measurements. Trans. ASAE 1978, 21, 0581–0584. [Google Scholar] [CrossRef]
- Jiang, H.Y.; Zhu, Y.J.; Wei, L.; Dai, J.; Song, T.; Yan, Y.L.; Chen, S. Analysis of protein, starch and oil content of single intact kernels by near infrared reflectance spectroscopy (NIRS) in maize (Zea mays L.). Plant Breed. 2007, 126, 492–497. [Google Scholar] [CrossRef]
- Esteve Agelet, L.; Ellis, D.D.; Duvick, S.; Goggi, A.S.; Hurburgh, C.R.; Gardner, C.A. Feasibility of near infrared spectroscopy for analyzing corn kernel damage and viability of soybean and corn kernels. J. Cereal Sci. 2012, 55, 160–165. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Peng, Y.; Zhuang, Q.; Zhao, X. Feasibility analysis of NIR for detecting sweet corn seeds vigor. J. Cereal Sci. 2020, 93, 102977. [Google Scholar] [CrossRef]
- Mukasa, P.; Wakholi, C.; Mo, C.; Oh, M.; Joo, H.-J.; Suh, H.K.; Cho, B.-K. Determination of viability of Retinispora (Hinoki cypress) seeds using FT-NIR spectroscopy. Infrared Phys. Technol. 2019, 98, 62–68. [Google Scholar] [CrossRef]
- Ma, T.; Tsuchikawa, S.; Inagaki, T. Rapid and non-destructive seed viability prediction using near-infrared hyperspectral imaging coupled with a deep learning approach. Comput. Electron. Agric. 2020, 177, 105683. [Google Scholar] [CrossRef]
- Pang, L.; Wang, L.; Yuan, P.; Yan, L.; Yang, Q.; Xiao, J. Feasibility study on identifying seed viability of Sophora japonica with optimized deep neural network and hyperspectral imaging. Comput. Electron. Agric. 2021, 190, 106426. [Google Scholar] [CrossRef]
- Pang, L.; Men, S.; Yan, L.; Xiao, J. Rapid Vitality Estimation and Prediction of Corn Seeds Based on Spectra and Images Using Deep Learning and Hyperspectral Imaging Techniques. IEEE Access 2020, 8, 123026–123036. [Google Scholar] [CrossRef]
- Zhang, J.; Dai, L.; Cheng, F. Identification of Corn Seeds with Different Freezing Damage Degree Based on Hyperspectral Reflectance Imaging and Deep Learning Method. Food Anal. Methods 2021, 14, 389–400. [Google Scholar] [CrossRef]
- Feng, L.; Zhu, S.; Zhang, C.; Bao, Y.; Feng, X.; He, Y. Identification of Maize Kernel Vigor under Different Accelerated Aging Times Using Hyperspectral Imaging. Molecules 2018, 23, 3078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dainty, J.C. Introduction. In Laser Speckle and Related Phenomena; Dainty, J.C., Ed.; Springer: Berlin/Heidelberg, Germany, 1975; pp. 1–7. [Google Scholar]
- Singh, P.; Chatterjee, A.; Rajput, L.S.; Rana, S.; Kumar, S.; Nataraj, V.; Bhatia, V.; Prakash, S. Development of an intelligent laser biospeckle system for early detection and classification of soybean seeds infected with seed-borne fungal pathogen (Colletotrichum truncatum). Biosyst. Eng. 2021, 212, 442–457. [Google Scholar] [CrossRef]
- Sutton, D.B.; Punja, Z.K. Investigating biospeckle laser analysis as a diagnostic method to assess sprouting damage in wheat seeds. Comput. Electron. Agric. 2017, 141, 238–247. [Google Scholar] [CrossRef]
- Rahmanian, A.; Mireei, S.A.; Sadri, S.; Gholami, M.; Nazeri, M. Application of biospeckle laser imaging for early detection of chilling and freezing disorders in orange. Postharvest Biol. Technol. 2020, 162, 111118. [Google Scholar] [CrossRef]
- Toderi, M.A.; Riquelme, B.D.; Galizzi, G.E. An experimental approach to study the red blood cell dynamics in a capillary tube by biospeckle laser. Opt. Lasers Eng. 2020, 127, 105943. [Google Scholar] [CrossRef]
- Singh, P.; Chatterjee, A.; Bhatia, V.; Prakash, S. Application of laser biospeckle analysis for assessment of seed priming treatments. Comput. Electron. Agric. 2020, 169, 105212. [Google Scholar] [CrossRef]
- Braga, R.A.; Dal Fabbro, I.M.; Borem, F.M.; Rabelo, G.; Arizaga, R.; Rabal, H.J.; Trivi, M. Assessment of Seed Viability by Laser Speckle Techniques. Biosyst. Eng. 2003, 86, 287–294. [Google Scholar] [CrossRef] [Green Version]
- Passoni, I.; Dai Pra, A.; Rabal, H.; Trivi, M.; Arizaga, R. Dynamic speckle processing using wavelets based entropy. Opt. Commun. 2005, 246, 219–228. [Google Scholar] [CrossRef]
- Dai Pra, A.L.; Passoni, L.I.; Rabal, H. Evaluation of laser dynamic speckle signals applying granular computing. Signal Process. 2009, 89, 266–274. [Google Scholar] [CrossRef]
- Braga, R.A.; Cardoso, R.R.; Bezerra, P.S.; Wouters, F.; Sampaio, G.R.; Varaschin, M.S. Biospeckle numerical values over spectral image maps of activity. Opt. Commun. 2012, 285, 553–561. [Google Scholar] [CrossRef]
- Thakur, P.S.; Tiwari, B.; Kumar, A.; Gedam, B.; Bhatia, V.; Krejcar, O.; Dobrovolny, M.; Nebhen, J.; Prakash, S. Deep transfer learning based photonics sensor for assessment of seed-quality. Comput. Electron. Agric. 2022, 196, 106891. [Google Scholar] [CrossRef]
- Rivera, F.P.; Braga, R.A.; Iannetta, P.; Toorop, P. Sound as a qualitative index of speckle laser to monitor biological systems. Comput. Electron. Agric. 2019, 158, 271–277. [Google Scholar] [CrossRef]
- Thakur, P.S.; Chatterjee, A.; Rajput, L.S.; Rana, S.; Bhatia, V.; Prakash, S. Laser biospeckle technique for characterizing the impact of temperature and initial moisture content on seed germination. Opt. Lasers Eng. 2022, 153, 106999. [Google Scholar] [CrossRef]
- Cardoso, R.R.; Costa, A.G.; Nobre, C.M.B.; Braga, R.A. Frequency signature of water activity by biospeckle laser. Opt. Commun. 2011, 284, 2131–2136. [Google Scholar] [CrossRef] [Green Version]
- Al-Hammad, B.A.; Al-Ammari, B.S. Seed viability of five wild Saudi Arabian species by germination and X-ray tests. Saudi J. Biol. Sci. 2017, 24, 1424–1429. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.R.; Yasmin, J.; Collins, W.; Cho, B.-K. X-ray CT image analysis for morphology of muskmelon seed in relation to germination. Biosyst. Eng. 2018, 175, 183–193. [Google Scholar] [CrossRef]
- Ahmed, M.R.; Yasmin, J.; Wakholi, C.; Mukasa, P.; Cho, B.-K. Classification of pepper seed quality based on internal structure using X-ray CT imaging. Comput. Electron. Agric. 2020, 179, 105839. [Google Scholar] [CrossRef]
- Liu, X.; Gao, C.; Xing, D. A non-invasive and rapid seed vigor biosensor based on quantitative measurement of superoxide generated by aleurone cell in intact seeds. Biosens. Bioelectron. 2009, 24, 1537–1542. [Google Scholar] [CrossRef]
- Ornellas, F.L.S.; de Sousa, A.O.; Pirovani, C.P.; Araújo, M.N.; da Costa, D.S.; Dantas, B.F.; Barbosa, R.M. Gene expression, biochemical and physiological activities in evaluating melon seed vigor through ethanol release. Sci. Hortic. 2020, 261, 108884. [Google Scholar] [CrossRef]
- Daneshvar, A.; Tigabu, M.; Karimidoost, A.; Odén, P.C. Flotation techniques to improve viability of Juniperus polycarpos seed lots. J. For. Res. 2017, 28, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Demelash, L.; Tigabu, M.; Oden, P.C. Separation of empty and dead-filled seeds from a seed lot of Pinus Patula with IDS technique. Seed Sci. Technol. 2002, 30, 677–681. [Google Scholar]
- Demelash, L.; Tigabu, M.; Oden, P.C. Enhancing germinability of Schinus molle L. seed lot from Ethiopia with specific gravity and IDS techniques. New For. 2003, 26, 33–41. [Google Scholar] [CrossRef]
- Hendeniya, N.; Sandanuwan, T.; Amarasinghe, D.A.S.; Attygalle, D.; Weragoda, S.C.; Ranaweera, B.; Rathnayake, K.; Lalanka, M. Atmospheric Pressure Plasma Treatment as a Cost-Effective and Eco-Friendly Pre-Treatment Method to Enhance Seed Perfomance in Germination and Early Seedling Growth. In Proceedings of the 2021 Moratuwa Engineering Research Conference (MERCon), Moratuwa, Sri Lanka, 27–29 July 2021; pp. 643–648. [Google Scholar]
- Jorge, M.H.A.; Ray, D.T. Germination characterization of guayule seed by morphology, mass and, X-ray analysis. Ind. Crops Prod. 2005, 22, 59–63. [Google Scholar] [CrossRef]
- Lin, H.; Man, Z.-x.; Kang, W.-c.; Guan, B.-b.; Chen, Q.-s.; Xue, Z.-l. A novel colorimetric sensor array based on boron-dipyrromethene dyes for monitoring the storage time of rice. Food Chem. 2018, 268, 300–306. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Physiological Alternation | Type of Seed | Method of Treating Seeds | Target | Method of Analysis | Performance | Ref. |
|---|---|---|---|---|---|---|
| Lipid peroxidation | Almond seeds of 4 varieties | NA | MDA | Spectrophotometry | MDA increase | [44] |
| Bitter gourd | AA a | MDA increase | [45] | |||
| Wheat | AA a | No significant change (0.38–0.45) | [25] | |||
| Karanj | NA | Positive correlation with aging days (r = 0.96) | [46] | |||
| Pea | Drying | MDA increase | [39] | |||
| Moringa oleifera oilseed | AA a | MDA increase | [43] | |||
| Spunta | Controlled environmental conditions | MDA increased | [47] | |||
| A. indicum and A. macrostachyum | No special treatment | MDA increase | [48] | |||
| Brassica oleracea L. and Lactuca sativa L. | Controlled deterioration | The change of cabbage seeds was not obvious, lettuce seeds increased significantly | [30] | |||
| Shorea robusta | No special treatment | MDA; CD; HNE; FFA; LOOH; TL | Other: Spectrophotometry TL: Petroleum ether distillation | MDA: Positive correlation with aging days (r = 0.98) CD: Positive correlation with aging days (r = 0.92) HNE: Rose and then fell. FFA: Positive correlation with storage days (r = 0.96) LOOH: Positive correlation with storage days (r = 0.98) TL: Negatively correlated with storage days (r = −0.98) | [49] | |
| Pea | NA | POL | TCL | POL increased after seed death | [50] | |
| Rice | NA | Crotonaldehyde; H NE | HPLC-ESI-SIM | Increased | [51] | |
| Enzyme activity anomalous | Rice | AA a | SOD APX CAT GR DHAR MDHAR | Spectrophotometry | APX, CAT, MDHAR: Decreased SOD, GR, DHAR: No significant change | [3] |
| Beech | NA | CAT | Decreased | [6] | ||
| Oat | AA a | SOD APX MDHAR DHAR GR | Decreased | [52] | ||
| Karanj | NA | SOD CAT APX | Decreased | [46] | ||
| Jatropha curcas L. | AA a | SOD CAT POX | Decreased after rise | [53] | ||
| Tomato | AA a | CAT POD SOD APX GR DHAR | Decreased | [54] | ||
| Nonheading Chinese cabbage | AA a | PGI MDH | Decreased after rise | [55] | ||
| Bitter gourd | AA a | SOD CAT APX | Decreased | [45] | ||
| Wheat | AA a | SOD CAT GR | Decreased | [25] | ||
| Spunta | Controlled environmental conditions | SOD CAT GPX | Relatively stable | [46] | ||
| Moringa oleifera Lam. | NA | CAT SOD APX POX | CAT, APX, SOD: Decreased POX: Increased | [21] | ||
| Brassica oleracea L. and Lactuca sativa L. | Controlled deterioration treatment | CAT GR SOD α-amylase β−1,3-glucanase | α-amylase: dinitrosalicylic acid, β−1,3-glucanase: Spectrophotometry | Decreased | [30] | |
| Gene damage | Maize | NA and AA b | AOX1, ADH1, COXc, ATPase | Spectrophotometry | Upregulation of ADH1. Downregulation of COXc. ATPase and AOX1 is almost unchanged | [56] |
| Oat | Controlled deterioration treatment | DEGs | Downregulation of TCA and ETC related genes. Upregulation of antioxidant enzyme related DEG | [57] | ||
| Pea | AA b | PsAPX, PsSOD, PsGRcyt, PsGRcm | qRT-PCR | Decreased in the transcript abundance | [58] | |
| Rice | NA | OsAKR-1-OsAKR-3, OsALDH2-1- OsALDH2-5, OsALDH3-1- OsALDH3-5, OsALDH7 | Transcript decreased | [50] | ||
| AA a | Cu/ZnSOD1- Cu/ZnSOD3, FeSOD, MnSOD, CAT1-CAT3, APX1-APX8GR1-GR3 DHAR1, MDHAR1, MDHAR2 | Downregulation of Cu/ZnSOD family genes, CAT1, CAT2, APX1, APX4, APX5, APX6, GR2, DHAR1 and MDHAR1. Upregulation of APX3, APX7, APX8, GR3. | [3] | |||
| Beech | NA | Genomic DNA | DNA extraction and electrophoresis | DNA laddering effect | [6] | |
| Pisum sativum | Drying | PsDHN2, PsDHN3, PsSBP65, PsHSP17.7, PsHSP18.1, PsHSP18.2, PsHSP22.7, Ps2-Cys prx | qRT-PCR | Expression enhancement | [39] | |
| Mung bean | Controlled deterioration treatment | FSD1, MSD1, GST1, ZEP/ABA1, SDR, CPS1, SAM1, ACS | Downregulation of FSD1, SAM1. ACS. Upregulation of MSD1, ZEP/ABA1, SDR. | [59] | ||
| Brassica napus L. | High temperature | DEGs | 40 °C (83 upregulated and 37 downregulated). 60 °C (96 upregulated and 117 downregulated). 40/60 °C of heat stress exposure (88 upregulated and 158 downregulated) | [60] | ||
| Rice | AA a | OLGLYI3 | Decreased expression | [61] |
| Type of Spectral | Material | Object | Band | Remarks | Ref. |
|---|---|---|---|---|---|
| NIR | Wheat | Protein, water, and starch | 1200–2400 nm | Peak 1 (1265 nm, 1272 nm and 1278 nm,): Related to the starch and protein. Peak 2 (1912 nm and 2168 nm): Related to the protein. Peak 3 (2027 nm): Related to the starch. | [9] |
| NIR | Maize | Protein, carbohydrate, and water | 1000–2400 nm | Peak 1 (1180 nm and 1420 nm): Second overtone of C-H stretching. Peak 2 (1700 nm and 1748 nm): Overtone of C-H stretching. Peak 3 (1918 nm): Related to the carbohydrate. Peak 4 (2035 nm): Related to the protein, water and starch. Peak 5 (2058 nm): Overtone of N-H stretching. Peak 6 (2275 nm): Overtone of O-H stretching. | [83] |
| SWIR | Maize | Starch, lipid, cellulose, protein | 1100–1600 nm | Peak 1 (1220–1230 nm): Second overtone of C-H stretching (starch, lipid, cellulose). Peak 2 (1560–1570 nm): First overtone of N-H stretching (protein). | [84] |
| NIR-HSI | Rice | Protein | 900–1700 nm | Peak 1 (1000 nm): Second overtone of N-H stretching (protein). Peak 2 (1200 nm): Second overtone of C-H stretching. Peak 3 (1400 nm): First overtone of O-H stretching. | [85] |
| HSI | Maize | Starch. Lipid, protein, cellulose, sugar | 1000–2200 nm | Peak 1 (1100–1300 nm): Second overtone of C-H stretching (starch). Peak 2 (1450 nm): First overtone of O-H stretching (starch and lipid). Peak 3 (1700–1750 nm): Second overtone of C-H stretching (protein, starch, and lipid). Peak 4 (1800–2250 nm): Related to the protein, cellulose, and sugar. | [86] |
| HSI | Wheat | Protein, starch, lipid | 949–1638 nm | Peak 1 (1030.9 nm and 1047.9 nm): Third overtone of N-H stretching (protein). Peak 2 (1152.4 nm and 1334.9 nm): Second overtone of C-H stretching (starch). Peak 3 (1413.3 nm and 1529.6 nm): First overtone of O-H stretching (starch and lipid). | [87] |
| Raman | Maize | Protein, starch, lipid, cellulose | 170–3200 cm−1 | Peak 1 (964 cm−1): Related to the cellulose. Peak 2 (1660 cm−1): Related to the protein. Peak 3 (2800–3000 cm−1): Related to the lipid. | [83] |
| Raman | Pepper | Protein, carbohydrate | 150–1800 cm−1 | Peak 1 (1090 cm−1): Overtone of -CO stretching. Peak 2 (1154 cm−1): Overtone of C-C stretching. Peak 3 (1263 cm−1): Overtone of =CH stretching. Peak 4 (1440 cm−1): Overtone of =CH2 stretching. Peak 5 (1520 cm−1): Overtone of C = C stretching. | [88] |
| FI-NIR | Maize | Protein, starch, and carbohydrate | 1110–2500 nm | Peak 1 (1500–1700 nm) and Peak 2 (1800–2100 nm): Overtone of C-H, O-H and N-H stretching. | [82] |
| Vis-NIR | Maize | Protein, starch, lipid | 500–1100 nm. 1000–1850 nm | Peak 1 (750 nm): Third overtone of O-H stretching. Peak 2 (800 nm): Third overtone of N-H stretching. Peak 3 (900 nm): Third overtone of C-H stretching. Peak 4 (1100 nm): Second overtone of C-H stretching. | [81] |
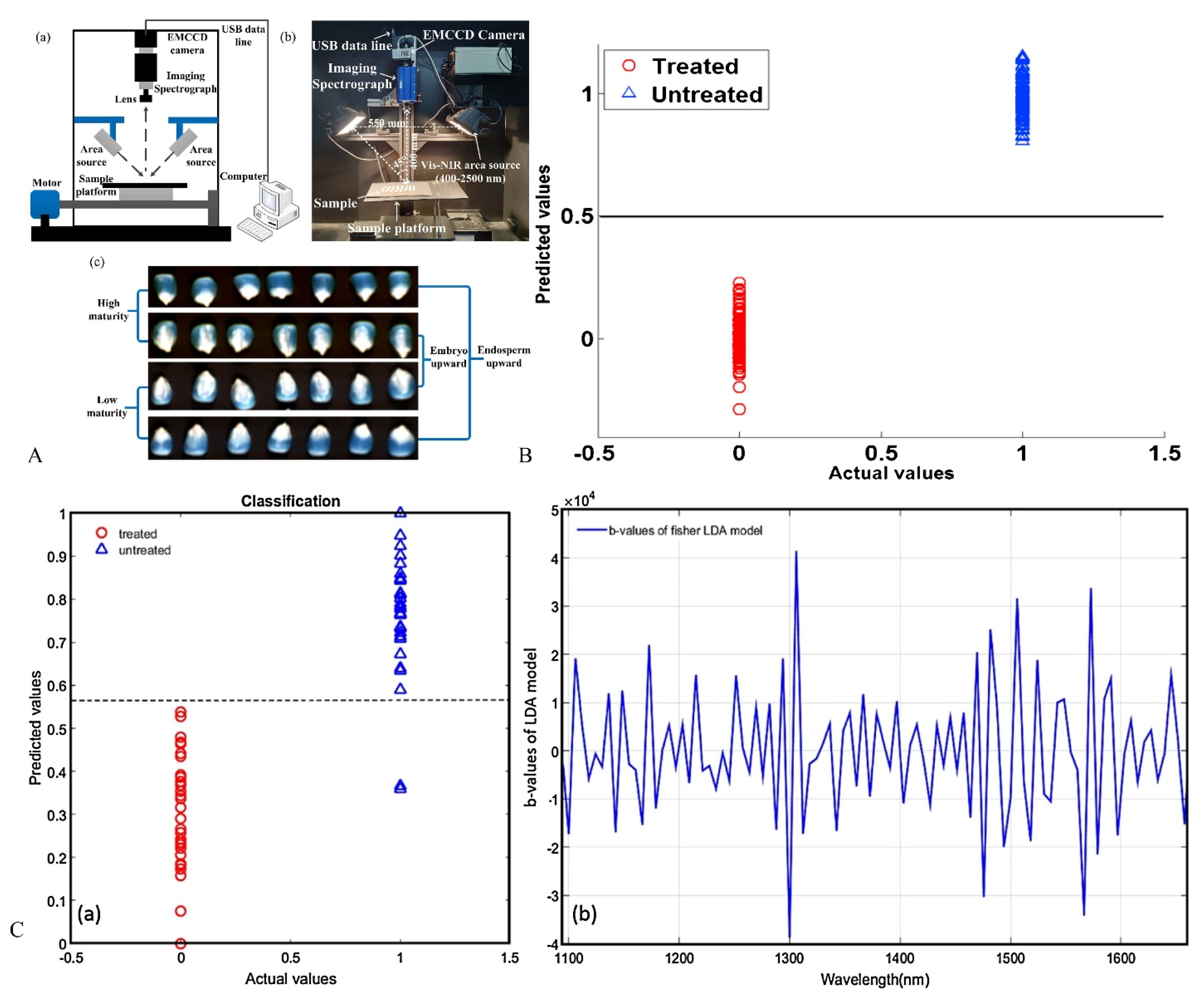
| Type of Seed | Preparation | Spectral Analysis Technology | Analysis Model | Classification Accuracy | Ref. |
|---|---|---|---|---|---|
| Maize | Microwave treatment | NIR | PLS-DA | 100% | [97] |
| Artificial aging | NIR. Vis-NIR | PLS-DA; (500–1100 nm and 1000–1800 nm) CARS-PLS-DA | 98.7% | [81] | |
| Maize (white, yellow, and purple corn) | Microwave treatment | FI-NIR. Raman | PLS-DA | 100% | [83] |
| Vis-NIR HSI. SWIR HSI | PLS-DA | 97.6% | [17] | ||
| Corn | Microwave treatment and frost treatment | NIR | PLS-DA. SIMCA. LS-SVM. K-NN | 99.0% | [96] |
| Maize | Frost treatment | NIR | PCA-OLDA or PLS-OLDA to extract spectral feature SVM, BPR, MD | 99% | [82] |
| Corn (yellow dent corn, white dent corn, and purple flint corn) | Microwave treatment | SWIR-HSI | LDA, PLS-DA, SVM | 100% | [84] |
| Maize | Artificial aging | ATR-FITR | PLS-DA | 85% | [41] |
| Maize (Huanong 101) | Artificial aging | HSI | SVM. ELM. DCNN | 100% | [101] |
| Maize (Deyu 977) | Frost damage | Vis-NIR-HSI | KNN. SVM. ELM. DCNN | 100% | [102] |
| Maize1 (101101) and Mazie2 (7879) | Artificial aging | HSI | SVM | 100% | [103] |
| Maize (Zhengdan 958) | Maturity division | NIR-HSI | PLS-DA. DT. AddBoost | 99.35% | [86] |
| Retinispora (Hinoki cypress) | Not mentioned (may be natural mold infection) | FI-NIR | PLS-DA. VIP-PLS-DA | 99.2% | [98] |
| Japanese mustard spinach (Brassica rapa var. perviridis) | Storage for one year | NIR-HSI | CNN | - | [99] |
| Wheat (Luyuan 502) | Artificial aging | Vis-NIR-HSI. SWIR-HSI | PLS-R. SPA-PLS-R. PLS-R-RC | 94.8% | [1] |
| Rice (Zhongzao39, Zhongzao 8, Y liangyou2) | Storage for 5–7 years | NIR-HSI | LR; SVM; CNN; PCA-LR; PCA-SVM; PCA-CNN | 86.67% | [85] |
| Rice (Yanfeng) | Frost damage | NIR-HSI | DT; KNN; SVM; DF | 99.93% | [87] |
| Wheat (Xinong 979, Xinong 20, Zhengmai 366, Xinmai 26, Zhoumai 26 and Chinese Spring) | Artificial aging | NIR | PCA/SPA + SVM/ELM/RF/AdaBoost | 92.2% | [9] |
| Method | Type of Seed | Preparation | Statistical Theory | Performance | Ref. |
|---|---|---|---|---|---|
| Biospeckle laser | Bean | Control moisture content. Prepare viable and nonviable seeds | COM-IM. GD; | Speckle can be used to distinguish the vigor of seeds | [110] |
| Maize | Imbibition and bisection | WE | The distribution area of maize seed vigor was found | [111] | |
| Maize | Gr | [112] | |||
| Maize | Bisection | IM | [113] | ||
| Maize and bean | Maize: Not mentioned Bean: dead and live seeds with different water content | Fuji; Entropy; IM | The distribution area of maize seed vigor was found and different manifestations of obtaining dead seeds and live seeds | [117] | |
| Coffee bean (Coffea arabica L.) | Not mentioned | STD biospeckle index | Accuracy: 87.5% | [115] | |
| Chickpea (Cicer arietinum L.) | Water treatment and chemical treatment | MSF-BA | BA and GR were positively correlated (R = 0.88, p < 0.01) and negatively correlated with MGT (r = −0.92, p < 0.01) | [109] | |
| Bean | Disinfect and dry | FTHSP + COM + AVD | BA was positively correlated with the initial water content of seeds (0.97–0.99) and related to seed germination. | [116] | |
| Soybean [Glycine max (L.) Merrill] | Imbibition | CNN; TL (VGG-16, VGG-19, InceptionV3, ResNet50) | Accuracy: 98.31% | [114] | |
| X-ray CT imaging | Moringa peregrina, Abruspreca torius, Acacia tortilis, Acacia ehrenbergiana and Arthrocnemum macrostachyum | No special treatment | - | High vigor seeds have clear morphology Low-vigor seeds have morphology damage | [118] |
| X-ray | Muskmelon seed | LDA | Accuracy: 98.9% | [119] | |
| X-ray CT imaging | Crambe abyssinica | CNN | Accuracy: 95% | [19] | |
| Biosensor (MCLA) | Pepper | Natural aging | PLS-DA | Accuracy: 88.7% | [120] |
| EIS | Rice (Jing Dao No. 21), Maize (Tai Gu No. 1/2), Wheat (Ze Yu No. 2) | Artificial aging | Chemiluminescence induced by MCLA | Positively correlated with TTC staining (r = 0.99) | [121] |
| Volatile metabolites (ethanol) | Rice | Natural aging | FLD | Accuracy: 90% | [20] |
| GC-IMS | Melon seed | - | Significant correlation with other germination indexes | [122] | |
| X-ray CT imaging | Sweet corn | Natural aging and Artificial aging | GC-IMS-AS-PCC-VIP-PLS-R | Accuracy: 94.7% | [40] |
| IDS | Juniperus polycarpos seed | No special treatment | - | Germination rate has improved | [123] |
| IDS | Pinus patula seeds | [124] | |||
| IDS | Schinus molle L. seeds | [125] | |||
| Atmospheric Pressure Plasma Treatment | Capsicum annuum L. and Trichosanthes cucumerina seeds. | Stored at 25 °C for one year | - | Germination rate has improved | [126] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xing, M.; Long, Y.; Wang, Q.; Tian, X.; Fan, S.; Zhang, C.; Huang, W. Physiological Alterations and Nondestructive Test Methods of Crop Seed Vigor: A Comprehensive Review. Agriculture 2023, 13, 527. https://doi.org/10.3390/agriculture13030527
Xing M, Long Y, Wang Q, Tian X, Fan S, Zhang C, Huang W. Physiological Alterations and Nondestructive Test Methods of Crop Seed Vigor: A Comprehensive Review. Agriculture. 2023; 13(3):527. https://doi.org/10.3390/agriculture13030527
Chicago/Turabian StyleXing, Muye, Yuan Long, Qingyan Wang, Xi Tian, Shuxiang Fan, Chi Zhang, and Wenqian Huang. 2023. "Physiological Alterations and Nondestructive Test Methods of Crop Seed Vigor: A Comprehensive Review" Agriculture 13, no. 3: 527. https://doi.org/10.3390/agriculture13030527
APA StyleXing, M., Long, Y., Wang, Q., Tian, X., Fan, S., Zhang, C., & Huang, W. (2023). Physiological Alterations and Nondestructive Test Methods of Crop Seed Vigor: A Comprehensive Review. Agriculture, 13(3), 527. https://doi.org/10.3390/agriculture13030527