
Effect of Zilpaterol Hydrochloride and Zinc Methionine on Growth, Carcass Traits, Meat Quality, Fatty Acid Profile and Gene Expression in Longissimus dorsi Muscle of Sheep in Intensive Fattening

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Treatments and Experimental Conditions
2.2. Carcass Traits
2.3. Meat Quality Assessment
Nutritional Composition, Tenderness, Water Retention Capacity and Maturation (pH and Color)
2.4. Analysis of Fatty Acids
2.5. Total RNA Extraction and Reverse Transcription
2.6. Primers and Real-Time PCR Quantification
2.7. Gene Expression Analysis
2.8. Statistical Analysis
3. Results
3.1. Productive Performance
3.2. Carcass Traits
3.3. pH Values and Meat Color
3.4. Fatty Acids
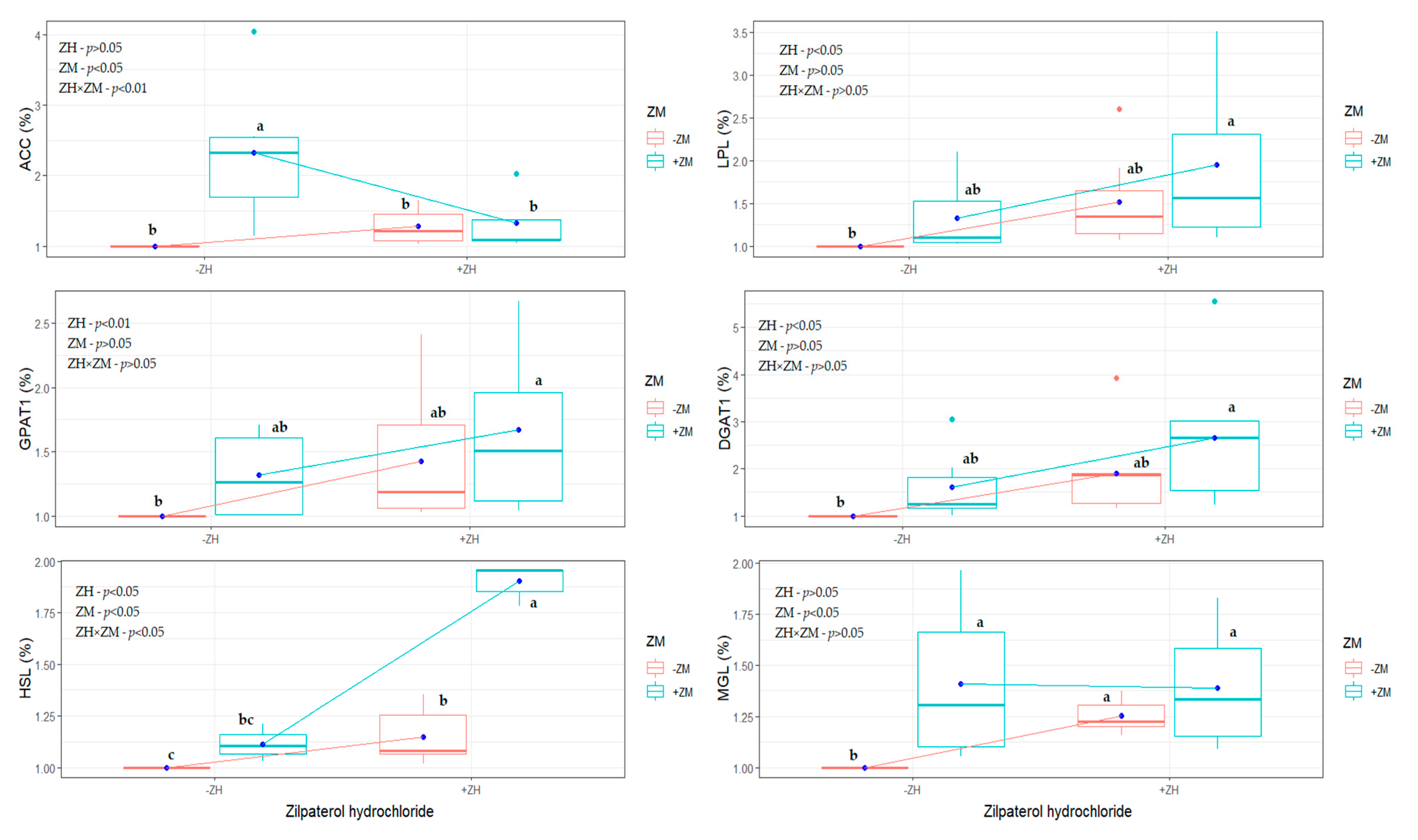
3.5. Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Testa, M.L.; Grigioni, G.; Panea, B.; Pavan, E. Color and marbling as predictors of meat quality perception of argentinian consumers. Foods 2021, 10, 1465. [Google Scholar] [CrossRef] [PubMed]
- Raza, S.H.A.; Gui, L.; Khan, R.; Schreurs, N.M.; Xiaoyu, W.; Wu, S.; Mei, C.; Wang, L.; Ma, X.; Wei, D. Association between FASN gene polymorphisms ultrasound carcass traits and intramuscular fat in Qinchuan cattle. Gene 2018, 645, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Savell, J.W.; Cross, R. The role of fat in the palatability of beef, pork and lamb. Meat Res. 1986, 1, 1–10. [Google Scholar]
- Cianzio, D.S.; Topel, D.G.; Whitehurst, G.B.; Beitz, D.C.; Self, H.L. Adipose tissue growth and cellularity: Changes in bovine adipocyte size and number. J. Anim. Sci. 1985, 60, 970–976. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Hopkins, D.L.; Butler, K.L.; Dunshea, F.R.; Warne, R.D. Genotype and age effects on sheep meat production 2. Carcass quality traits. Aust. J. Exp. Agric. 2007, 47, 1147–1154. [Google Scholar] [CrossRef]
- McPhee, M.J.; Hopkins, D.L.; Pethick, D.W. Intramuscular fat levels in sheep muscle during growth. Aust. J. Exp. Agric. 2008, 48, 904–909. [Google Scholar] [CrossRef]
- Reeds, P.J.; Mersmann, H.J. Protein and energy requirements of animal treated with adrenergic agonists: A discussion. J. Anim. Sci. 1991, 59, 1247–1255. [Google Scholar] [CrossRef]
- Domínguez, V.I.; Mondragón, A.J.; González, R.M.; Salazar, G.F.; Bórquez, G.J.; Aragón, A.M. Los β-agonistas adrenérgicos como modificadores metabólicos y su efecto en la producción, calidad e inocuidad de la carne de bovinos y ovinos: Una revisión. CIENCIA Ergo Sum 2009, 16, 278–284. [Google Scholar]
- Dikeman, M.E. Effect of metabolic modifiers on carcass traits and meat quality. Meat Sci. 2007, 7, 121–135. [Google Scholar] [CrossRef]
- Delmore, R.J.; Hodgen, J.M.; Johnson, B.J. Perspectives on the application of zilpaterol hydrochloride in the United States beef industry. J. Anim. Sci. 2010, 88, 2825–2828. [Google Scholar] [CrossRef]
- Beckett, J.L.; Delmore, R.J.; Duff, G.C.; Yates, D.A.; Allen, D.M.; Lawrence, T.E. Effects of zilpaterol hydrochloride on growth rates, feed conversion, and carcass traits in calf-fed Holstein steers. J. Anim. Sci. 2009, 87, 4092–4100. [Google Scholar] [CrossRef]
- Rodríguez-Gaxiola, M.; Domínguez-Vara, I.A.; Barajas-Cruz, C.B.; Mariezcurrema-Berazain, B.A.; Bórquez-Gastelum, J.L.; Cervantes-Pacheco, P.B. Effects of zilpaterol hydrochloride and zinc methionine on growth performance and carcass characteristics of beef bulls. Can. J. Anim. Sci. 2015, 95, 609–615. [Google Scholar] [CrossRef]
- Partida, P.J.A.; Casaya, A.T.; Rubio, S.M.; Méndez, D.R. Meat quality in Katahdin lamb terminal crosses treated with zilpaterol hydrochloride. J. Food Res. 2015, 4, 48–57. [Google Scholar] [CrossRef]
- Choi, C.B.; Jung, K.K.; Chung, K.Y.; Yang, B.S.; Chin, K.B.; Suh, S.W.; Oh, D.H.; Jeon, M.S.; Baek, K.H.; Lee, S.O.; et al. Administration of zilpaterol hydrochloride alters feedlot performance, carcass characteristics, muscle, and fat profiling in finishing Hanwoo steers. Livest. Sci. 2013, 157, 435–441. [Google Scholar] [CrossRef]
- Van Bibber-Krueger, C.L.; Miller, K.A.; Parsons, G.L.; Thompson, L.K.; Drouillard, J.S. Effects of zilpaterol hydrochloride on growth performance, blood metabolites, and fatty acid profiles of plasma and adipose tissue in finishing steers. J. Anim. Sci. 2015, 93, 2419–2427. [Google Scholar] [CrossRef]
- Rodríguez, G.M.A. Efecto de la Complementación Dietaria del Clorhidrato de Zilpaterol y su Interacción con Zinc y Cromo Orgánicos en la Respuesta Productiva, Características de la Canal y Calidad de la Carne de Bovinos y Ovinos en Engorda Intensiva. Ph.D. Thesis, Facultad de Medicina Veterinaria y Zootecnia, Universidad Autónoma del Estado de México, Mexico City, Mexico, 2015; p. 120. [Google Scholar]
- Dávila-Ramos, H.; Robles-Estrada, J.C. Zilpaterol hydrochloride modify the fatty acids profile of intramuscular fat of feedlot lambs. J. Anim. Sci. 2014, 92 (Suppl. S2), 1929p. [Google Scholar]
- Mondragón, A.J.; Domínguez-Vara, I.A.; Pinos-Rodríguez, J.M.; González, M.; Bórquez, J.L.; Domínguez, A.; Mejía, M.L. Effects of feed supplementation of zilpaterol hydrochloride on growth performance and carcass traits of finishing lambs. Acta Agric. Scan. Anim. Sci. 2010, 60, 47–52. [Google Scholar] [CrossRef]
- Partida, P.J.A.; Braña, V.D. Metodología para la evaluación de la canal ovina. Foll. Técnico INIFAP 2011, 9, 2–57. [Google Scholar]
- Smith, O.B.; Akinbamizo, O.O. Micronutrients and reproduction in farm animals. Anim. Reprod. Sci. 2000, 60–61, 549–560. [Google Scholar] [CrossRef]
- Vierboom, M.; Engle, T.E.; Kimberling, C.V. Effects of gestational status on apparent absorption and retention of copper and zinc in mature Angus cows and Suffolk ewes. Asian Austr. J. Anim. Sci. 2003, 16, 515–518. [Google Scholar] [CrossRef]
- Malcolm-Callis, K.J.; Duff, J.C.; Gunter, S.A.; Kegley, E.B.; Vermeire, D.A. Effects of supplemental zinc concentration and source on performance, carcass characteristics, and serum values in finishing beef steers. J. Anim. Sci. 2000, 78, 2801–2808. [Google Scholar] [CrossRef] [PubMed]
- Spears, J.W.; Kegley, E.B. Effect of zinc source (zinc oxide vs zinc proteinate) and level on performance, carcass characteristics, and immune response of growing and finishing steers. J. Anim. Sci. 2002, 80, 2747–2752. [Google Scholar] [PubMed]
- Rodríguez-Maya, M.A.; Domínguez-Vara, I.A.; Trujillo-Gutiérrez, D.; Morales-Almaráz, E.; Sánchez-Torres, J.E.; Bórquez-Gastelum, J.L.; Acosta-Dibarrat, J.; Grageola-Nuñez, F.; Rodríguez-Carpena, J.G. Growth performance parameters, carcass traits and meat quality of lambs supplemented with zinc methionine or/and zinc oxide in feedlot system. Can. J. Anim. Sci. 2019, 99, 585–595. [Google Scholar] [CrossRef]
- Dudley-Cash, W.A. Organic form of zinc may provide additional benefits in poultry. Feedstuffs 1997, 6, 11–17. [Google Scholar]
- Association of American Feed Control Officials (AAFCO). Official Publication; AAFCO: Atlanta, GA, USA, 1997. [Google Scholar]
- Miles, R.D.; Henry, P.R. Trace mineral bioavailability. Cienc. Anim. Bras. 2000, 1, 73–93. [Google Scholar]
- Díaz-Zagoya, J.; Hicks-Gómez, J.J. Bioquímica, 2nd ed.; Interamericana-McGraw-Hill: Mexico City, México, 1995; pp. 203–253. [Google Scholar]
- Kennedy, K.J.; Rains, T.M.; Shay, N.F. Zinc deficiency changes preferred macronutrient intake in subpopulations of Sprague-Dawley outbred rats and reduces hepatic pyruvate kinase gene expression. J. Nutr. 1998, 128, 43–49. [Google Scholar] [CrossRef]
- Kawasaki, E. ZnT8 and type 1 diabetes. Endocr. J. 2012, 59, 531–537. [Google Scholar] [CrossRef]
- Oh, Y.S.; Choi, C.B. Effects of zinc on lipogenesis of bovine intramuscular adipocytes. Asian Australas. J. Anim. Sci. 2004, 17, 1378–1382. [Google Scholar] [CrossRef]
- Chen, M.D.; Lin, P.Y.; Cheng, V.; Lin, V.H.L. Zinc supplementation aggravates body fat accumulation in genetically obese (ob/ob) mice and dietary-obese mice. Biol. Trace Elem. Res. 1996, 52, 125–132. [Google Scholar] [CrossRef]
- Greene, L.W.; Lunt, D.K.; Byers, F.M.; Chirase, N.K.; Richmond, C.E.; Knutson, R.E.; Schelling, G.T. Performance and carcass quality of steers supplemented with zinc oxide or zinc methionine. J. Anim. Sci. 1988, 66, 1818–1823. [Google Scholar] [CrossRef]
- Faustman, C.; Sun, Q.; Mancini, R.; Suman, S.O.P. Myoglobin and lipid oxidation interactions: Mechanistic bases and control. Meat Sci. 2010, 86, 86–94. [Google Scholar] [CrossRef] [PubMed]
- NRC (National Research Council). Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids, and New World Camelids; The National Academies Press: Washington, DC, USA, 2007; p. 384. [Google Scholar]
- NAS (National Academic of Science, Engineering, and Medicine). Nutrient Requirements of Beef Cattle, 8th ed.; The National Academic Press: Washington, DC, USA, 2016; 475p. [Google Scholar]
- Huang, L.; Tepaamorndech, S.; Kirschke, C.P.; Newman, J.W.; Keyes, W.R.; Pedersen, T.L.; Dum, J. Aberrant fatty acid metabolism in skeletal muscle contributes to insulin resistance in zinc transporter 7 (Znt7)-knockout mice. J. Biol. Chem. 2018, 293, 7549–7563. [Google Scholar] [CrossRef] [PubMed]
- NOM-062-ZOO-1999. Especificaciones Técnicas Para la Producción, Cuidado y uso de los Animales de Laboratorio. Available online: http://www.gob.mx/cms/uploads/attachment/file/203498/NOM-062-ZOO-1999_220801.pdf (accessed on 10 January 2018).
- Association of Official Analytical Chemists. Official Methods of Analysis Association of Official Analytical Chemists, 18th ed.; AOAC: Arlington, VA, USA, 2007. [Google Scholar]
- Fick, K.A.; McDowell, L.R.; Miles, P.H.; Wilkinson, N.S.; Funk, J.D.; Conrad, J.H.; Valdivia, R. Métodos de Análisis de Minerales Para Tejidos de Plantas y Animales; Universidad de Florida: Gainesville, FL, USA, 1979; p. 135. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- NOM-033-SAG/ZOO-2014. Métodos Para dar Muerte a los Animales Domésticos y Silvestres. Available online: http://www.dof.gob.mx/nota_detalle.php?codigo=5405210&fecha=26/08/2015 (accessed on 10 January 2018).
- European Community. Commission Regulation (CE) No. 1249/2008 of 10 December 2008 laying down detailed rules on the implementation of the Community scales for the classification of beef, pig and sheep carcasses and the reporting of prices thereof. Off. J. Eur. Union. 2008, L 337/3. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT/?qid=1473400865387&uri=CELEX:32008R1249 (accessed on 5 May 2022).
- NMX-FF-106 SCFI-2006. Norma Mexicana Para la Clasificación de Carne Ovina en Canal. Productos Pecuarios. Publicada en el Diario Oficial de la Federación el 21 de Agosto de 2006. Available online: https://www.dof.gob.mx/nota_detalle.php?codigo=4916518&fecha=04/07/2006#gsc.tab=0 (accessed on 22 January 2023).
- Beltrán, J.A.; Roncalés, P. Determinación de textura. In Metodología Para el Estudio de Calidad de Canal y Carne en Rumiantes; Cañeque, V., Sañudo, C., Eds.; Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria: Madrid, Spain, 2000; pp. 168–174. (In Spanish) [Google Scholar]
- Honikel, K.O. Reference methods for the assessment of physical characteristics of meat. Meat Sci. 1998, 49, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Hunt, M.C.; Acton, J.C.; Benedict, R.C.; Calkins, C.R.; Cornforth, D.P.; Jeremiah, L.E. American Meat Science Association Guidelines for Meat Colour Evaluation. In Proceedings of the 44th Annual Reciprocal Meat Conference, Manhattan, KS, USA, 9–12 July 1991. [Google Scholar]
- CIE. Recommendations on Uniform Color Spaces–Color Equations, Psychometric Color Terms. Supp nr 2 to CIE Publ nr 15 (E-1.3.L) 1971 (9TC-1-3); CIE: Paris, France, 1978. [Google Scholar]
- American Meat Science Association. Meat Color Measurement Guidelines; American Meat Science Association: Champaign, IL, USA, 2012; p. 125. [Google Scholar]
- Enser, M.; Hallett, K.G.; Hewett, B.; Fursey, G.A.J.; Wood, O.J.D.; Harrington, G. Fatty acid content and composition of UK beef and lamb muscle in relation to production system and implications for human nutition. Meat Sci. 1998, 49, 329–341. [Google Scholar] [CrossRef]
- Ulbricht, T.L.; Southgate, D.A.T. Coronary heart disease: Seven dietary factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef]
- Jeong, J.; Kwon, E.G.; Im, S.K.; Seo, K.S.; Baik, M. Expression of fat deposition and fat removal genes is associated with intramuscular fat content in Longissimus dorsi muscle of Korean cattle steers. J. Anim. Sci. 2012, 90, 2044–2053. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS User’s Guide: Statistics Version 9.1.3; SAS Institute Inc.: Cary, NC, USA, 2006. [Google Scholar]
- Salinas, C.J.; Domínguez, M.M.; Díaz, M.R.; Cruz, B.P.; Montaño, G.M.F.; Arzola, A.C. Effect of duration of zilpaterol hidrochloride treatment on carcass characteristics and weight gain in grazing Pelibuey lambs. J. Appl. Anim. Res. 2006, 29, 25–28. [Google Scholar] [CrossRef]
- Dávila-Ramírez, J.L.; Avendaño-Reyes, L.; Macías-Cruz, U.; Peña-Ramos, E.A.; Islava-Lagarda, T.Y.; Zamorano-García, L. Fatty acid composition and physicochemical and sensory characteristics of meat from ewe lambs supplemented with zilpaterol hydrochloride and soybean oil. Anim. Prod. Sci. 2016, 57, 767–777. [Google Scholar] [CrossRef]
- Mersmann, H.J. Overview of the effects of beta-adrenergic receptor agonists on animal growth including mechanisms of action. J. Anim. Sci. 1998, 76, 160–172. [Google Scholar] [CrossRef]
- López, C.M.A.; Ramírez, R.G.; Aguilera, S.J.; Aréchiga, C.F.; Méndez, L.F.; Rodríguez, H.; Silva, J.M. Effect of ractopamine hydrochloride and zilpaterol hydrochloride on growth, diet digestibility, intake and carcass characteristics of feedlot lambs. Livest. Sci. 2010, 131, 23–30. [Google Scholar] [CrossRef]
- Li, Y.Z.; Cristopherson, B.T.; Ly, T.; Moibi, J.A. Effects of a beta-adrenergic Agonist (L-644,969) on performance and carcass traits of growing lambs in a cold environment. Can. J. Anim. Sci. 2000, 80, 459–465. [Google Scholar] [CrossRef]
- Miller, M.F.; García, D.K.; Coleman, M.E.; Ekeren, P.A.; Lunt, D.K.; Wagrner, K.A.; Procknor, M.; Welsh, T.H.; Smith, S.B. Adipose tissue, Longissimus muscle and anterior pituitary growth and function in clenbuterol-fed heifers. J. Anim. Sci. 1988, 66, 12–20. [Google Scholar] [CrossRef]
- Kubik, R.H.; Tietze, S.M.; Schmidt, T.B.; Yates, D.T.; Petersen, J.L. Investigation of the skeletal muscle trascriptome in lambs fed β adrenergic agonist and subjected to heat stress for 21 d. Transl. Anim. Sci. 2018, 2, S53–S56. [Google Scholar] [CrossRef]
- Koohmaraie, M. Biochemical factors regulating the toughening and tenderization processes of meat. Meat Sci. 1996, 43, 193–201. [Google Scholar] [CrossRef]
- Van Hoof, N.; Schilt, R.; Van der Vlis, E.; Boshuis, P.; Van Baak, M.; Draaijer, A.; De Wasch, K.; Van de Wiele, M.; Van Hende, J.; Courtheyn, D.; et al. Detection of zilpaterol (Zilmax®) in calf urine and faeces with liquid chromatography-tandem mass spectrometry. Anal. Chem. Acta 2005, 529, 189–197. [Google Scholar] [CrossRef]
- Brand, T.S.; Genis, M.P.; Hoffman, L.C.; van de VyVER, W.F.J.; Swart, R.; Jordaan, G.F. The effect of dietary energy and the inclusión of a β-adrenergic agonist in the diet on the meat quality of feedlot lambs. S. Afr. J. Anim. Sci. 2013, 43, S141–S145. [Google Scholar] [CrossRef]
- McBeth, L.J.; Stein, D.R.; Pillai, A.T.V.; Hersom, M.J.; Krehbiel, C.R.; De Silva, U.; Geisert, R.D.; Malayer, J.R.; Morgan, J.B.; Larson, C.K.; et al. Effect of zinc source and level on finishing cattle performance, carcass characteristics, and adipocyte differentiation. Okla. Agric. Exp. Stn. Res. 2005, 4, 1–9. [Google Scholar]
- Park, K.S.; Lee, N.D.; Lee, K.H.; Seo, J.T.; Choi, K.Y. The ERK pathway involves positive and negative regulations of HT-29 colorectal cancer cell growth by extracellular zinc. Am. J. Physiol.-Gastrointest. Liver Physiol. 2003, 285, 1181–1188. [Google Scholar] [CrossRef]
- Chen, M.D.; Liou, S.J.; Lin, P.Y.; Yang, V.C.; Alexander, P.S.; Lin, W.H. Effects of zinc supplementation on the plasma glucose level and insulin activity in genetically obese (ob/ob) mice. Biol. Trace Elem. Res. 1998, 61, 303–311. [Google Scholar] [CrossRef]
- Jobgen, W.S.; Fried, S.K.; Fu, W.J.; Meininger, C.J.; Wu, G. Regulatory role for the arginine–nitric oxide pathway in metabolism of energy substrates. J. Nutr. Biochem. 2006, 17, 571–588. [Google Scholar] [CrossRef] [PubMed]
- Partida-De la Peña, J.A.; Casaya-Rodríguez, T.A.; Rubio-Lozano, M.S.; Méndez-Medina, R.D. Efecto del clorhidrato de zilpaterol sobre las características de la canal en cruzas terminales de corderos Kathadin. Vet. México 2015, 2, 1–13. [Google Scholar]
- Dávila, R.J.L.; Avendaño, R.L.; Macías, C.U.; Torrentera, O.G.; Zamorano, G.L.; Peña, R.A.; González, R.H. Effects of zilpaterol hydrochloride and soybean oil supplementation on physicochemical and sensory characteristics of meat from hair lambs. Small Rum. Res. 2013, 114, 253–257. [Google Scholar] [CrossRef]
- Moloney, A.P.; Beermann, D.H. Mechanisms by Which Beta-Adrenergic Agonists Alter Growth and Body Composition in Ruminants. Residues of Veterinary Drugs and Mycotoxin in Animal Production. New Methods for Risk Assessment and Residues Control; Wagening: Pers, The Netherlands, 1996; pp. 124–136. [Google Scholar]
- Garrido, M.D.; Bañon, S.; Álvarez, D. Medida del pH. In Estandarización de las Metodologías Para Evaluar la Calidad del Producto (Animal Vivo, Canal, Carne y Grasa) en Los Rumiantes; Cañeque, V., y Sañudo, C., Eds.; Serie ganadera No. 3; Monografías INIA: Madrid, España, 2005. [Google Scholar]
- Khliji, S.; van de Ven, R.J.; Lamb, T.A.; Lanza, M.; Hopkins, D.L. Relationship between consumer ranking of lamb colour and objective measures of colour. Meat Sci. 2010, 85, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.B.; Lunt, D.K.; Chung, K.Y.; Choi, C.B.; Tume, R.K.; Zembayashi, M. Adiposity, fatty acid composition, and delta-9 desaturase activity during growth in beef cattle. Anim. Sci. J. 2006, 77, 478–486. [Google Scholar] [CrossRef]
- Fritsche, S.; Rumsey, T.S.; Yurawecz, M.P.; Ku, Y.; Fritscher, J. Influence of growth promoting implants on fatty acid composition including conjugated linoleico acid isomers in beef fat. Eur. Food Res. Technol. 2001, 212, 621–629. [Google Scholar] [CrossRef]
- Ibrahim, R.M.; Marchelo, J.A.; Duff, G.C. Effects of implanting beef steers with zeranol on fatty acid composition of subcutaneous and intramuscular fat. Prof. Anim. Sci. 2006, 22, 301–306. [Google Scholar] [CrossRef]
- McKeith, F.K.; Perkins, E.G.; Bechtel, P.J.; Jones, D.J.; Mowery, D.H. Effects of ractopamine on the fatty acid composition and cholesterol content of pork Longissimus muscle. J. Anim. Sci. 1990, 68, 336. [Google Scholar]
- Engeseth, N.J.; Lee, K.O.; Bergen, W.G.; Helferich, W.G.; Knudson, B.K.; Merkel, R.A. Fatty acid profiles of lipid depots and cholesterol concentration in muscle tissue of finishing pigs fed ractopamine. J. Food Sci. 1992, 57, 1060–1062. [Google Scholar] [CrossRef]
- Carr, S.N.; Rincker, P.J.; Killefer, J.; Baker, D.H.; Ellis, M.; McKeith, F.K. Effects of different cereal grains and ractopamine hydrochloride on performance, carcass characteristics, and fat quality in late-finishing pigs. J. Anim. Sci. 2005, 83, 223–230. [Google Scholar] [CrossRef]
- Spurlock, M.E.; Cusumano, J.C.; Ji, S.Q.; Anderson, D.B.; Smith II, C.K.; Hancock, D.L.; Mills, S.E. The effect of ractopamine on β-adrenoceptor density and affinity in porcine adipose and skeletal muscle tissue. J. Anim. Sci. 1994, 72, 75–80. [Google Scholar] [CrossRef]
- Mills, S.; Mersmann, H.J. Beta-Adrenergic Agonists, Their Receptors, and Growth: Special Reference to the Peculiarities in Pigs; Smith, S.B., Smith, D.R., Eds.; The Biology of Fat in Meat Animals: Current Advances; American Society of Animal Science: Champaign, IL, USA, 1995; pp. 1–34. [Google Scholar]
- Adeola, O.; McBride, B.W.; Young, L.G. Metabolic responses induced by isoproterenol in ractopamine-fed pigs. J. Nutr. 1992, 122, 1280–1286. [Google Scholar] [CrossRef]
- Eisemann, J.H.; Huntington, G.B.; Ferrell, C.L. Effects of dietary clenbuterol on metabolism of the hindquarters in steers. J. Anim. Sci. 1988, 66, 342–353. [Google Scholar] [CrossRef]
- Beermann, D.H.; Butler, W.R.; Hogue, D.E.; Fishell, V.K.; Dalrymple, R.H.; Ricks, C.A.; Scanes, C.G. Cimaterol-induced muscle hypertrophy and altered endocrine status in lambs. J. Anim. Sci. 1987, 65, 1514–1524. [Google Scholar] [CrossRef]
- Santos-Silva, J.; Mendes, I.; Portugal, P.; Bessa, R. Effect of particle size and soybean oil supplementation on growth performance, carcass and meat quality and fatty acid composition of intramuscular lipids of lambs. Livest. Prod. Sci. 2004, 90, 79–88. [Google Scholar] [CrossRef]
- Moloney, A.P.; Mooney, M.T.; Kerry, J.P.; Troy, D.J. Producing tender and flavoursome beef with enhanced nutritional characteristics. Proc. Nutr. Soc. 2001, 60, 221–229. [Google Scholar] [CrossRef]
- Clejan, S.; Castro-Magana, M.; Collipp, P.J.; Jonas, E.; Maddaiah, V.T. Effects of zinc deficiency and castration on fatty acid composition and desaturation in rats. Lipids 1982, 17, 129–135. [Google Scholar] [CrossRef]
- Cunnane, S. Evidence that adverse effects of zinc deficiency on essential fatty acid composition in rats are independent of food intake. Br. J. Nutr. 1988, 59, 273–278. [Google Scholar] [CrossRef]
- Bettger, W.J.; Reeves, P.G.; Moscatelli, E.A.; Reynolds, G.; O’Dell, B.L. Interaction of zinc and essential fatty acids in the rat. J. Nutr. 1979, 109, 480–488. [Google Scholar] [CrossRef]
- Eder, K.; Kirchgessner, M. Dietary zinc deficiency and fatty acid metabolism in rats. Nutr. Res. 1996, 16, 1179–1189. [Google Scholar] [CrossRef]
- Hovenier, R.; Kanis, E.; Verhoeven, J.A. Repeatability of taste panel tenderness scores and their relationships to objective pig meat quality traits. J. Anim. Sci. 1993, 71, 2018–2025. [Google Scholar] [CrossRef] [PubMed]
- Hausman, G.J.; Dodson, M.V.; Ajuwon, K.; Azain, M.; Barnes, K.M.; Guan, L.L.; Jiang, Z.; Poulos, S.P.; Sainz, R.D.; Smith, S.; et al. Board-invited review: The biology and regulation of preadipocytes and adipocytes in meat animals. J. Anim. Sci. 2009, 87, 1218–1246. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, M.; Arias-Darraz, L.; Cárcamo, J. Effect of breed on transcriptional and protein expression of lipogenic enzymes in tail and subcutaneous adipose tissue from two grazing breeds of lambs. Animals 2019, 9, 64. [Google Scholar] [CrossRef] [PubMed]
- Browning, J.D.; Horton, J.D. Molecular mediators of hepatic steatosis and liver injury. J. Clin. Investig. 2004, 114, 147–152. [Google Scholar] [CrossRef]
- Mwangi, F.W.; Pewan, S.B.; Otto, J.R.; Adegboye, O.A.; Charmley, E.; Gardiner, C.P.; Malau-Aduli, B.S.; Kinobe, R.T.; Malau-Aduli, A.E.O. Single nucleotide polymorphisms in the fatty acid binding protein 4, fatty acid synthase and stearoyl-CoA desaturase genes influence carcass characteristics of tropical crossbred beef steers. Agriculture 2022, 12, 1171. [Google Scholar] [CrossRef]
- Kneeland, J.; Li, C.; Basarab, J.; Snelling, W.M.; Benkel, B.; Murdoch, B.; Hansen, C.; Moore, S.S. Identification and fine mapping of quantitative trait loci for growth traits on bovine chromosomes 2, 6, 14, 19, 21, and 23 within one commercial line of Bos taurus. J. Anim. Sci. 2004, 82, 3405–3414. [Google Scholar] [CrossRef]
- Rempel, L.A.; Casas, E.; Shackelford, S.D.; Wheeler, T.L. Relationship of polymorphisms within metabolic genes and carcass traits in crossbred beef cattle. J. Anim. Sci. 2012, 90, 1311–1316. [Google Scholar] [CrossRef]
- Wang, H.; Eckel, R.H. Lipoprotein lipase: From gene to obesity. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E271–E288. [Google Scholar] [CrossRef]
- Farese, R.V., Jr.; Yost, T.J.; Eckel, R.H. Tissue-specific regulation of lipoprotein lipase activity by inslulin/glucose in normal-weight humans. Metabol. Clin. Exp. 1991, 40, 214–216. [Google Scholar] [CrossRef]
- Weinstock, P.H.; Levak-Frank, S.; Hudgins, L.C.; Radner, H.; Friedman, J.M.; Zechner, R.; Breslow, J.L. Lipoprotein lipase controls fatty acid entry into adipose tissue, but fat mass is preserved by endogenous synthesis in mice deficient in adipose tissue lipoprotein lipase. Proc. Natl. Acad. Sci. USA 1997, 94, 1026–10266. [Google Scholar] [CrossRef]
- Newsom, S.A.; Schenk, S.; Li, M.; Everett, A.C.; Horowitz, J.F. High fatty acid availability after exercise alters the regulation of muscle lipid metabolism. Metab. Clin. Exp. 2011, 60, 852–859. [Google Scholar] [CrossRef]
- Bitou, N.; Ninomiya, M.; Tsujita, T.; Okuda, H. Screening of lipase inhibitors from marine algae. Lipids 1999, 34, 441–445. [Google Scholar] [CrossRef]
- Buhman, K.K.; Chen, H.C.; Farese, R.V., Jr. The enzymes of neutral lipid synthesis. J. Biol. Chem. 2001, 276, 40369–40372. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, Y.; Chen, N.; Shi, X.; Tsang, B.; Yu, Y.H. Upregulation of myocellular DGAT1 augments triglyceride synthesis in skeletal muscle and protects against fat-induced insulin resistance. J. Clin. Investig. 2007, 117, 1679–1689. [Google Scholar] [CrossRef]
- Zechner, R.; Kienesberger, P.C.; Haemmerle, G.; Zimmermann, R.; Lass, A. Adipose triglyceride lipase and the lipolytic catabolism of cellular fat stores. J. Lipid Res. 2009, 50, 3–21. [Google Scholar] [CrossRef]
- Kazala, E.C.; Petrak, J.L.; Lozeman, F.J.; Mir, P.S.; Laroche, A.; Deng, J.; Weselake, R.J. Hormone-sensitive lipase activity in relation to fat content of muscle in Wagyu hybrid cattle. Livest. Prod. Sci. 2003, 79, 87–96. [Google Scholar] [CrossRef]


{kind=link}
| Composition | (g kg−1 DM) |
|---|---|
| Milled corn | 600.00 |
| Corn stover | 160.00 |
| Soybean meal | 140.00 |
| Wheat bran | 60.00 |
| Sodium bicarbonate | 15.00 |
| Pre-mixed vitamins and minerals 1 | 25.00 |
| Chemical composition | |
| Dry matter (g kg−1) 2 | 876.80 |
| Crude protein (g kg−1 DM) 2 | 140.80 |
| Neutral detergent fiber (g kg−1 DM) 2 | 214.60 |
| Metabolizable energy (MJ kg−1 DM) 3 | 11.83 |
| Net energy for gain (MJ kg−1 DM) 3 | 5.32 |
| Calcium (g kg−1 DM) 4 | 7.50 |
| Phosphorus (g kg−1 DM) 5 | 3.62 |
| Zinc (mg kg−1 DM) 4 | 18.03 |
| Gene/Symbol | Gene Bank ID | 5′→3′ | Sequence | Amplicon Size, bp |
|---|---|---|---|---|
| Acetyl-coenzyme A carboxylase alpha, ACC | NM_174224 | Forward Reverse | aggagggaagggaatcagaa gcttgaacctgtcggaagag | 69 |
| Lipoprotein lipase, LPL | NM_001075120 | Forward | acttgccacctcattcctg | 119 |
| Reverse | acccaactctcatacattcctg | |||
| Glycerol-3-phosphate Acyltransferase 1, GPAT1 | NM_001012282 | Forward Reverse | tgtgctatctgctctccaatg ctccgccactataagaatg | 116 |
| Diacylglycerol acyltransferase 1, DGAT1 | NM_174693 | Forward Reverse | tcttccactcctgcctgaac agtaggtgatggactcggag | 96 |
| Hormone-sensitive lipase, HSL | NM_001080220 | Forward | gatgagagggtaattgccg | 100 |
| Reverse | ggatggcaggtgtgaact | |||
| Noglyceride lipase, MGL | NM_001163689 | Forward | cacttttcaaggtcttcgctg | 110 |
| Reverse | gatgtccacctccgtcttattc | |||
| Ribosomal protein S9, RPS9 1 | NM_001101152 | Forward | cctcgaccaagagctgaag | 64 |
| Reverse | cctccagacctcacgttgttc |
| Variables | Treatments | 1 SEM | Effect | |||||
|---|---|---|---|---|---|---|---|---|
| −ZH | +ZH | ZH | ZM | ZH × ZM | ||||
| −ZM | +ZM | −ZM | +ZM | |||||
| ILW (kg) 2 | 25.375 | 25.150 | 25.400 | 25.150 | 2.053 | ------- | ------- | ------- |
| FLW (kg) 3 | 44.300 | 44.100 | 44.975 | 45.500 | 1.395 | ns | ns | ns |
| TGW (kg) 4 | 18.925 | 18.950 | 19.575 | 20.375 | 1.548 | ns | ns | ns |
| DMI (kg d−1) 5 | 1.436 | 1.308 | 1.349 | 1.331 | 0.102 | ns | ns | ns |
| DWG (kg d−1) 6 | 0.253 | 0.255 | 0.264 | 0.275 | 0.021 | ns | ns | ns |
| FC (kg) 7 | 5.676 | 5.131 | 5.109 | 4.839 | 0.287 | 0.076 ᴌ | ns | ns |
| Variables | Treatments | 1 SEM | Effect | |||||
|---|---|---|---|---|---|---|---|---|
| −ZH | +ZH | ZH | ZM | ZH × ZM | ||||
| −ZM | +ZM | −ZM | +ZM | |||||
| LWS (kg) 2 | 42.000 | 40.925 | 42.425 | 42.687 | 0.975 | ns | ns | ns |
| HCW (kg) 3 | 19.687 | 20.587 | 21.675 | 22.212 | 0.860 | 0.074 ᴌ | ns | ns |
| CCW (kg) 4 | 18.675 | 18.900 | 20.075 | 20.800 | 1.418 | 0.065 ᴌ | ns | ns |
| HCY (%) 5 | 46.762 | 50.287 | 51.075 | 51.887 | 1.303 | 0.046 * | ns | ns |
| CCY (%) 6 | 44.275 | 46.175 | 47.287 | 48.700 | 0.490 | 0.042 * | ns | ns |
| LC (cm) 7 | 62.440 | 64.370 | 62.630 | 62.750 | 1.965 | ns | ns | ns |
| CI 8 | 0.298 | 0.293 | 0.317 | 0.331 | 0.011 | 0.024 * | ns | ns |
| CMUE (1–5) 9 | 3.100 | 3.100 | 3.500 | 3.600 | 0.230 | 0.065 ᴌ | ns | ns |
| Greasing (1–5) 10 | 3.000 | 3.100 | 2.700 | 2.500 | 0.110 | 0.014 * | ns | ns |
| Chop area (cm2) | 12.30 | 15.400 | 17.400 | 16.800 | 1.310 | 0.020 * | ns | ns |
| Back fat (mm) | 2.000 | 3.100 | 2.200 | 2.200 | 0.970 | ns | ns | ns |
| Kidney fat (1–4) 11 | 1.800 | 2.000 | 1.800 | 1.800 | 0.960 | ns | ns | ns |
| Dry matter (g/100 g) | 28.800 | 30.500 | 29.400 | 29.600 | 0.690 | ns | ns | ns |
| Ashes (g/100 g) | 1.460 | 1.550 | 1.660 | 1.520 | 0.210 | ns | ns | ns |
| Ether extract (g/100 g) | 4.400 | 6.130 | 4.090 | 6.920 | 0.690 | ns | 0.001 | 0.043 * |
| Crude protein (g/100 g) | 22.200 | 22.000 | 23.700 | 23.700 | 0.820 | 0.049 * | ns | ns |
| Water loss 24 h (g/kg−1) | 8.150 | 9.240 | 7.930 | 7.950 | 0.47 | ns | ns | ns |
| Cutting force (kg/cm2) | 7.880 | 8.390 | 10.700 | 9.850 | 0.64 | ns | ns | ns |
| Variables | Treatments | 1 SEM | Effect | |||||
|---|---|---|---|---|---|---|---|---|
| −ZH | +ZH | ZH | ZM | ZH × ZM | ||||
| −ZM | +ZM | −ZM | +ZM | |||||
| pH at 24 h | 5.62 | 5.85 | 6.13 | 6.06 | 0.637 | 0.001 ** | ns | ns |
| pH at 4 d | 5.66 | 6.60 | 6.13 | 6.00 | 0.107 | 0.001 ** | ns | ns |
| pH at 8 d | 5.71 | 6.60 | 6.18 | 6.11 | 0.114 | 0.001 ** | ns | ns |
| Color | ||||||||
| L24h | 41.1 | 39.2 | 35.4 | 35.8 | 1.878 | 0.019 * | ns | ns |
| a24h | 16.1 | 14.2 | 12.2 | 12.0 | 1.387 | 0.004 ** | ns | ns |
| b24h | 6.15 | 5.38 | 3.6 | 3.55 | 0.577 | 0.001 ** | ns | ns |
| C24h | 17.2 | 16.4 | 12.7 | 12.6 | 1.071 | 0.001 ** | ns | ns |
| H°24h | 21.5 | 18.8 | 14.2 | 16.3 | 2.452 | 0.009 ** | ns | ns |
| L4d | 39.7 | 39.6 | 33.3 | 33.5 | 1.444 | 0.001 ** | ns | ns |
| a4d | 15.2 | 13.9 | 11.3 | 11.8 | 0.982 | 0.002 ** | ns | ns |
| b4d | 9.09 | 8.99 | 5.46 | 5.30 | 0.976 | 0.001 ** | ns | ns |
| C4d | 17.8 | 16.6 | 12.6 | 13.0 | 1.190 | 0.010 ** | ns | ns |
| H°4d | 30.6 | 30.5 | 21.3 | 21.1 | 3.573 | 0.001 ** | ns | ns |
| L8d | 38.1 | 38.9 | 34.0 | 33.0 | 1.999 | 0.010 ** | ns | ns |
| a8d | 11.9 | 11.7 | 10.6 | 10.1 | 1.703 | ns | ns | 0.091 ᴌ |
| b8d | 9.78 | 9.54 | 5.39 | 5.08 | 0.777 | 0.001 ** | ns | ns |
| C8d | 15.7 | 15.2 | 12.1 | 12.0 | 0.871 | 0.004 ** | ns | 0.082 ᴌ |
| H°8d | 39.4 | 39 | 27.6 | 27.4 | 2.389 | 0.001 ** | ns | ns |
| Treatments | Effect | |||||||
|---|---|---|---|---|---|---|---|---|
| Fatty Acids | −ZH | +ZH | 1 SEM | ZH | ZM | ZH × ZM | ||
| −ZM | +ZM | −ZM | +ZM | |||||
| C10:0, Caproic | 0.128 | 0.130 | 0.155 | 0.113 | 0.018 | ns | ns | ns |
| C12:0, Lauric | 0.130 | 0.193 | 0.203 | 0.161 | 0.034 | ns | ns | 0.050 * |
| C14:0, Myristic | 2.393 | 2.981 | 2.948 | 2.93 | 0.278 | ns | ns | ns |
| C14:1 n-5, myristoleic | 0.301 | 0.288 | 0.363 | 0.296 | 0.034 | ns | ns | ns |
| C15:0, Pentadecanoic | 0.705 | 0.553 | 0.728 | 0.736 | 0.114 | ns | ns | ns |
| C16:0, Palmitic | 25.71 | 26.26 | 25.78 | 24.80 | 0.723 | ns | ns | ns |
| C16:1, Palmitoleic | 1.235 | 1.530 | 1.601 | 1.605 | 0.095 | 0.009 ** | 0.050 * | 0.073 ᴌ |
| C17:0, Heptadecanoic | 0.886 | 0.753 | 0.803 | 0.746 | 0.079 | ns | ns | ns |
| C17:1, Heptadecenoic | 0.566 | 0.571 | 0.586 | 0.606 | 0.067 | ns | ns | ns |
| C18:0, Stearic | 18.26 | 17.01 | 17.3 | 16.72 | 1.089 | 0.048 * | ns | ns |
| C18:1n9c, Oleic | 41.56 | 41.84 | 40.72 | 41.67 | 1.291 | ns | ns | ns |
| C18:2 c9c12, Linoleic | 5.785 | 5.905 | 6.461 | 6.963 | 1.911 | ns | ns | 0.050 * |
| C18:3 c 9c12c15, Linolenic | 0.033 | 0.041 | 0.05 | 0.058 | 0.01 | ns | ns | 0.050 * |
| C20:0, Arachidic | 0.038 | 0.036 | 0.031 | 0.016 | 0.009 | 0.050 * | ns | ns |
| C20:4n6, Eicosatetraenoic | 0.986 | 1.175 | 1.613 | 1.971 | 0.312 | 0.011 * | ns | 0.049 * |
| C22:0, Behenic | 0.086 | 0.151 | 0.15 | 0.211 | 0.039 | 0.060 ᴌ | 0.061 ᴌ | ns |
| Other trans acids | 1.181 | 0.565 | 0.491 | 0.39 | 0.294 | 0.081 ᴌ | 0.101 ᴌ | ns |
| C18:1 n-7, Vaccenic acid | 1.063 | 0.42 | 0.33 | 0.24 | 0.286 | 0.060 ᴌ | 0.101 ᴌ | ns |
| Conjugated linoleic acid | 0.115 | 0.143 | 0.161 | 0.17 | 0.029 | 0.045 * | 0.031 * | ns |
| Fatty Acids | Treatments | 1 SEM | Effect | |||||
|---|---|---|---|---|---|---|---|---|
| −ZH | +ZH | ZH | ZM | ZH × ZM | ||||
| −ZM | +ZM | −ZM | +ZM | |||||
| SFA 2 | 48.350 | 48.082 | 48.112 | 46.447 | 1.718 | ns | ns | 0.048 * |
| UFA 3 | 52.635 | 53.003 | 52.980 | 54.713 | 1.582 | ns | ns | 0.050 * |
| MFA 4 | 44.730 | 44.650 | 43.603 | 44.420 | 1.324 | ns | ns | ns |
| PUFA 5 | 7.067 | 7.422 | 8.462 | 9.335 | 1.185 | 0.06 | ns | 0.049 * |
| Ω-3FA | 0.033 | 0.042 | 0.050 | 0.048 | 0.009 | ns | ns | ns |
| Ω-6FA | 6.772 | 7.082 | 8.075 | 8.937 | 1.158 | 0.061 ᴌ | ns | ns |
| IAI 6 | 0.542 | 0.565 | 0.573 | 0.540 | 0.049 | ns | ns | ns |
| TRI 7 | 1.000 | 0.990 | 1.005 | 0.938 | 0.064 | ns | ns | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerrero-Bárcena, M.; Domínguez-Vara, I.A.; Morales-Almaraz, E.; Sánchez-Torres, J.E.; Bórquez-Gastelum, J.L.; Hernández-Ramírez, D.; Trujillo-Gutiérrez, D.; Rodríguez-Gaxiola, M.A.; Pinos-Rodríguez, J.M.; Velázquez-Garduño, G.; et al. Effect of Zilpaterol Hydrochloride and Zinc Methionine on Growth, Carcass Traits, Meat Quality, Fatty Acid Profile and Gene Expression in Longissimus dorsi Muscle of Sheep in Intensive Fattening. Agriculture 2023, 13, 684. https://doi.org/10.3390/agriculture13030684
Guerrero-Bárcena M, Domínguez-Vara IA, Morales-Almaraz E, Sánchez-Torres JE, Bórquez-Gastelum JL, Hernández-Ramírez D, Trujillo-Gutiérrez D, Rodríguez-Gaxiola MA, Pinos-Rodríguez JM, Velázquez-Garduño G, et al. Effect of Zilpaterol Hydrochloride and Zinc Methionine on Growth, Carcass Traits, Meat Quality, Fatty Acid Profile and Gene Expression in Longissimus dorsi Muscle of Sheep in Intensive Fattening. Agriculture. 2023; 13(3):684. https://doi.org/10.3390/agriculture13030684
Chicago/Turabian StyleGuerrero-Bárcena, Manuel, Ignacio Arturo Domínguez-Vara, Ernesto Morales-Almaraz, Juan Edrei Sánchez-Torres, José Luis Bórquez-Gastelum, Daniel Hernández-Ramírez, Daniel Trujillo-Gutiérrez, Miguel Angel. Rodríguez-Gaxiola, Juan Manuel Pinos-Rodríguez, Gisela Velázquez-Garduño, and et al. 2023. "Effect of Zilpaterol Hydrochloride and Zinc Methionine on Growth, Carcass Traits, Meat Quality, Fatty Acid Profile and Gene Expression in Longissimus dorsi Muscle of Sheep in Intensive Fattening" Agriculture 13, no. 3: 684. https://doi.org/10.3390/agriculture13030684
APA StyleGuerrero-Bárcena, M., Domínguez-Vara, I. A., Morales-Almaraz, E., Sánchez-Torres, J. E., Bórquez-Gastelum, J. L., Hernández-Ramírez, D., Trujillo-Gutiérrez, D., Rodríguez-Gaxiola, M. A., Pinos-Rodríguez, J. M., Velázquez-Garduño, G., & Grageola-Nuñez, F. (2023). Effect of Zilpaterol Hydrochloride and Zinc Methionine on Growth, Carcass Traits, Meat Quality, Fatty Acid Profile and Gene Expression in Longissimus dorsi Muscle of Sheep in Intensive Fattening. Agriculture, 13(3), 684. https://doi.org/10.3390/agriculture13030684