
Two-Sex Life Table Analysis of the Predator Arma chinensis (Hemiptera: Pentatomidae) and the Prediction of Its Ability to Suppress Populations of Scopula subpunctaria (Lepidoptera: Geometridae)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Rearing of Insects
2.2. Life Table and Predation Rate Studies
2.2.1. Life Table and Predation Rate Study of A. chinensis
2.2.2. Life Table Study of S. subpunctaria
2.3. Data Analysis
2.3.1. Two-Sex Life Table Analysis
2.3.2. Predation Rate Analysis
2.3.3. Population and Predation Projections
3. Results
3.1. Two-Sex Life Table and Predation Rate
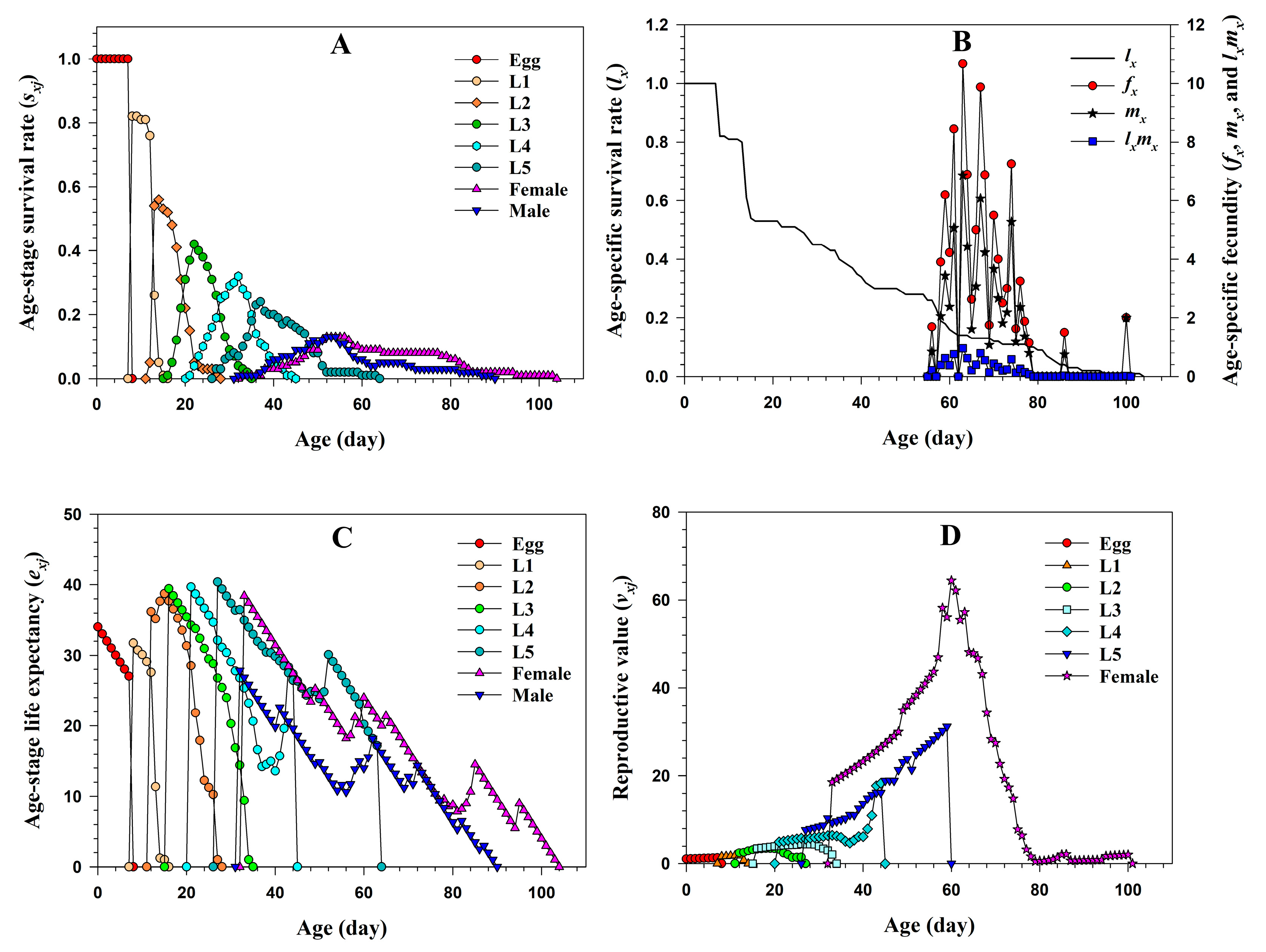
3.1.1. Two-Sex Life Table of A. chinensis Fed on Third Instar S. subpunctaria
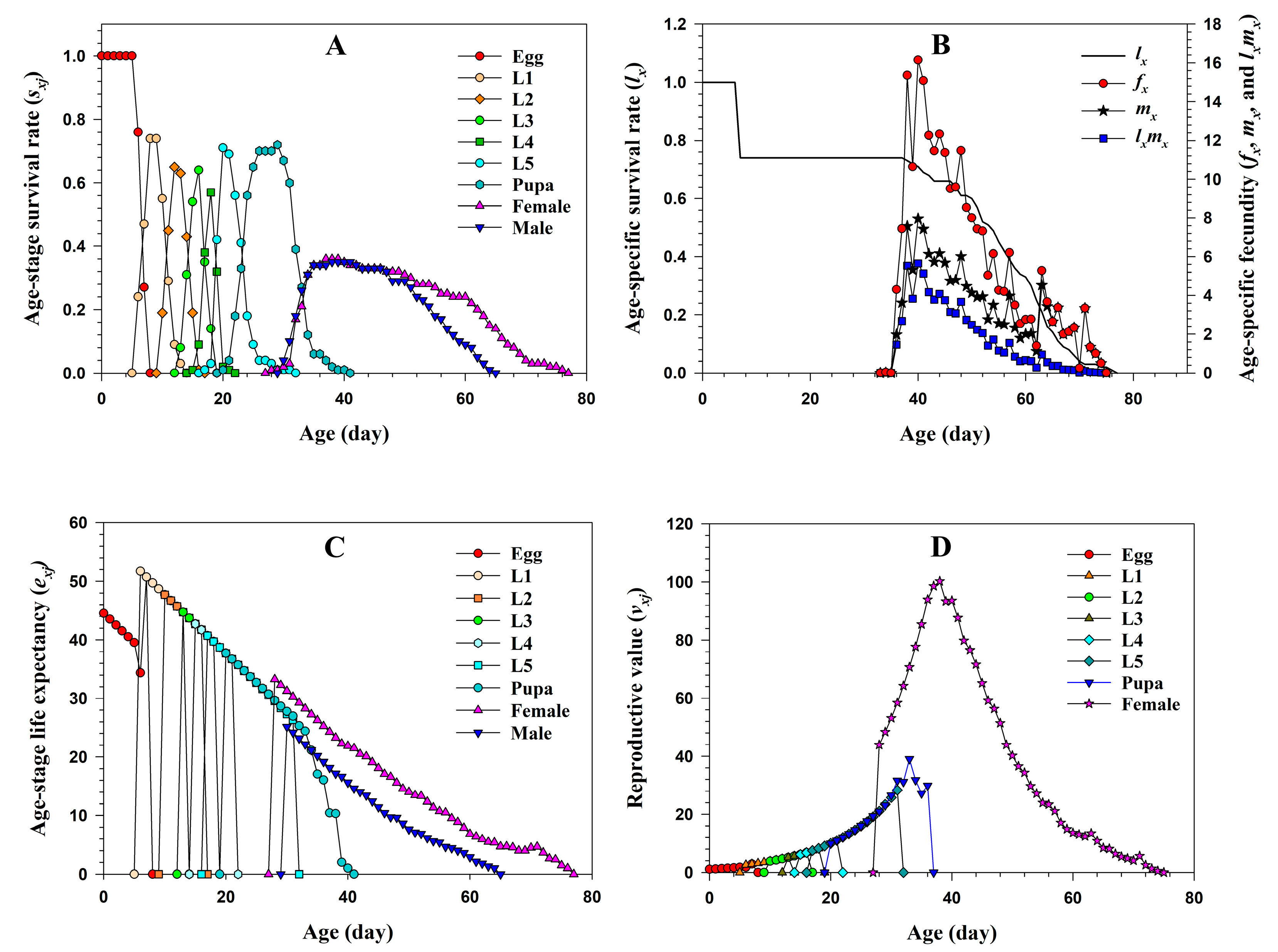
3.1.2. Two-Sex Life Table of S. subpunctaria Fed on Tea
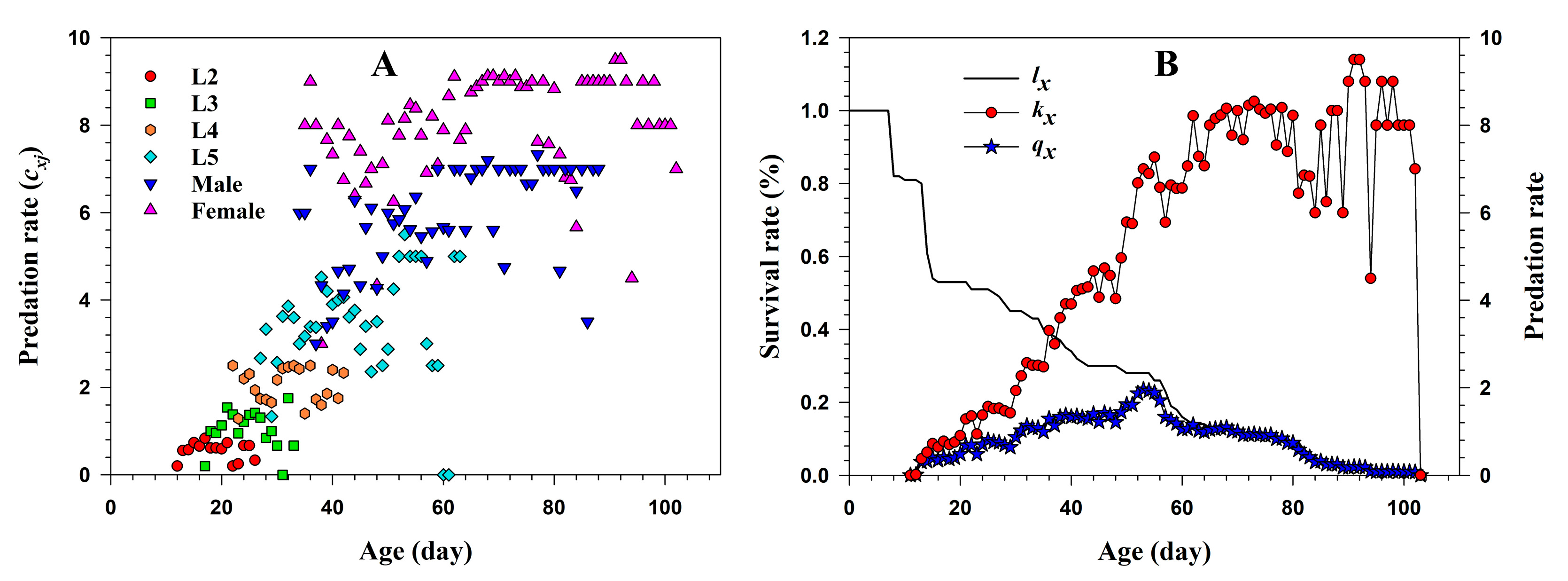
3.1.3. Predation Rate
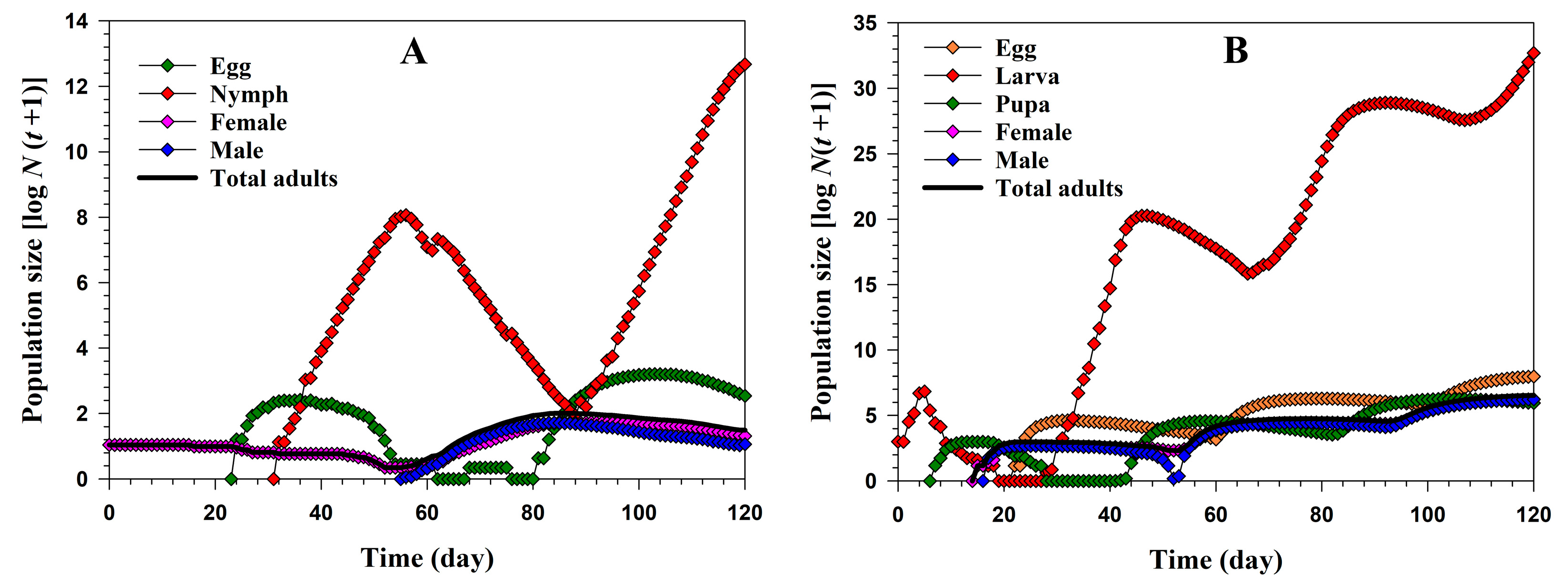
3.2. Population Projection
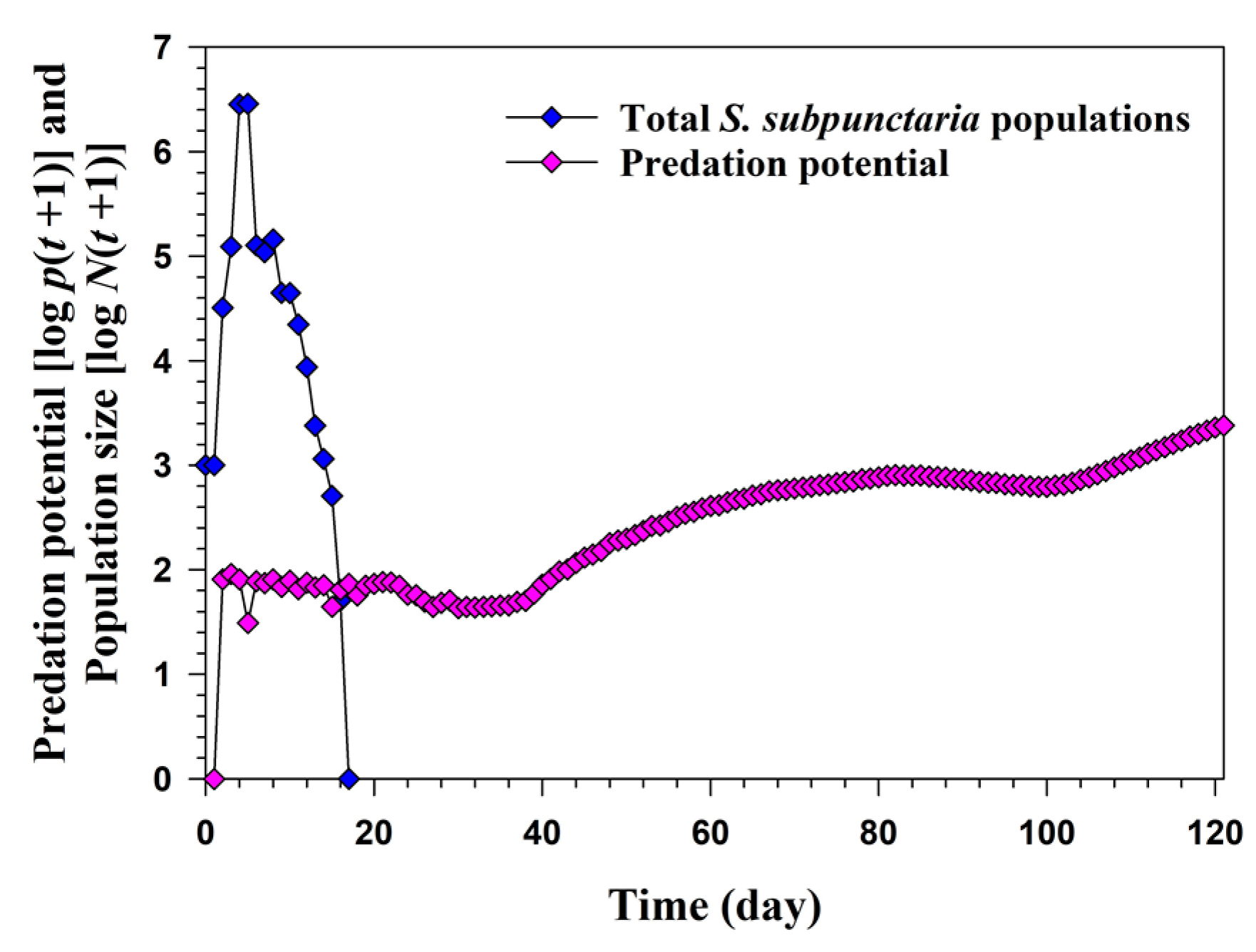
3.3. Population Growth and Predation Capacity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geng, S.; Hou, H.; Wang, G.; Jung, C.; Yin, J.; Qiao, L. Temperature-dependent oviposition model of Scopula subpunctaria (Lepidoptera: Geometridae). J. Asia-Pac. Entomol. 2021, 24, 948–953. [Google Scholar] [CrossRef]
- Qian, J.L.; Luo, Z.X.; Li, J.L.; Cai, X.M.; Bian, L.; Xiu, C.L.; Li, Z.Q.; Chen, Z.M.; Zhang, L.W. Identification of cytochrome, odorant-binding protein, and chemosensory protein genes involved in Type II sex pheromone biosynthesis and transportation in the tea pest, Scopula subpunctaria. Pestic. Biochem. Physiol. 2020, 169, 104650. [Google Scholar] [CrossRef] [PubMed]
- Preliminary report on Scopula subpunctaria research. Tea Technol. Brief. 1977, 7, 9–12.
- Hu, W.J.; Chen, W.L.; Wei, W.; Xu, X.Q. Scanning electron microscopic observation of sensilla on the antenna of male adult Scopula subpunctaria. Chin. Bull. Entomol. 2010, 47, 938–940. [Google Scholar]
- Xia, Y.S.; Li, B.; Hu, F.D.; Liu, X.; Yuan, Q.F. Preliminary research on the biological characteristics of the Scopulasub punctaria in Shandong Tea Area. China Tea 2022, 44, 55–59. [Google Scholar]
- Guo, H.W.; Luo, Z.X. Green arch-shaped insect in the tea plantation—Scopula subpunctaria. China Tea 2019, 41, 15–16. [Google Scholar]
- Ma, T.; Shi, X.; Lin, N.; Wang, Z.; Xiao, Q.; Sun, Z.; Wen, X. Temporal pattern of adult emergence and sexual behavior of Scopula subpunctaria (Lepidoptera: Geometridae). Phytoparasitica 2019, 47, 17–29. [Google Scholar] [CrossRef]
- Zhu, Y.H.; Wu, M.R. Meothrin is effective in the control of Scopula subpunctaria and Parasa lepida. Seric. Tea Newsl. 1991, 1, 40. [Google Scholar]
- Brattsten, L.; Holyoke Jr, C.; Leeper, J.; Raffa, K. Insecticide resistance: Challenge to pest management and basic research. Science 1986, 231, 1255–1260. [Google Scholar] [CrossRef] [Green Version]
- Pan, M.Z.; Fu, Z.X.; Li, Y.Y.; Chen, H.Y.; Zhang, L.S.; Liu, T.X. Role of host plants in the suitability and dispersal of an omnivorous predator Arma chinensis Fallou (Hemiptera: Pentatomidae: Asopinae) in a biological control context. J. Plant Dis. Prot. 2022, 129, 861–868. [Google Scholar] [CrossRef]
- Zheng, L.Y. Arma Hahn (Hemiptera: Stinkbugs) in China. Nat. Enemies Insect 1981, 4, 28–32. [Google Scholar]
- Liu, H.Q.; Gao, Z.A. Utilization value of Arma chinensis. Tianjin Agric. For. Sci. Technol. 1992, 2, 14–16. [Google Scholar]
- Rider, D.A.; Zheng, L.Y.; Kerzhner, I.M. Checklist and nomenclatural notes on the chinese pentatomidae (Heteroptera). II. pentatominae. Zoosyst. Ross. 2002, 11, 135–153. [Google Scholar] [CrossRef]
- Pan, M.Z.; Zhang, H.P.; Zhang, C.H.; Yi, Z.J.; Chen, H.Y. Effects of rearing density and sex ratio of adult Arma chinensis (Hemiptera: Pentatomidae) on their survival, fecundity and offspring’s suitability. Chin. J. Biol. Control 2018, 34, 52–58. [Google Scholar]
- Liao, P.; Miao, S.M.; Xu, R.N.; Liu, C.X.; Chen, G.K.; Wng, M.Q.; Mao, J.J.; Zhang, L.S.; Chen, H.Y. Evaluation of a new liquid artificial diet of Arma chinensis Fallou (Hemiptera: Pentatomidae). Chin. J. Biol. Control 2019, 35, 9–14. [Google Scholar]
- Li, X.P. Influence of Two Artificial Diets to Development and Cold Storage of Arma custos (Hemiptera: Pentatomidae); Beijing Forestry University: Beijing, China, 2020. [Google Scholar]
- Yang, C.; Mu, Y.L.; Wang, J.; Huang, C.Y.; Zhang, C.H.; Yu, H.P.; Chen, X.S. Predatory functional responses of Arma chinensis (Fallou) to the eggs and 3rd—instar larvae of two tobacco pests. Plant Prot. 2022, 48, 158–162+172. [Google Scholar]
- Xiao, G.R. Forest Insects of China; China Forestry Publishing House: Beijing, China, 1992; pp. 304–306. [Google Scholar]
- Zou, D.; Coudron, T.A.; Liu, C.; Zhang, L.; Wang, M.; Chen, H. Nutrigenomics in Arma chinensis: Transcriptome analysis of Arma chinensis fed on artificial diet and Chinese oak silk moth Antheraea pernyi pupae. PLoS ONE 2013, 8, e60881. [Google Scholar] [CrossRef] [Green Version]
- Li, J.J.; Zhang, C.H.; Yi, Z.J.; Ran, X.C.; Zhang, L.S.; Liu, C.X. Effects of three prey species on development and fecundity of the predaceous stinkbug Arma chinensis (Hemiptera: Pentatomidae). Chin. J. Biol. Control 2016, 32, 553–561. [Google Scholar]
- Wang, P.; Wang, S.J.; Ji, Y.H.; Si, Q.; Wang, Y.F.; Chen, R.Y. Functional responses of Arma chinensis preying on Pyrrhalta aenescens in laboratory. For. Pest Dis. 2021, 40, 32–36. [Google Scholar]
- Meng, J.Y.; Li, Z.M.; Dong, X.L.; Wang, H.C.; Tan, X.F.; Guo, X.G. Predation ability of Arma chinensis larvae on 3rd to 5th instar larvae of the Spodoptera frugiperda. Jiangsu Agric. Sci. 2022, 50, 1–5. [Google Scholar]
- Sun, J.J.; Wang, M.Q.; Zhang, C.H.; Tang, Y.T.; Li, X.Y.; Zhang, L.S.; Li, H. Predation by stink bug Arma custos on larvae of the Asian corn borer Ostrinia furnacalis. J. Plant Prot. 2022, 49, 1187–1193. [Google Scholar]
- Chi, H.; Fu, J.W.; You, M.S. Age-stage, two-sex life table and its application in population ecology and integrated pest management. Acta Entomol. Sin. 2019, 62, 255–262. [Google Scholar]
- Birch, L.C. The intrinsic rate of natural increase of an insect population. J. Anim. Ecol. 1948, 17, 15–26. [Google Scholar] [CrossRef]
- Southwood, T.R.E.; Henderson, P.A. Ecological Methods; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- van Lenteren, J.C.; Manzaroli, G. Evaluation and use of predators and parasitoids for biological control of pests in greenhouses. Integr. Pest Dis. Manag. Greenh. Crops 1999, 14, 183–201. [Google Scholar]
- Zang, Z.Y.; Chen, Y.M.; Xu, W.; Zang, L.S. Evidence of two-sex life table analysis supporting Anastatus japonicus, a more effective biological control agent of Caligula japonica compared with other two Anastatus species. Biological. Control 2023, 180, 105188. [Google Scholar] [CrossRef]
- Tuan, S.J.; Yang, C.M.; Chung, Y.T.; Lai, W.H.; Ding, H.Y.; Saska, P.; Peng, S.C. Comparison of demographic parameters and predation rates of Orius strigicollis (Hemiptera: Anthocoridae) fed on eggs of Tetranychus urticae (Acari: Tetranychidae) and Cadra cautella (Lepidoptera: Pyralidae). J. Econ. Entomol. 2016, 109, 1529–1538. [Google Scholar] [CrossRef]
- Zheng, X.M.; Chi, H.; Chu, D. A simplified recording method for insect life table studies:a case study based on Bemisia tabaci (Hemiptera: Aleyrodidae) data. Acta Entomol. Sin. 2016, 59, 663–668. [Google Scholar]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Zheng, Z.Y. Biology Character and Mass Pearing of Arma chinensis; China Biological Control Symposium: Beijing, China, 1995. [Google Scholar]
- Zheng, Y.X.; Su, G.L. Predatory natural enemies—Arma custos. Nat. Enemies Insects 1985, 02, 87–89. [Google Scholar]
- Wu, S.L.; Deng, W.; Cai, H.L.; Yang, J.S.; Zeng, W.A.; Zhou, Z.C.; Li, Y.Z. The occurrence period and effect of intraspecific cannibalism behavior of Arma chinensis under starvation. Chin. J. Biol. Control 2020, 36, 169–174. [Google Scholar]
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis. 2015, p. 197. Available online: http://140.120.197.173/Ecology/prod02.htm (accessed on 22 October 2022).
- Efron, B.; Tibshirani, R.J. An Introduction to the Bootstrap; CRC Press: Boca Raton, FL, USA, 1994. [Google Scholar]
- Huang, Y.B.; Chi, H. Age-stage, two-sex life tables of Bactrocera cucurbitae (Coquillett)(Diptera: Tephritidae) with a discussion on the problem of applying female age-specific life tables to insect populations. Insect Sci. 2012, 19, 263–273. [Google Scholar] [CrossRef]
- Wei, M.F.; Chi, H.; Guo, Y.F.; Li, X.W.; Zhao, L.L.; Ma, R.Y. Demography of Cacopsylla chinensis (Hemiptera: Psyllidae) reared on four cultivars of Pyrus bretschneideri (Rosales: Rosaceae) and P. communis pears with estimations of confidence intervals of specific life table statistics. J. Econ. Entomol. 2020, 113, 2343–2353. [Google Scholar] [CrossRef] [PubMed]
- Tuan, S.J.; Lee, C.C.; Chi, H. Population and damage projection of Spodoptera litura (F.) on peanuts (Arachis hypogaea L.) under different conditions using the age-stage, two-sex life table. Pest Manag. Sci. 2014, 70, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.Z.; Chi, H.; Chen, B.H. Comparison of the life tables and predation rates of Harmonia dimidiata (F.)(Coleoptera: Coccinellidae) fed on Aphis gossypii Glover (Hemiptera: Aphididae) at different temperatures. Biol. Control 2013, 64, 1–9. [Google Scholar] [CrossRef]
- Chi, H. TIMING-MSChart: A Computer Program for the Population Projection Based on Age-Stage, Two-Sex Life Table; National Chung Hsing University: Taichung, Taiwan, 2020. [Google Scholar]
- Chi, H. Timing of control based on the stage structure of pest populations: A simulation approach. J. Econ. Entomol. 1990, 83, 1143–1150. [Google Scholar] [CrossRef]
- Huang, H.W.; Chi, H.; Smith, C.L. Linking demography and consumption of Henosepilachna vigintioctopunctata (Coleoptera: Coccinellidae) fed on Solanum photeinocarpum (Solanales: Solanaceae): With a new method to project the uncertainty of population growth and consumption. J. Econ. Entomol. 2018, 111, 1–9. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Li, G.Y.; Yang, L.; Chi, H.; Chen, X.S. Demography and mass rearing of the medicinal blister beetle Epicauta impressicornis (Pic)(Coleoptera: Meloidae) at different temperatures. J. Econ. Entomol. 2018, 111, 2364–2374. [Google Scholar] [CrossRef] [PubMed]
- Kakde, V.; Vaishali, D.G. Status of vector borne disease, dengue in buldana town-a study report. Int. J. Life Sci. 2014, 2, 410–412. [Google Scholar]
- Sobhy, I.S.; Sarhan, A.A.; Shoukry, A.A.; El-Kady, G.A.; Mandour, N.S.; Reitz, S.R. Development, consumption rates and reproductive biology of Orius albidipennis reared on various prey. BioControl 2010, 55, 753–765. [Google Scholar] [CrossRef]
- Schoener, T.W. Dynamics of predation: The dynamics of arthropod predator-prey systems. Science 1979, 204, 491–492. [Google Scholar] [CrossRef]
- Lan, R.; Ren, X.; Cao, K.; Zhou, X.; Jin, L. Demographic evaluation of the control potential of Orius minutus (Hemiptera: Anthocoridae) preying on Dendrothrips minowai Priesner (Thysanoptera: Thripidae) at different temperatures. Insects 2022, 13, 1158. [Google Scholar] [CrossRef]
- He, Z.; Guo, J.F.; Reitz, S.R.; Lei, Z.R.; Wu, S.Y. A global invasion by the thrip, Frankliniella occidentalis: Current virus vector status and its management. Insect Sci. 2020, 27, 626–645. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.Y.; Lin, Y.Y.; Tuan, S.J.; Tang, L.C.; Chi, H.; Atlıhan, R.; Özgökçe, M.S.; Güncan, A. Integrating demography, predation rate, and computer simulation for evaluation of Orius strigicollis as biological control agent against Frankliniella intonsa. Entomol. Gen. 2021, 41, 179–196. [Google Scholar] [CrossRef]
- Campbell, A.; Frazer, B.D.; Gilbert, N.G.A.P.; Gutierrez, A.P.; Mackauer, M. Temperature requirements of some aphids and their parasites. J. Appl. Ecol. 1974, 11, 431–438. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, F.; Mu, W.; Wang, Q.H.; Li, H. Comparison of Bradysia odoriphaga Yang and Zhang reared on artificial diet and different host plants based on an age-stage, two-sex life table. Phytoparasitica 2015, 43, 107–120. [Google Scholar] [CrossRef]
- Tu, X.Y.; Chen, Y.; Zhi, Y. Effects of light-emitting diode illumination on insect behavior and biological characters. Plant Prot. 2014, 40, 11–15. [Google Scholar]
- Yang, Y.T.; Li, W.X.; Xie, W.; Wu, Q.J.; Xu, B.Y.; Wang, S.L.; Li, C.R.; Zhang, Y.J. Development of Bradysia odoriphaga (Diptera: Sciaridae) as affected by humidity: An age–stage, two-sex, life-table study. Appl. Entomol. Zool. 2015, 50, 3–10. [Google Scholar] [CrossRef]
- Fischer, K.; Kölzow, N.; Höltje, H.; Karl, I. Assay conditions in laboratory experiments: Is the use of constant rather than fluctuating temperatures justified when investigating temperature-induced plasticity? Oecologia 2011, 166, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.Y.; Liu, Y.Q.; Zhang, L.; Cheng, Y.X.; Lou, L.Z.; Jiang, X.F. Effects of temperatures on the development and reproduction of the armyworm, Mythimna roseilinea: Analysis using an age-stage, two-sex life table. J. Integr. Agric. 2018, 17, 1506–1515. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | n | Mean ± SE |
|---|---|---|
| Egg | 82 | 8.00 ± 0.00d |
| First nymphs (L1) | 61 | 5.03 ± 0.06f |
| Second nymphs (L2) | 50 | 7.00 ± 0.29e |
| Third nymphs (L3) | 43 | 7.58 ± 0.40de |
| Fourth nymphs (L4) | 31 | 7.23 ± 0.37e |
| Fifth nymphs (L5) | 31 | 11.26 ± 0.74c |
| Preadult | 31 | 45.42 ± 1.29 |
| Female adult longevity | 15 | 26.67 ± 3.82a |
| Male adult longevity | 16 | 19.00 ± 3.14ab |
| Total longevity of females | 15 | 73.20 ± 4.17 |
| Total longevity of males | 16 | 63.38 ± 3.37 |
| Mean longevity of all the individuals | 100 | 34.02 ± 2.62 |
| Sex ratio (female/male) | 0.94 (15/16) | - |
| Adult preoviposition period (APOP) | 15 | 13.56 ± 1.34 |
| Total preoviposition period (TPOP) | 15 | 61.33 ± 1.94 |
| Oviposition days (Od) | 15 | 5.67 ± 1.20 |
| Mean fecundity (F) (eggs/female) | 15 | 56.60 ± 16.94 |
| Population Parameters | Mean ± SE |
|---|---|
| Cohort size (n) | 100 |
| Intrinsic rate of increase (r) (d−1) | 0.0321 ± 0.0066 |
| Finite rate of increase (λ) (d−1) | 1.0327 ± 0.0068 |
| Net reproductive rate (R0) (offspring/individual) | 8.49 ± 3.19 |
| Mean generation time (T) (d) | 66.55 ± 0.70 |
| Stage | n | Mean ± SE |
|---|---|---|
| Egg | 74 | 7.04 ± 0.10d |
| First larvae (L1) | 74 | 4.26 ± 0.12e |
| Second larvae (L2) | 74 | 3.45 ± 0.12f |
| Third larvae (L3) | 74 | 2.78 ± 0.11g |
| Fourth larvae (L4) | 74 | 1.89 ± 0.08h |
| Fifth larvae (L5) | 74 | 4.38 ± 0.17e |
| Pupa | 72 | 9.25 ± 0.14c |
| Preadult | 72 | 32.86 ± 0.22 |
| Female adult longevity | 36 | 28.28 ± 1.56a |
| Male adult longevity | 36 | 22.44 ± 1.09b |
| Female adult longevity | 36 | 61.28 ± 1.57 |
| Male adult longevity | 36 | 55.17 ± 1.09 |
| Mean longevity of all the individuals | 100 | 44.53 ± 2.37 |
| Sex ratio (female/male) | 1 (36/36) | - |
| Adult preoviposition period (APOP) | 36 | 5.29 ± 0.40 |
| Total preoviposition period (TPOP) | 36 | 38.15 ± 0.45 |
| Oviposition days (Od) | 36 | 19.18 ± 1.15 |
| Mean fecundity (F) (eggs/female) | 36 | 200.64 ± 30.45 |
| Population Parameters | Mean ± SE |
|---|---|
| Cohort size (n) | 100 |
| Intrinsic rate of increase (r) (d−1) | 0.0951 ± 0.0053 |
| Finite rate of increase (λ) (d−1) | 1.0998 ± 0.0058 |
| Net reproductive rate (R0) (offspring/individual) | 72.23 ± 14.47 |
| Mean generation time (T) (d) | 45.00 ± 0.65 |
| Stage | Pj | Uj | ||
|---|---|---|---|---|
| nj | Mean ± SE | mj | Mean ± SE | |
| Second nymphs | 50 | 4.60 ± 0.16 | 61 | 4. 11 ± 0.26 |
| Third nymphs | 43 | 9.09 ± 0.60 | 50 | 8.76 ± 0.59 |
| Fourth nymphs | 31 | 16.77 ± 1.19 | 43 | 16.42 ± 1.21 |
| Fifth nymphs | 31 | 39.48 ± 3.33 | 31 | 39.48 ± 3.33 |
| Preadult | 31 | 69.26 ± 4.17 | 100 | 26.19 ± 3.34 |
| Female adult | 15 | 210.47 ± 3.87 | 15 | 210.47 ± 3.87 |
| Male adult | 16 | 107.44 ± 4.00 | 16 | 107.44 ± 4.00 |
| Adult | 31 | 157.29 ± 21.56 | 31 | 157.29 ± 21.56 |
| Predation Parameters | Mean ± SE |
|---|---|
| Net predation rate (C0) (prey/individual) | 74.95 ± 12.32 |
| Finite predation rate (ω) (d−1) | 0.98 ± 0.09 |
| Transformation rate (Qp) (C0/R0) | 8.83 ± 6.79 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, K.; Lan, R.; Yang, X.; Gong, B.; Zhang, J.; Zhou, X.; Jin, L. Two-Sex Life Table Analysis of the Predator Arma chinensis (Hemiptera: Pentatomidae) and the Prediction of Its Ability to Suppress Populations of Scopula subpunctaria (Lepidoptera: Geometridae). Agriculture 2023, 13, 1254. https://doi.org/10.3390/agriculture13061254
Cao K, Lan R, Yang X, Gong B, Zhang J, Zhou X, Jin L. Two-Sex Life Table Analysis of the Predator Arma chinensis (Hemiptera: Pentatomidae) and the Prediction of Its Ability to Suppress Populations of Scopula subpunctaria (Lepidoptera: Geometridae). Agriculture. 2023; 13(6):1254. https://doi.org/10.3390/agriculture13061254
Chicago/Turabian StyleCao, Kunqian, Rongmeng Lan, Xiuju Yang, Bing Gong, Jingjing Zhang, Xia Zhou, and Linhong Jin. 2023. "Two-Sex Life Table Analysis of the Predator Arma chinensis (Hemiptera: Pentatomidae) and the Prediction of Its Ability to Suppress Populations of Scopula subpunctaria (Lepidoptera: Geometridae)" Agriculture 13, no. 6: 1254. https://doi.org/10.3390/agriculture13061254
APA StyleCao, K., Lan, R., Yang, X., Gong, B., Zhang, J., Zhou, X., & Jin, L. (2023). Two-Sex Life Table Analysis of the Predator Arma chinensis (Hemiptera: Pentatomidae) and the Prediction of Its Ability to Suppress Populations of Scopula subpunctaria (Lepidoptera: Geometridae). Agriculture, 13(6), 1254. https://doi.org/10.3390/agriculture13061254