


Characteristics of Schisandra chinensis (Turcz.) Baillon Collected in Habitats of South Korea



Abstract
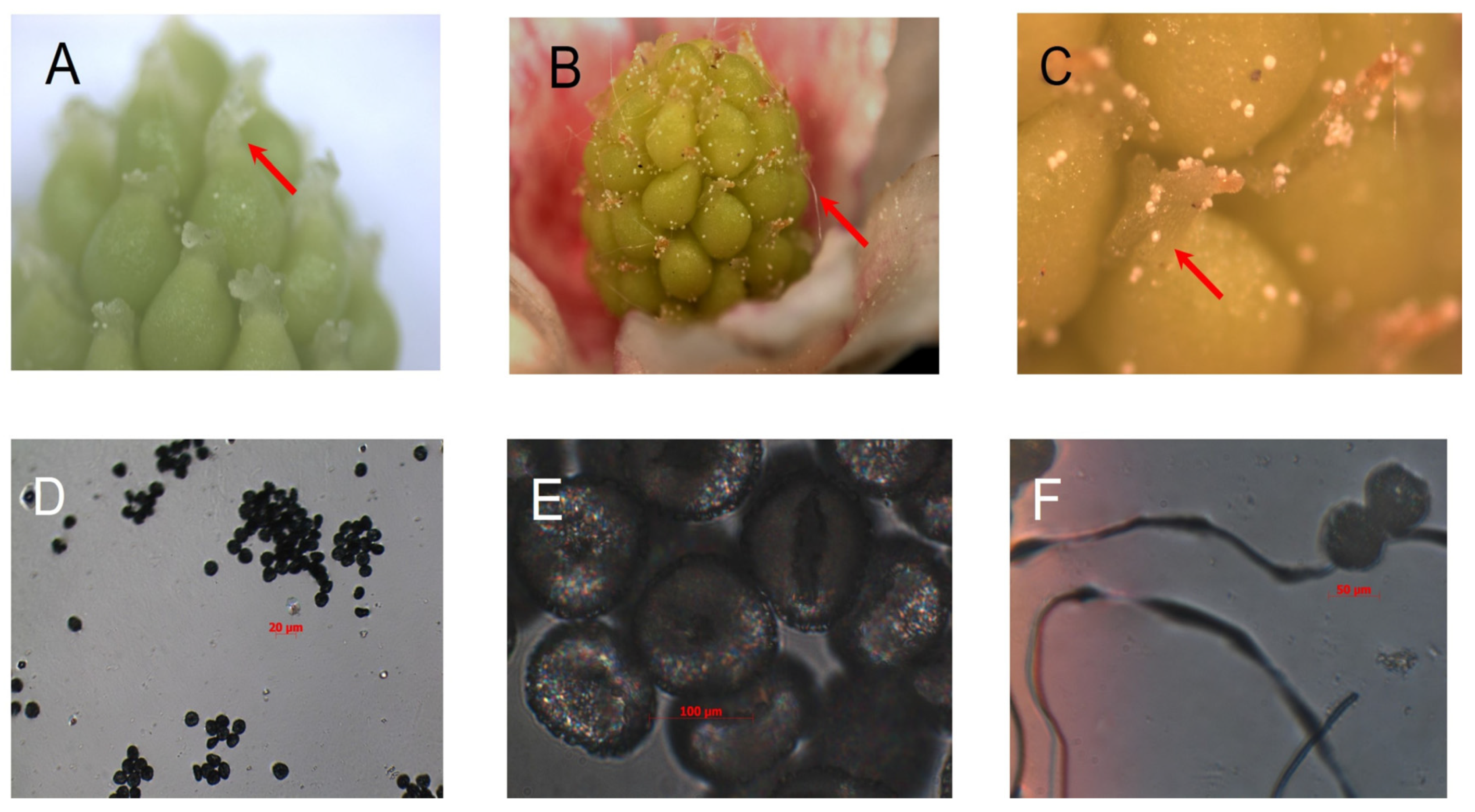
:1. Introduction
2. Materials and Methods
2.1. Design of Field Trial
2.2. Evaluation of Agronomic Traits
2.3. Evaluation of Schizandrin, Gomisin A, and Gomisin N
2.4. Statistical Analysis
2.5. PCA
2.6. Hierarchical Cluster Analysis
3. Results
3.1. ANOVA for Phenotypic Measurements
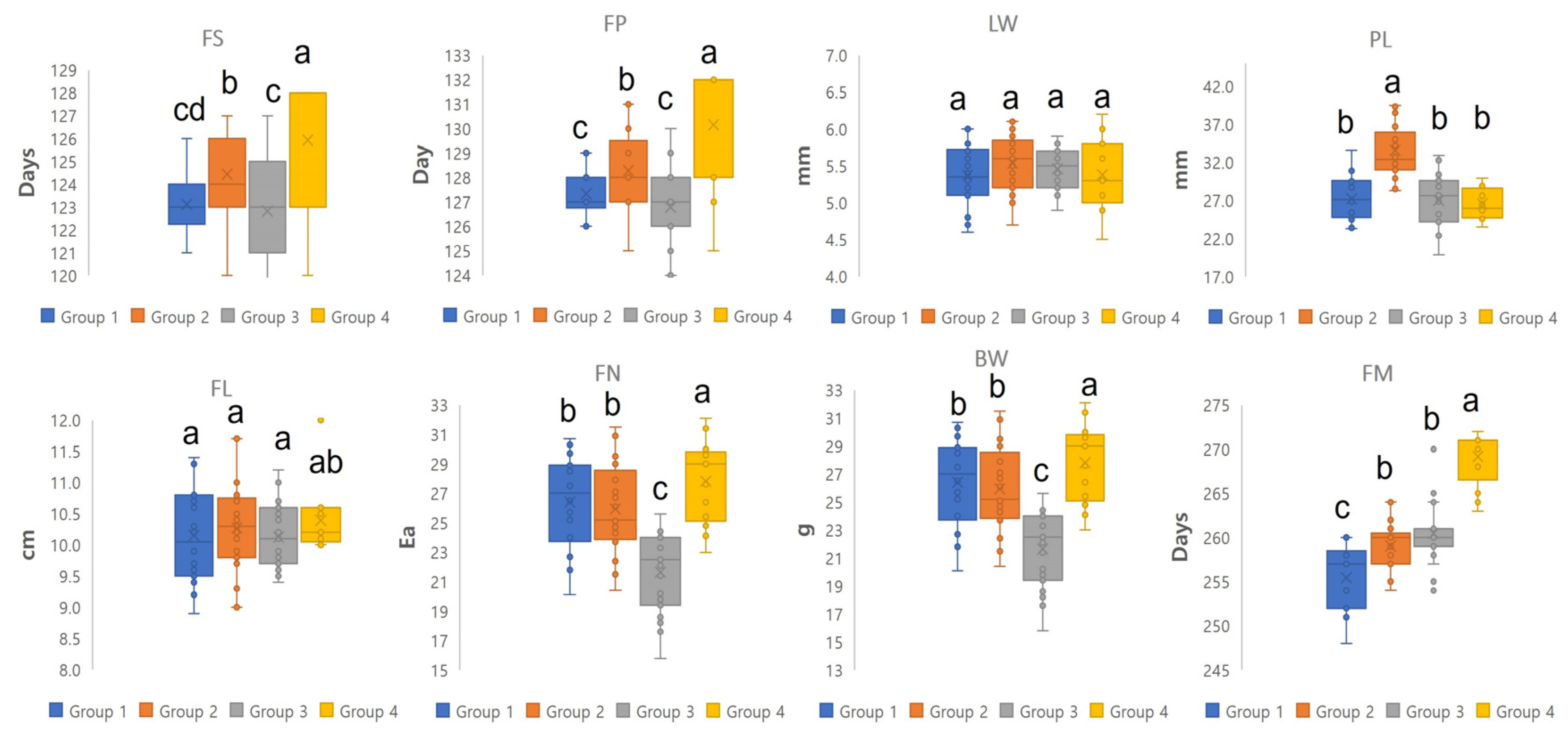
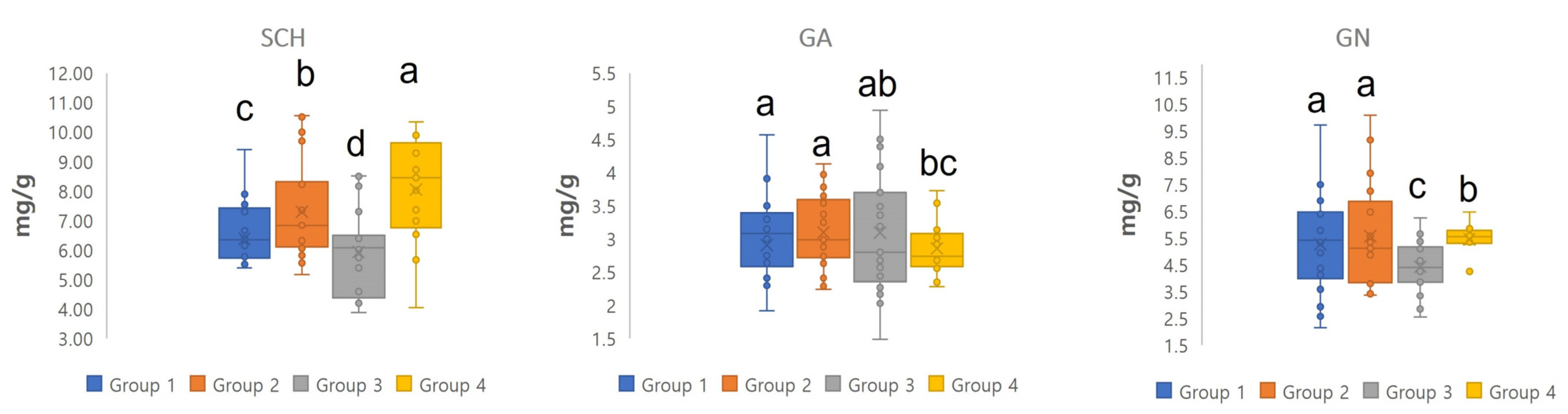
3.2. Phenotypic Traits and Lignan Compounds of Schisandra chinensis
3.3. Correlation Analysis of 96 Accessions of Schisandra chinensis
3.4. PCA of 96 Accessions of Schisandra chinensis
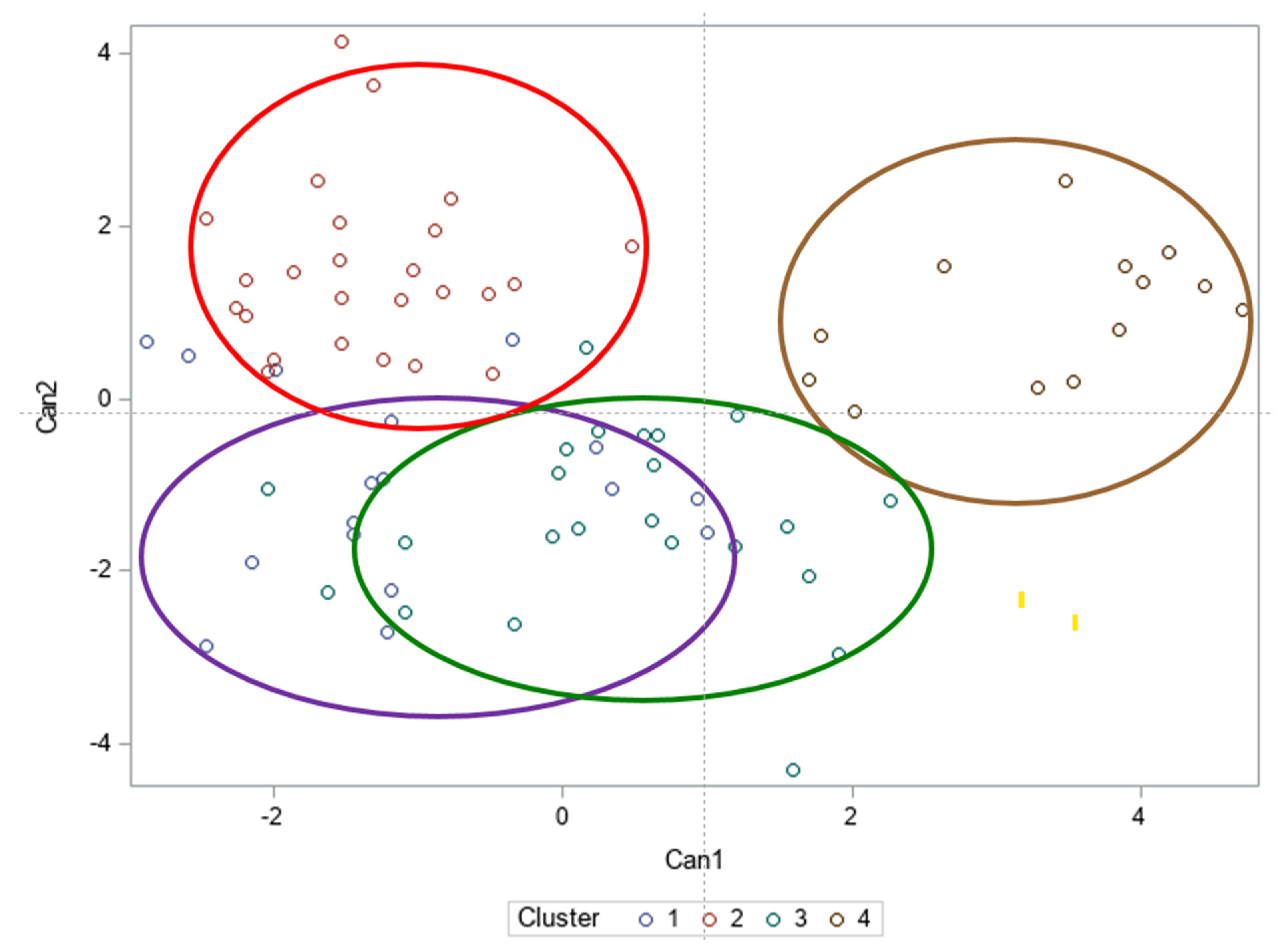
3.5. Hierarchical Cluster Analysis of 96 Accessions of Schisandra chinensis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Foster, S. An illustrate guide. In 101 Medicinal Herbs; Interweave Press: Loveland, OH, USA, 1998. [Google Scholar]
- Saunders, R.M.K. Systematic Botany Monographs. In Monograph of Schisandra (Schisandraceae); American Society of Plant Taxonomists: Ann Arbor, MI, USA, 2000; Volume 58, pp. 3–20. [Google Scholar] [CrossRef]
- Ueda, K. Sex change in a woody vine species, Schisandra chinensis, a preliminary note. J. Jpn. Bot. 1988, 63, 319–321. [Google Scholar] [CrossRef]
- Ministry of Agriculture, Food and Rural Affairs (MAFRA). Production Statistics of Special Crop; Ministry of Agriculture, Food and Rural Affairs: Sejong, Republic of Korea, 2021; Volume 48, pp. 23–48. [Google Scholar]
- Lee, J.S. Literature review on the Omija activities in the Dongeuibogam. J. East Asian Soc. Diet. Life 1995, 5, 1–6. [Google Scholar]
- Chae, Y.B.; Kim, W.J.; Gi, Y.P.; Ahan, M.J.; No, Y.J. Overview of the Korea Useful Plant Accessions Research; Korea Research Institute of Chemical Technology: Daejeon, Republic of Korea, 1988; p. 832. [Google Scholar]
- Jiang, S.; Fan, X.; Wang, Z. Hepato-protective effects of six Schisandra lignans on acetaminophen induced liver injury are partially associated with the inhibition of CYP-mediated bioactivation. Chem. Biol. Interact. 2015, 231, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Panossian, A.; Wikman, G. Pharmacology of Schisandra chinensis Baillon, an overview of Russian research and uses in medicine. J. Ethnopharm. 2008, 118, 183–212. [Google Scholar] [CrossRef]
- Xu, L.; Grandi, N.; Del Vecchio, C.; Mandas, D.; Corona, A.; Piano, D.; Esposito, F.; Parolin, C.; Tramontano, E. From the traditional Chinese medicine plant Schisandra chinensis new scaffolds effective on HIV-1 reverse transcriptase resistant to non-nucleoside inhibitors. J. Microbiol. 2015, 53, 288–293. [Google Scholar] [CrossRef]
- Wang, Q.Y.; Deng, L.L.; Liu, J.J.; Zhang, J.X.; Hao, X.J.; Mu, S.Z. Schisanhenol derivatives and their biological evaluation against tobacco mosaic virus (TMV). Fitoterapia. 2015, 101, 117–124. [Google Scholar] [CrossRef]
- Hwang, D.; Shin, S.Y.; Lee, Y.; Hyun, J.Y.; Yong, Y.J.; Park, J.C.; Lee, Y.H.; Lim, Y.G. A compound isolated from Schisandra chinensis induces apoptosis. Bioorg. Med. Chem. Lett. 2011, 21, 6054–6057. [Google Scholar] [CrossRef] [PubMed]
- Waiwut, P.; Shin, M.S.; Yokoyama, S. Gomisin A enhances tumor necrosis factor-α-induced G1 cell cycle arrest via signal transducer and activator of transcription 1-mediated phosphorylation of retinoblastoma protein. Biol. Pharm. Bull. 2012, 35, 1997–2003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casarin, E.; Dall’Acqua, S.; Smejkal, K. Molecular mechanisms of antiproliferative effects induced by Schisandra-derived dibenzocyclooctadiene lignans (+)-deoxyschisandrin and (−)-gomisin N in human tumor cell lines. Fitoterapia 2014, 98, 241–247. [Google Scholar] [CrossRef]
- Yim, S.Y.; Lee, Y.J.; Lee, Y.K.; Jung, S.E.; Kim, J.H.; Kim, J.E.; Kim, H.J. Gomisin N isolated from Schisandra chinensis significantly induces antiproliferative and proapoptotic effects in hepatic carcinoma. Mol. Med. Rep. 2009, 2, 725–732. [Google Scholar] [CrossRef] [Green Version]
- Jeong, E.J.; Lee, H.K.; Lee, K.Y.; Jeon, B.J.; Kim, D.H.; Park, J.H.; Song, J.H.; Huh, J.M.; Lee, J.H.; Sung, S.H. The effects of lignan-riched extract of Schisandra chinensis on amyloid-β induced cognitive impairment and neurotoxicity in the cortex and hippocampus of mouse. J. Ethnopharm. 2013, 146, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Sa, F.; Zhang, L.Q.; Chong, C.M.; Guo, B.J.; Li, S. Discovery of novel anti-parkinsonian effect of schisantherin A in in vitro and in vivo. Neurosci. Lett. 2015, 593, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Mao, X.; Zhao, X.; Liu, Z.; Liu, B.; Li, H. Gomisin N isolated from Schisandra chinensis augments pentobarbital-induced sleep behaviors through the modification of the serotonergic and GABAergic system. Fitoterapia 2014, 96, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Chiu, P.Y.; Luk, K.F.; Leung, H.Y. Schisandrin B stereoisomers protect against hypoxia/reoxygenation-induced apoptosis and inhibit associated changes in Ca2+-induced mitochondrial permeability transition and mitochondrial membrane potential in H9c2 cardiomyocytes. Life Sci. 2008, 82, 1092–1101. [Google Scholar] [CrossRef]
- Park, H.J.; Cho, J.Y.; Kim, M.K.; Koh, P.O.; Cho, K.W.; Kim, C.H.; Lee, K.S.; Chung, B.Y.; Kim, G.S.; Cho, J.H. Anti-obesity effect of Schisandra chinensis in 3T3-L1 cells and high fat diet-induced obese rats. Food Chem. 2012, 134, 227–234. [Google Scholar] [CrossRef]
- Jang, M.K.; Yun, Y.R.; Kim, J.H.; Park, M.H.; Jung, M.H. Gomisin N inhibits adipogenesis and prevents high-fat diet-induced obesity. Sci. Rep. 2017, 7, 40345. [Google Scholar] [CrossRef] [Green Version]
- Yang, J. The evaluation on the effective as a cosmetic material of oil extracted from Schisadra chinensis seed. J. Korean Oil Chem. Soc. 2012, 29, 231–237. [Google Scholar]
- Quirin, K.W. Supercritical Schisandra extracts a new concept for personal care cosmetics. Cosmet. Sci. Technol. 2008, 1, 28. [Google Scholar]
- Skrypchenko, N.; Kushnir, N.; Sljusar, G. Schisandra chinensis in the collection of the M. Grishko National Botanical Garden of the Ukrainian NAS in Kyiv. Ann. Warsaw Univ. Life Sci.—SGGW—Hortic. Landsc. Archit. 2018, 50, 43–50. [Google Scholar] [CrossRef]
- Kozo, P. The floral mechanism of Woo-we-zy Schizandra chinensis (Turcz.) Baillon. C. R. (Dokl.) l’Acad. Sci. l’URSS 1946, 53, 749–751. [Google Scholar]
- Yuan, L.C.; Luo, Y.B.; Thien, L.B.; Fan, J.H.; Xu, H.L.; Chen, Z.D. Pollination of Schisandra henryi (Schisandraceae) by female, pollen-eating Megommata species (Cecidomyiidae, Diptera) in south-central China. Ann. Bot. 2007, 99, 451–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.N.; Huang, S.J.; Zhao, J.M.; Zhang, Y.W. Gender variation in a monoecious woody vine Schisandra chinensis (Schisandraceae) in northeast China. Ann. Bot. Fennici. 2013, 50, 21. [Google Scholar] [CrossRef]
- Park, C.G.; Chang, Y.H.; Kim, D.H. Characteristics of flower and fruit in collected Schisandra chinensis Baillon. Korean J. Med. Crop Sci. 1995, 3, 35–39. [Google Scholar]
- Han, S.H.; Jang, J.K.; Ma, K.H.; Kim, Y.J.; Kim, S.M.; Lee, H.J. Selection of superior accessions through analysis of growth characteristics and physiological activity of Schisandra chiensis Collection. Korean J. Med. Crop Sci. 2019, 27, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.Y.; Kim, C.S.; Gang, Y.J.; Lee, Y.G.; Jung, U.J.; Jung, J.S.; Sin, S.C.; Kim, H.J. Development of Natural Beverage and a Special Components and Methods of Mass Production and Cultivation Technique of Schisandra nigra as Special-Product of Cheju; Ministry of Agriculture, Food and Rural Affairs: Sejong, Republic of Korea, 1999. [Google Scholar]
- Kim, J.Y.; Park, C.B.; Kim, D.H.; Lim, J.R.; Choi, Y.G. A new Schisandraceae vine (Schisandra chinensis) cultivar, “Chung sun”. Korean J. Breed. 2004, 36, 229. [Google Scholar] [CrossRef]
- Ryo, H.C. A study on the categorization of the strategy group of program provider (PP). J. Korean Data Inf. Sci. Soc. 2008, 19, 913–924. [Google Scholar]
- Fu, J.E.; Feng, L.; Wei, S.G.; Ma, X.J.; Huang, R.S.; Feng, S.X. Distinctive morphological characteristics contribute to the identification of Artemisia annua L. accessions with high yield and high artemisinin content. J. Appl Res. Med. Aromat Plants 2016, 3, 43–47. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.R.; Kim, C.S.; Kim, J.Y.; You, D.H.; Man Kim, J.M.; Sun Kim, Y.S.; Song, E.J.; Kim, Y.G.; Seob Ahn, Y.S.; Choi, D.G. Changes of antioxidant activity and lignan contents in Schisandra chinensis by harvesting times. Korean J. Med. Crop Sci. 2011, 19, 414–420. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.S.; Park, C.G.; Bang, J.K. Varietal and yearly differences of lignin contents in fruit of collected lines of Schizandra chinensis Baillon. Korean J. Med. Crop Sci. 2003, 3, 35–39. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Accessions | Altitude (m) | Collection Site |
|---|---|---|
| 8 | 361 | Bongseongmyeon, Bonghwagun, Gyeongsangbukdo |
| 5 | 450 | Jaesanmyeon, Bonghwagun, Gyeongsangbukdo |
| 4 | 433 | Beopjeonmyeon, Bonghwagun, Gyeongsangbukdo |
| 3 | 419 | Chunyangmyeon, Bonghwagun, Gyeongsangbukdo |
| 23 | 520 | Dongnomyeon, Mungyeongsi, Gyeongsangbukdo |
| 12 | 687 | Daegangmyeon, Danyanggun, Chungcheongbukdo |
| 10 | 706 | Sangdongeup, Yeongwolgun, Gangwondo |
| 10 | 878 | Hwangjidong, Taebaeksi, Gangwondo |
| 8 | 359 | Irwolmyeon, Yeongyanggun, Gyeongsangbukdo |
| 11 | 545 | Subimyeon, Yeongyanggun, Gyeongsangbukdo |
| 2 | 471 | Jinaneup, Jinangun, Jeollabuk-do |
| Source of Variation | DF | F Value | ||||
| FS (1) | FP | FFF | LW | LL | ||
| Year (Y) | 1 | 775.92 *** | 832.54 *** | 1.36 ns (2) | 97.70 *** | 160.94 *** |
| Genotype (G) | 95 | 1.68 *** | 4.88 ** | 2.17 ** | 9.46 *** | 4.55 *** |
| Y × G | 95 | 0.28 ns | 0.06 ns | 0.04 ns | 0.72 ns | 1.32 ns |
| Source of Variation | DF | F Value | ||||
| PL | PM | ANT | FL | FW | ||
| Y | 1 | 74.64 *** | 0.79 ns | 2.59 ns | 5.15 * | 0.25 ns |
| G | 95 | 1.40 ns | 1.42 ns | 0.81 ns | 1.81 * | 2.64 *** |
| Y × G | 95 | 0.72 ns | 1.12 ns | 0.00 ns | 0.88 ns | 0.50 ns |
| Source of Variation | DF | F Value | ||||
| FN | BW | TWB | FM | |||
| Y | 1 | 0.66 ns | 0.43 ns | 1.93 ns | 31.12 *** | |
| G | 95 | 1.22 *** | 12.68 ns | 1.29 ns | 25.30 *** | |
| Y × G | 95 | 0.50 ns | 0.79 ns | 0.83 ns | 0.15 ns | |
| Source of Variation | DF | F Value | ||||
| Schizandrin | Gomisin A | Gomisin N | ||||
| Y | 1 | 718,521.00 *** | 405,053.00 *** | 190,368.00 *** | ||
| G | 95 | 975.97 *** | 372.59 *** | 301.89 *** | ||
| Y × G | 95 | 872.38 *** | 339.36 *** | 268.26 *** | ||
| Corrected Total | 155 | |||||
| Traits (Unit) (1) | Maximum | Minimum | Mean ± S.D | C.V. (%) |
|---|---|---|---|---|
| FS (days) | 128.0 | 122.0 | 123.7 ± 7.6 | 1.9 |
| FP (days) | 131.5 | 126.5 | 127.7 ± 0.4 | 1.6 |
| FFF (%) | 95.0 | 0.0 | 75.7 ± 1.8 | 23.7 |
| LW (cm) | 6.9 | 3.9 | 5.4 ± 0.0 | 7.7 |
| LL (cm) | 17.1 | 6.4 | 9.2 ± 0.1 | 11.8 |
| PL (cm) | 4.8 | 1.4 | 2.9 ± 0.5 | 17.5 |
| PM (%) | 25.0 | 0.0 | 3.5 ± 0.3 | 101.6 |
| ANT (%) | 25.0 | 0.0 | 1.2 ± 0.3 | 381.3 |
| FL (cm) | 12.8 | 8.0 | 10.2 ± 0.1 | 6.1 |
| FW (cm) | 12.3 | 7.8 | 9.8 ± 0.1 | 6.8 |
| FN (ea) | 36.2 | 14.7 | 25.1 ± 0.3 | 16.9 |
| BW (g) | 26.9 | 6.1 | 14.2 ± 0.3 | 22.4 |
| TWB (g/plant) | 5844.0 | 109.0 | 1367.5 ± 87.5 | 71.3 |
| FM (days) | 273.0 | 248.5 | 260.0 ± 2.8 | 2.0 |
| Schizandrin (mg/g) | 14.5 | 2.2 | 6.9 ± 0.1 | 28.3 |
| Gomisin A (mg/g) | 9.8 | 0.9 | 3.2 ± 0.1 | 25.9 |
| Gomisin N (mg/g) | 12.2 | 2.1 | 5.2 ± 0.1 | 9.5 |
| White | Light Pink | Dark Pink | |
|---|---|---|---|
| Accessions | 12 | 61 | 23 |
| Traits | FS (1) | FP | FFF | LW | LL | PL | PM | ANT | FL | FW | FN | BW | TW B | FM |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FS | 1 | |||||||||||||
| FP | 0.990 *** | 1 | ||||||||||||
| FFF | 0.192 * | 0.178 * | 1 | |||||||||||
| LW | 0.516 *** | 0.534 *** | −0.074 | 1 | ||||||||||
| LL | 0.702 *** | 0.708 *** | −0.030 | 0.705 *** | 1 | |||||||||
| PL | −0.518 *** | −0.514 *** | −0.013 | −0.302 *** | −0.405 *** | 1 | ||||||||
| PM | −0.121 | −0.112 | −0.028 | −0.107 | −0.118 | −0.131 | 1 | |||||||
| ANT | 0.111 | 0.111 | 0.097 | −0.064 | 0.104 | −0.107 | 0.017 | 1 | ||||||
| FL | 0.110 | 0.120 | −0.162 * | 0.160 * | 0.069 | −0.057 | −0.186 * | 0.005 | 1 | |||||
| FW | 0.011 | 0.025 | −0.213 ** | 0.160 * | 0.044 | −0.057 | −0.122 | −0.088 | 0.812 *** | 1 | ||||
| FN | −0.029 | −0.010 | 0.209 ** | −0.002 | −0.052 | 0.175 * | −0.051 | −0.002 | −0.109 | −0.017 | 1 | |||
| BW | 0.209 ** | 0.231 ** | −0.023 | 0.283 *** | 0.233 ** | −0.014 | −0.200 * | −0.020 | 0.516 *** | 0.627 *** | 0.630 *** | 1 | ||
| TWB | −0.140 | −0.129 | 0.247 ** | 0.079 | −0.089 | 0.177 * | −0.039 | 0.022 | 0.007 | 0.073 | 0.397 *** | 0.321 *** | 1 | |
| FM | 0.243 ** | 0.244 ** | −0.122 | −0.022 | −0.031 | −0.212 ** | −0.075 | −0.08 | 0.134 | −0.023 | 0.083 | 0.074 | −0.210 ** | 1 |
| Traits (1) | Schizandrin | Gomisin A | Gomisin N |
|---|---|---|---|
| FS | 0.143 | −0.522 **** | 0.425 **** (2) |
| FP | 0.154 | −0.535 **** | 0.419 **** |
| FFF | 0.223 * | 0.128 | 0.016 |
| LW | −0.027 | −0.346 **** | 0.206 ** |
| LL | −0.115 | −0.436 **** | 0.335 **** |
| PL | −0.026 | 0.400 **** | −0.234 ** |
| PM | −0.042 | 0.102 | 0.009 |
| ANT | 0.151 | −0.072 | 0.102 |
| FL | −0.040 | −0.256 ** | 0.060 |
| FW | −0.112 | −0.272 *** | 0.077 |
| FN | 0.194 | 0.146 | 0.196 * |
| BW | −0.005 | −0.263 *** | 0.245 * |
| TWB | 0.022 | 0.083 | −0.047 |
| FM | 0.233 ** | −0.153 | 0.148 |
| Schizandrin | 1 | 0.006 | 0.234 ** |
| Gomisin A | 0.007 | 1 | −0.272 *** |
| Gomisin N | 0.234 ** | −0.271 *** | 1 |
| Eigenvalues of the Correlation Matrix | ||||
|---|---|---|---|---|
| Eigenvalue | Difference | Proportion | Cumulative | |
| 1 | 2.799 | 0.181 | 0.215 | 0.215 |
| 2 | 2.618 | 0.884 | 0.201 | 0.417 |
| 3 | 1.735 | 0.275 | 0.133 | 0.550 |
| 4 | 1.460 | 0.518 | 0.112 | 0.662 |
| 5 | 0.941 | 0.063 | 0.072 | 0.735 |
| 6 | 0.878 | 0.096 | 0.068 | 0.802 |
| 7 | 0.783 | 0.200 | 0.060 | 0.863 |
| 8 | 0.583 | 0.090 | 0.045 | 0.907 |
| 9 | 0.492 | 0.044 | 0.038 | 0.945 |
| 10 | 0.449 | 0.294 | 0.035 | 0.980 |
| 11 | 0.155 | 0.097 | 0.012 | 0.992 |
| 12 | 0.058 | 0.009 | 0.005 | 0.996 |
| 13 | 0.049 | 0.004 | 1.000 | |
| Eigenvectors | |||||
|---|---|---|---|---|---|
| Traits (1) | Prin1 | Prin2 | Prin3 | Prin4 | Prin5 |
| SF | 0.500 | −0.121 | 0.084 | 0.249 | −0.277 |
| FP | 0.521 | −0.078 | 0.087 | 0.227 | −0.256 |
| LW | −0.111 | 0.214 | 0.358 | −0.061 | 0.047 |
| LL | 0.065 | −0.091 | 0.131 | 0.539 | 0.617 |
| PL | −0.172 | 0.063 | 0.479 | 0.295 | −0.389 |
| FL | 0.096 | 0.135 | 0.433 | 0.048 | 0.089 |
| FW | 0.074 | 0.495 | −0.244 | 0.080 | −0.020 |
| FN | 0.003 | 0.558 | −0.198 | 0.019 | 0.041 |
| BW | 0.341 | 0.152 | 0.309 | −0.338 | 0.114 |
| FM | 0.238 | 0.511 | 0.124 | −0.104 | 0.051 |
| Schizandrin | 0.331 | −0.120 | −0.307 | −0.227 | −0.210 |
| Gomisin A | 0.360 | −0.152 | −0.013 | −0.191 | −0.503 |
| Gomisin N | −0.073 | −0.156 | 0.345 | −0.532 | −0.002 |
| Traits (1) | Total STD | Within STD | R-Square | RSQ/(1-RSQ) |
|---|---|---|---|---|
| FS | 2.327 | 2.079 | 0.233 | 0.303 |
| FP | 2.040 | 1.715 | 0.320 | 0.472 |
| LW | 0.386 | 0.387 | 0.031 | 0.033 |
| LL | 0.686 | 0.676 | 0.066 | 0.071 |
| PL | 4.363 | 3.129 | 0.505 | 1.022 |
| FL | 0.643 | 0.648 | 0.023 | 0.024 |
| FW | 0.642 | 0.639 | 0.047 | 0.050 |
| FN | 3.661 | 2.891 | 0.400 | 0.668 |
| BW | 3.137 | 2.808 | 0.230 | 0.298 |
| FM | 5.497 | 3.411 | 0.630 | 1.700 |
| Schizandrin | 1.671 | 1.523 | 0.201 | 0.252 |
| Gomisin A | 0.980 | 0.986 | 0.028 | 0.029 |
| Gomisin N | 1.745 | 1.687 | 0.102 | 0.113 |
| OVERALL | 2.639 | 2.011 | 0.441 | 0.790 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, B.S.; Kim, J.S.; Seo, Y.J.; Oh, T.Y.; Lee, J.-D. Characteristics of Schisandra chinensis (Turcz.) Baillon Collected in Habitats of South Korea. Agriculture 2023, 13, 1256. https://doi.org/10.3390/agriculture13061256
Kim BS, Kim JS, Seo YJ, Oh TY, Lee J-D. Characteristics of Schisandra chinensis (Turcz.) Baillon Collected in Habitats of South Korea. Agriculture. 2023; 13(6):1256. https://doi.org/10.3390/agriculture13061256
Chicago/Turabian StyleKim, Beung Sung, Jong Soo Kim, Young Jin Seo, Tae Young Oh, and Jeong-Dong Lee. 2023. "Characteristics of Schisandra chinensis (Turcz.) Baillon Collected in Habitats of South Korea" Agriculture 13, no. 6: 1256. https://doi.org/10.3390/agriculture13061256
APA StyleKim, B. S., Kim, J. S., Seo, Y. J., Oh, T. Y., & Lee, J. -D. (2023). Characteristics of Schisandra chinensis (Turcz.) Baillon Collected in Habitats of South Korea. Agriculture, 13(6), 1256. https://doi.org/10.3390/agriculture13061256