


Exploring the Genetic and Morphological Variation and Disease Resistance in Local and Foreign Prunus persica (L.) Batsch Cultivars

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Leaf and Fruit Morphological Characterization
2.3. Susceptibility to Cylindrocarpon destructans and Monilinia laxa
2.4. Molecular Analysis
2.5. Statistical Analysis
2.5.1. Phenotypic Data
2.5.2. Molecular Data
3. Results and Discussion
3.1. Prospections
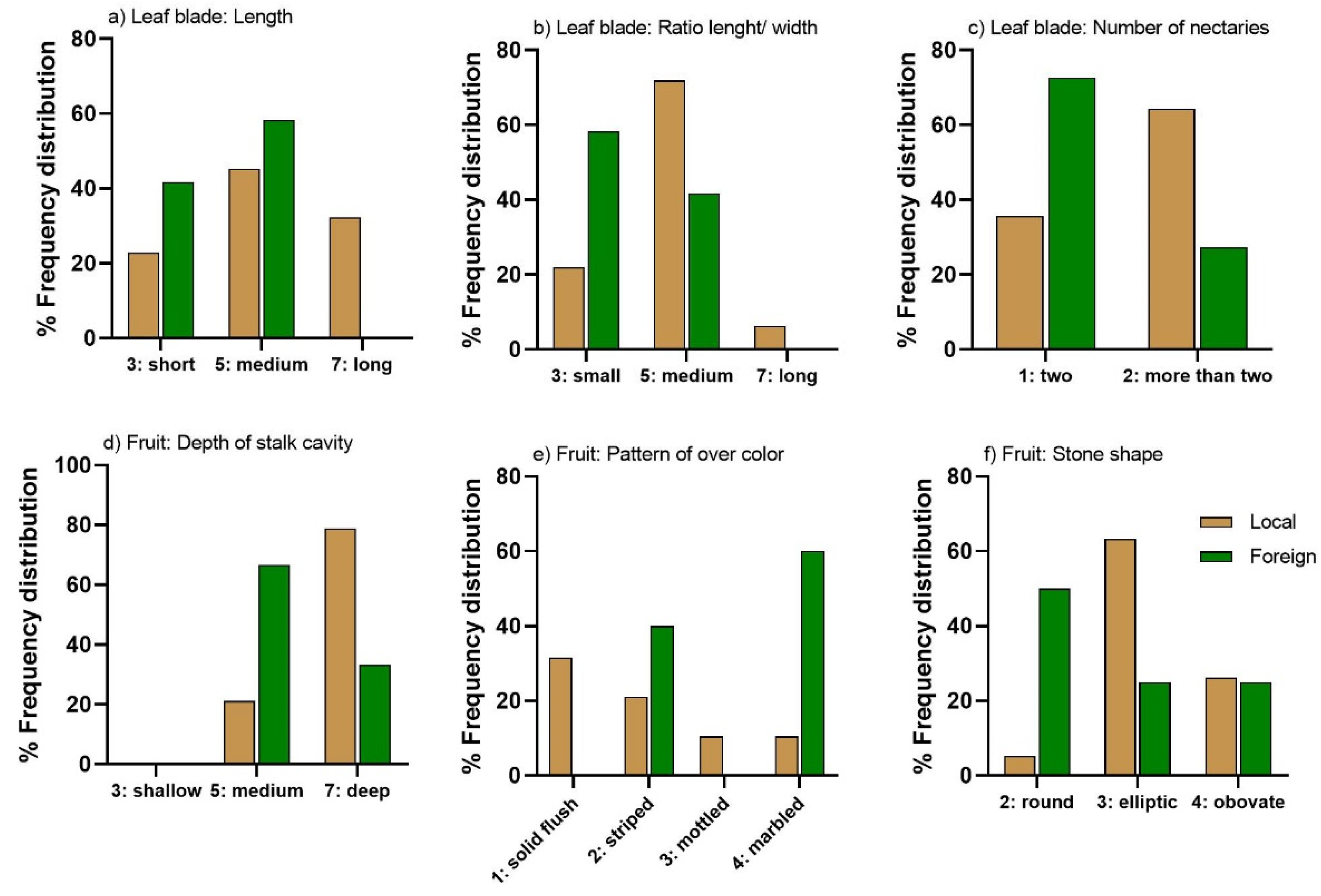
3.2. Leaf Morphological Characterization
3.3. Ripening Time and Fruit Morphological Characterization
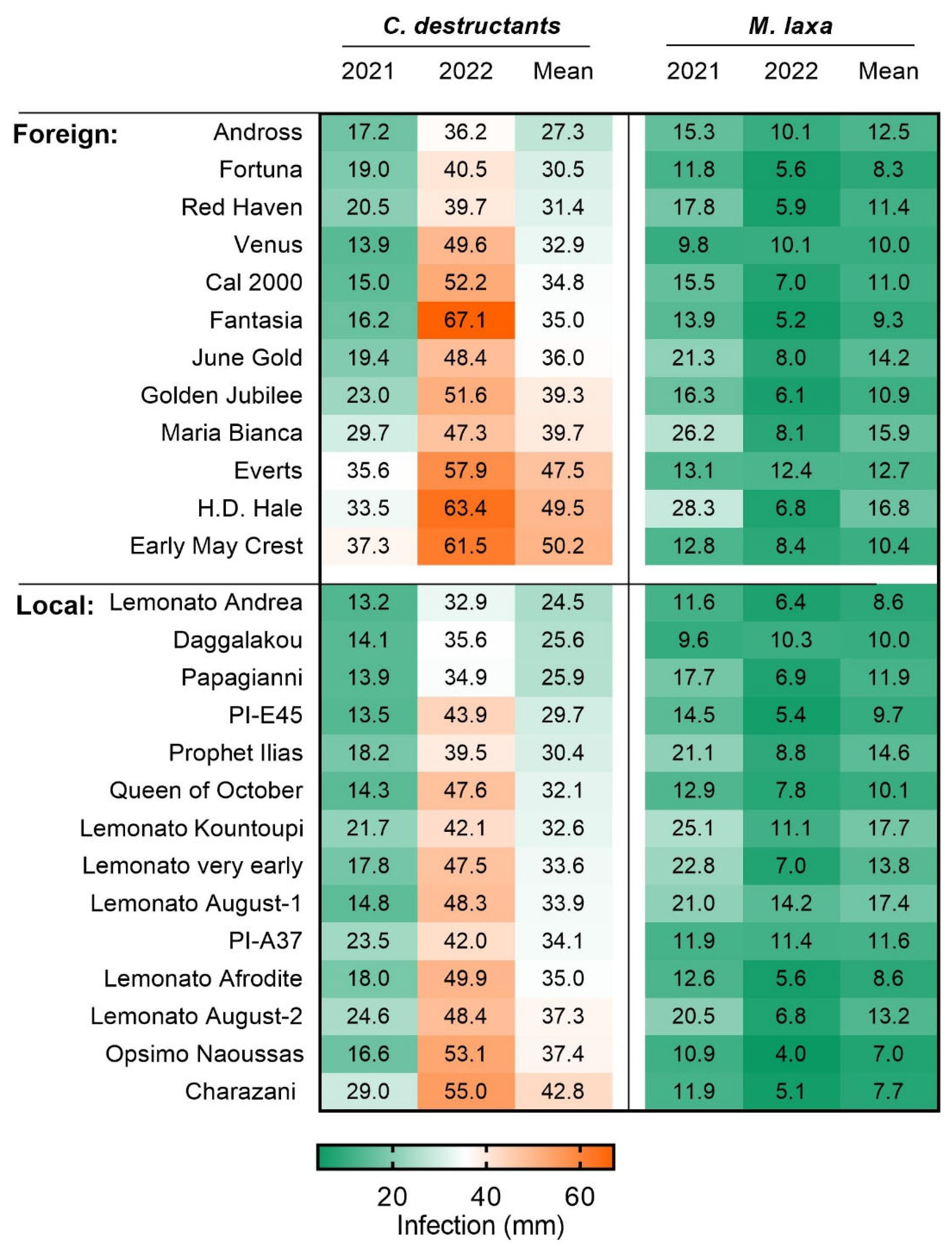
3.4. Susceptibility to Cylindrocarpon destructans and Monilinia laxa
3.5. Correlation Analysis
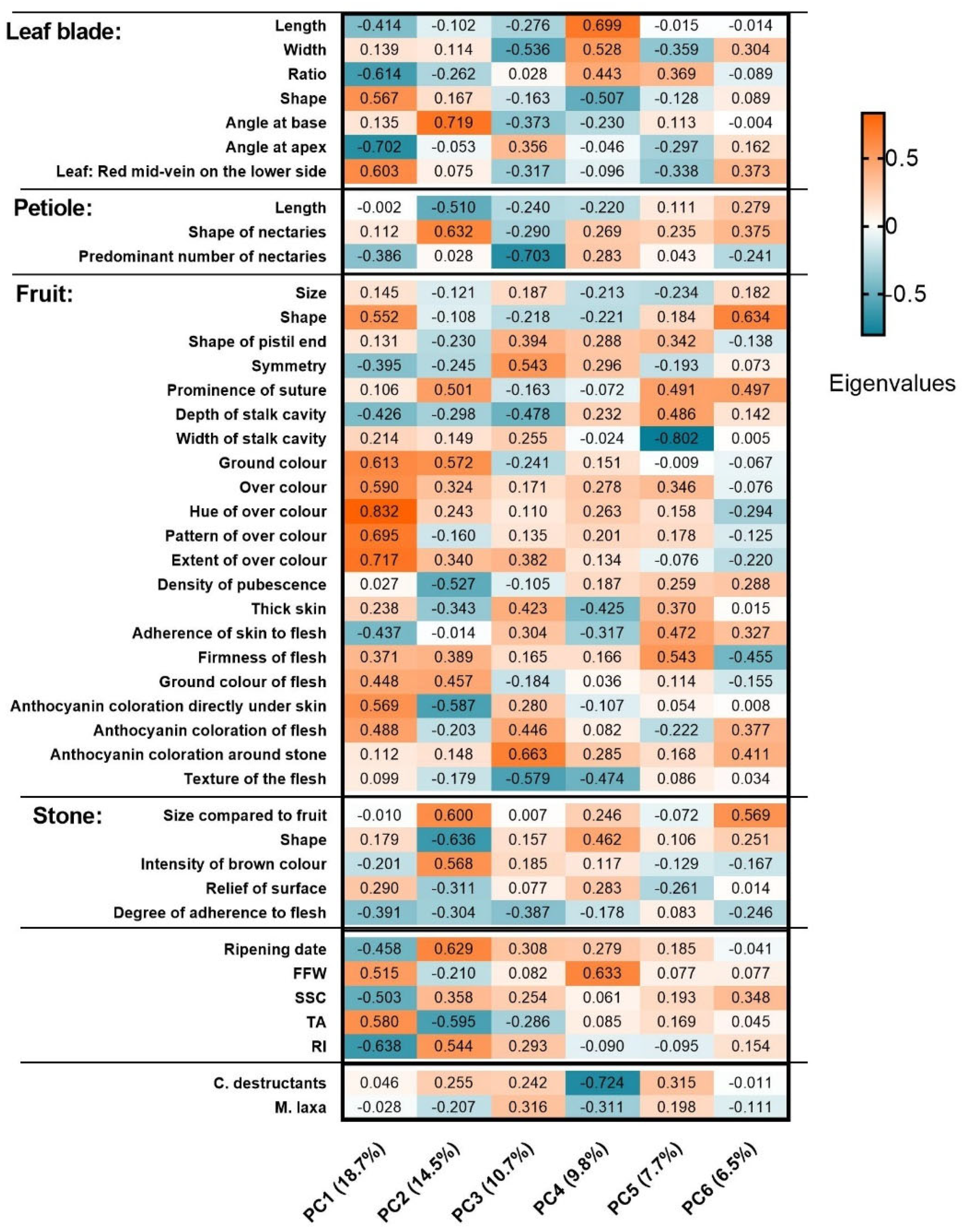
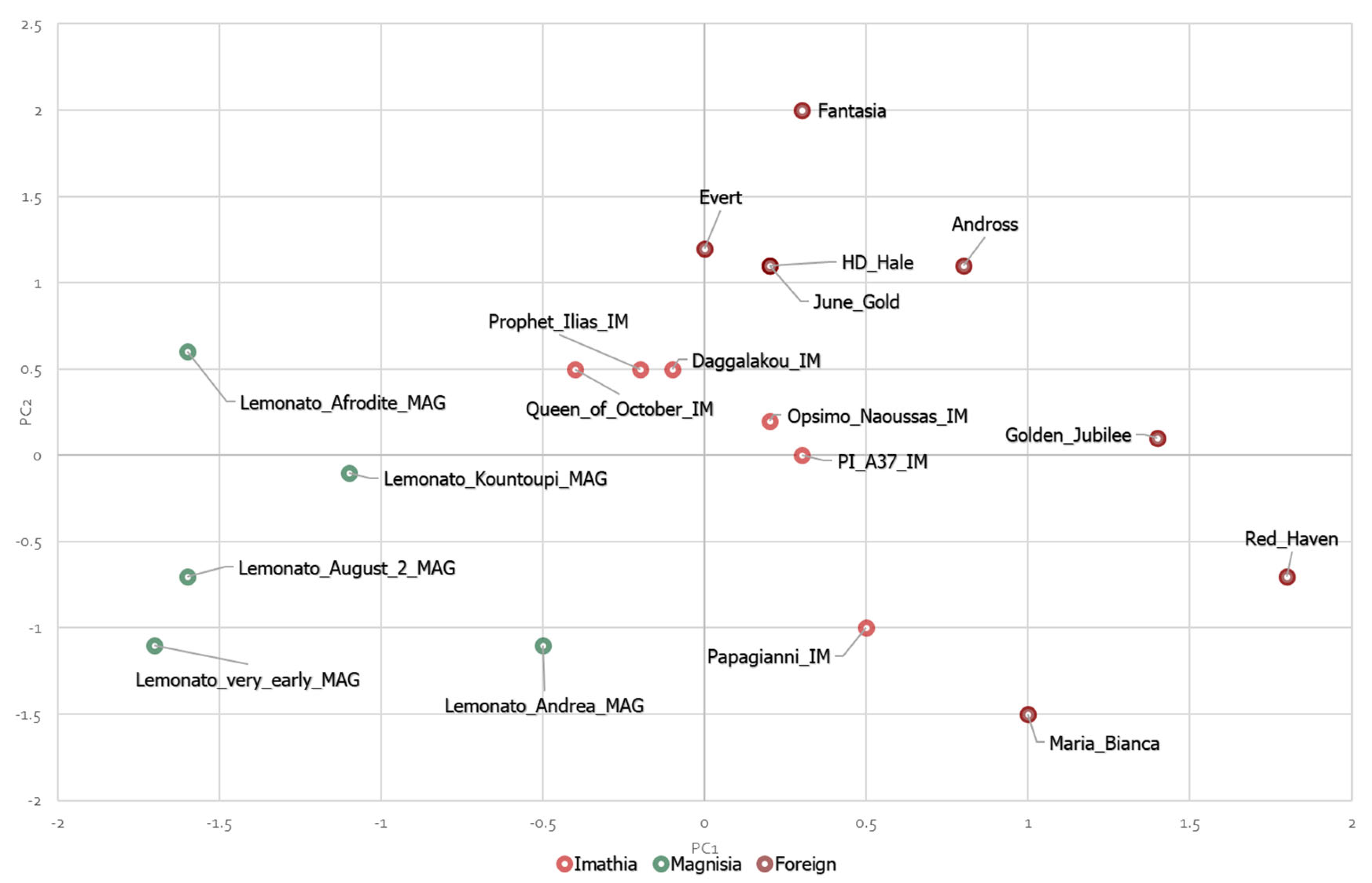
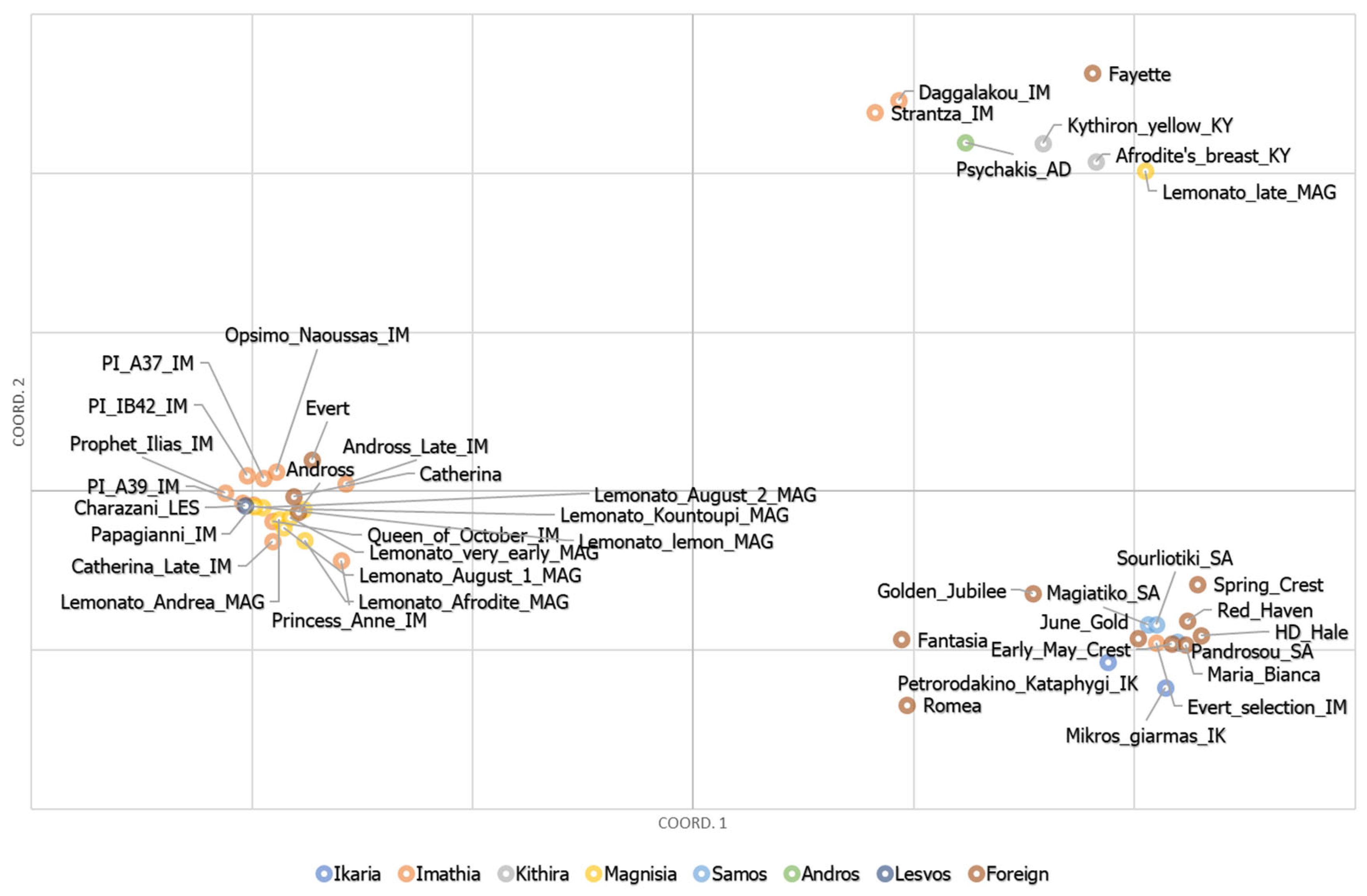
3.6. Principal Component Analyses (PCA)
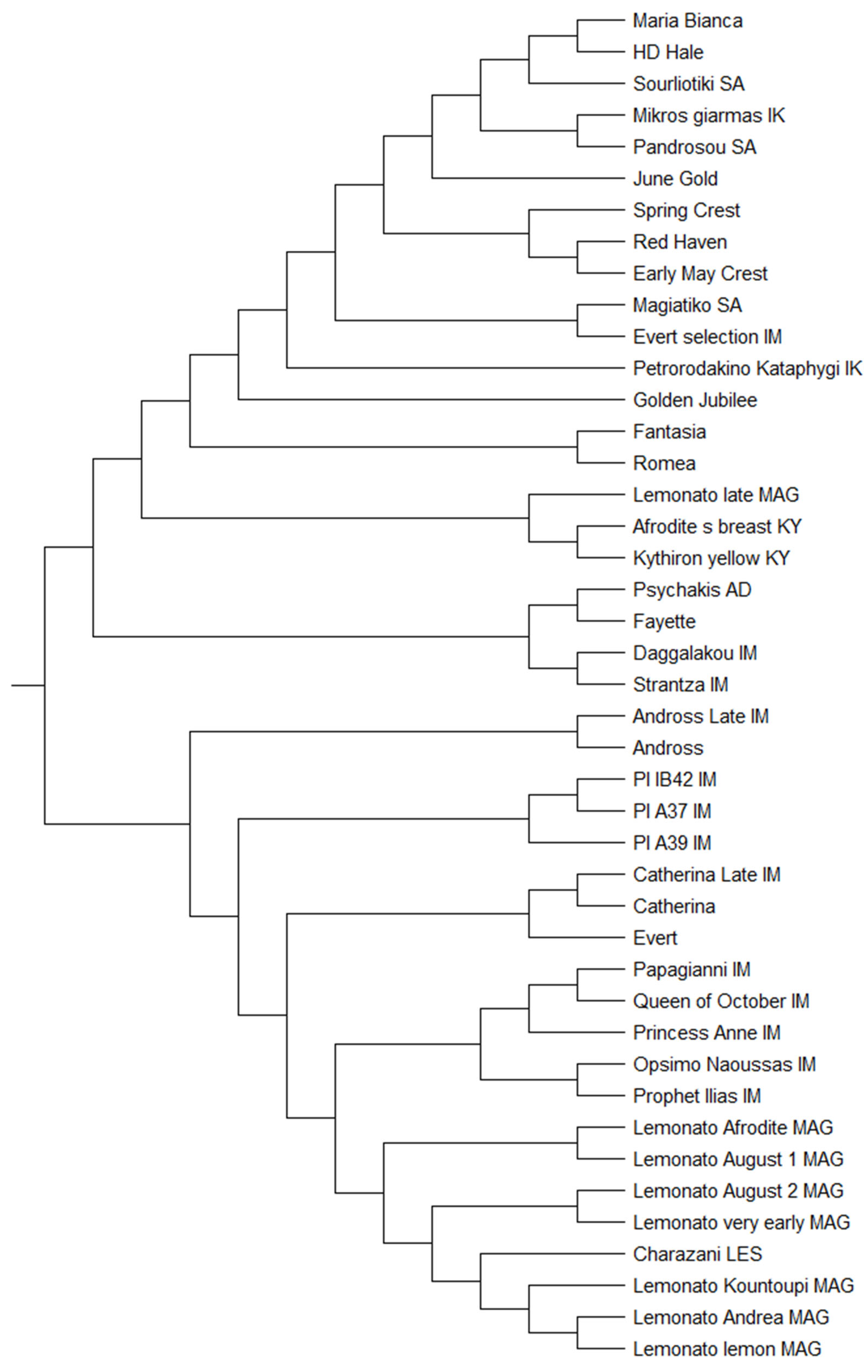
3.7. Genetic Characterization
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scorza, R.; Okie, W.R. Peaches (Prunus). Acta Hortic. 1991, 290, 177–234. [Google Scholar] [CrossRef]
- Bassi, D.; Monet, R. Botany and taxonomy. In The Peach: Botany, Production and Uses; Layne, D.R., Bassi, D., Eds.; CABI Publishing: Oxfordshire, UK, 2008; pp. 1–36. [Google Scholar]
- International Peach Genome Initiative; Verde, I.; Abbott, A.G.; Scalabrin, S.; Jung, S.; Shu, S.; Marroni, F.; Zhebentyayeva, T.; Dettori, M.T.; Grimwood, J.; et al. The high-quality draft genome of peach (Prunus persica) identifies unique patterns of 636 genetic diversity, domestication and genome evolution. Nat. Genet. 2013, 45, 487–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aranzana, M.J.; Abbassi, E.K.; Howad, W.; Arús, P. Genetic variation, population structure and linkage disequilibrium in peach commercial varieties. BMC Genet. 2010, 11, 69. [Google Scholar] [CrossRef] [Green Version]
- Mitsopoulou, N.K.; Nanos, G.D.; Grigoriadou, E.; Katis, N. ‘Lemonato’ peach: A series of cultivated clones with high fruit quality. Acta Hortic. 2019, 1242, 363–368. [Google Scholar] [CrossRef]
- Drogoudi, P.; Pantelidis, G.E.; Goulas, V.; Manganaris, G.A.; Ziogas, V.; Manganaris, A. The appraisal of qualitative parameters and antioxidant contents during postharvest peach fruit ripening underlines the genotype significance. Postharvest Biol. Technol. 2016, 115, 142–150. [Google Scholar] [CrossRef]
- Charou, E.; Kythiri, M.P. Kosmas Panaretos and the ‘Breasts of Aphrodite’. 2017. Available online: https://www.eleniharou.gr/o-kythirios-vouleftis-kosmas-panaretos-ke-i-masti-tis-afroditis/ (accessed on 27 February 2023).
- Oikonomidis, L. The Fruit Trees of Greece; Central Fund for Agriculture, Livestock and Forestry: Athens, Greece, 1950; p. 245. (In Greek) [Google Scholar]
- Koukourgiannis, V. Systematice arboriculture in central-west Macedonia completed its 80 years (1927–2007). Ga. -Ktinotrofia 2008, 1, 34–40. (In Greek) [Google Scholar]
- OPEKEPE. Greek Payment Authority of Common Agricultural Policy (C.A.P.) Aid Schemes. 2016. Available online: www.opekepe.gr (accessed on 15 December 2022).
- Dini, M.; do Carmo Bassols Raseira, M.; Ueno, B. Blossom blight resistance in peach: Heritability and segregation in progenies from reciprocal crosses. Rev. Ceres 2021, 68, 555–563. [Google Scholar] [CrossRef]
- Montenegro, J.; Obi, V.I.; Barriuso, J.J.; Gogorcena, Y. Identification of ‘Calanda’-type Peach genotypes tolerant to Monilinia laxa (Aderh. & Ruhland) Honey. Agronomy 2022, 12, 2662. [Google Scholar] [CrossRef]
- Koball, D.C.; Wilcox, W.F.; Seem, R.C. Influence of incubation—Period humidity on the development of brown rot blossom blight of sour cherry. Phytopathology 1997, 87, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Morgan, D.P.; Michailides, T.J. Risk analysis of brown rot blossom blight of prune caused by Monilinia fructicola. Phytopathology 2001, 91, 759–768. [Google Scholar] [CrossRef] [Green Version]
- Dini, M.; do Carmo Bassols Raseira, M.; Scariotto, S.; Carpenedo, S.; Ueno, B. Peach and nectarine susceptibility to brown rot and protocol optimization to evaluate Monilinia fructicola sporulation. Acta Sci. Agron. 2022, 44, e55850. [Google Scholar] [CrossRef]
- Mustafa, M.; Bassi, D.; Corre, M.N.; Lino, L.; Signoret, V.; Quilot-Turion, B.; Cirilli, M. Phenotyping brown rot susceptibility in stone fruit: A literature review with emphasis on peach. Horticulturae 2021, 7, 115. [Google Scholar] [CrossRef]
- Thomidis, T.; Michaildies, T.J. Development and implementation of cost-effective strategies to manage brown rot of peach trees in Imathia, Greece. Eur. J. Plant Pathol. 2010, 126, 575–582. [Google Scholar] [CrossRef]
- Govi, G. Two species of Cylindrocarpon on fruit trees. Ann. Sper. Agrar. 1952, 6, 793–804. [Google Scholar]
- Tewoldemedhin, Y.T.; Mazzola, M.; Mostert, L.; McLeod, A. Cylindrocarpon species associated with apple tree roots in South Africa and their quantification using real-time PCR. Eur. J. Plant Pathol. 2011, 129, 637–651. [Google Scholar] [CrossRef]
- Isutsa, D.K.; Merwin, I.A. Malus germplasm varies in resistance or tolerance to apple replant disease in a mixture of New York orchard soils. HortScience 2000, 35, 262–268. [Google Scholar] [CrossRef] [Green Version]
- Arús, P.; Verde, I.; Sosinski, B.; Zhebentyayeva, T.; Abbott, A.G. The peach genome. Tree Genet. Genomes 2012, 8, 531–547. [Google Scholar] [CrossRef]
- Dirlewanger, E.; Graziano, E.; Joobeur, T.; Garriga-Calderé, F.; Cosson, P.; Howad, W.; Arús, P. Comparative mapping and marker-assisted selection in Rosaceae fruit crops. Proc. Natl. Acad. Sci. USA 2004, 101, 9891–9896. [Google Scholar] [CrossRef] [Green Version]
- Marimon, N.; Luque, J.; Arús, P.; Eduardo, I. Fine mapping and identification of candidate genes for the peach powdery mildew resistance gene Vr3. Hortic. Res. 2020, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, I.; Bassi, D.; Eduardo, I.; Ciacciulli, A.; Pirona, R.; Rossini, L.; Vecchietti, A. Qtl mapping for brown rot (Monilinia fructigena) resistance in an intraspecific peach (Prunus persica L. Batsch) F1 progeny. Tree Genet. Genomes 2014, 10, 1223–1242. [Google Scholar] [CrossRef]
- Fan, S.; Bielenberg, D.G.; Zhebentyayeva, T.N.; Reighard, G.L.; Okie, W.R.; Holland, D.; Abbott, A.G. Mapping quantitative trait loci associated with chilling requirement, heat requirement and bloom date in peach (Prunus persica). New Phytol. 2010, 185, 917–930. [Google Scholar] [CrossRef]
- Peace, C.; Norelli, J. Genomics approaches to crop improvement in the Rosaceae. In Genomics of the Rosaceae; Folta, K.M., Gardiner, S.E., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 19–54. [Google Scholar]
- Boudehri, K.; Bendahmane, A.; Cardinet, G.; Troadec, C.; Moing, A.; Dirlewanger, E. Phenotypic and fine genetic characterization of the D locus controlling fruit acidity in peach. BMC Plant Biol. 2009, 9, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dirlewanger, E.; Cosson, P.; Boudehri, K.; Renaud, C.; Capdeville, G.; Tauzin, Y.; Laigret, F.; Moing, A. Development of a second-generation genetic linkage map for peach [Prunus persica (L.) Batsch] and characterization of morphological traits affecting flower and fruit. Tree Genet. Genomes 2006, 3, 1–13. [Google Scholar] [CrossRef]
- Cantini, C.; Cimato, A.; Sani, G. Morphological evaluation of olive germplasm present in Tuscany region. Euphytica 1999, 109, 173–181. [Google Scholar] [CrossRef]
- Sharma, P.; Sharma, R. DNA fingerprinting of peach (Prunus persica) germplasm in accessing genetic variation using arbitrary oligonucleotide markers system. Indian J. Biotechnol. 2018, 17, 484–491. [Google Scholar]
- Joobeur, T.; Viruel, M.A.; de Vicente, M.C.; Jáuregui, B.; Ballester, J.; Dettori, M.T.; Verde, I.; Truco, M.J.; Messeguer, R.; Batlle, I.; et al. Construction of a saturated linkage map for Prunus using an almond×peach F2 progeny. Theor. Appl. Genet. 1998, 97, 1034–1041. [Google Scholar] [CrossRef]
- Yamamoto, T.; Mochida, K.; Imai, T.; Shi, Y.Z.; Ogiwara, I.; Hayashi, T. Microsatellite markers in peach [Prunus persica (L.) Batsch] derived from an enriched genomic and cDNA libraries. Mol. Ecol. Notes 2002, 2, 298–301. [Google Scholar] [CrossRef]
- Butiuc-Keul, A.; Coste, A.; Postolache, D.; Laslo, V.; Halmagyi, A.; Cristea, V.; Farkas, A. Molecular Characterization of Prunus Cultivars from Romania by Microsatellite Markers. Horticulturae 2022, 8, 291. [Google Scholar] [CrossRef]
- Howad, W.; Yamamoto, T.; Dirlewanger, E.; Testolin, R.; Cosson, P.; Cipriani, G.; Monforte, A.J.; Georgi, L.; Abbott, A.G.; Arús, P. Mapping with a few plants: Using selective mapping for microsatellite saturation of the Prunus reference map. Genetics 2005, 171, 1305–1309. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Pan, L.; Wei, B.; Niu, L.; Cui, G.; Wang, L.; Zeng, W.; Wang, Z. Fine Mapping of the Gene Controlling the Fruit Skin Hairiness of Prunus persica and Its Uses for MAS in Progenies. Plants 2021, 10, 1433. [Google Scholar] [CrossRef]
- Zietkiewicz, E.; Rafalski, A.; Labuda, D. Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplification. Genomics 1994, 20, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Parashuram, S.N.V.; Singh, N.N.; Gaikwad, G.; Corrado, P.R.; Sowjanya, B.; Basile, N.S.; Devaraja, R. Chandra. K.D.; Babu, P.G.; Patil, P.; et al. Marathe Morphological, Biochemical, and Molecular Diversity of an Indian Ex Situ Collection of Pomegranate (Punica granatum L.). Plants 2022, 11, 3518. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, E.; Pinar, H.; Uzun, A.; Yaman, M.; Sumbul, A.; Ercisli, S. Identification of genetic diversity among Juglans regia L. genotypes using molecular, morphological, and fatty acid data. Genet. Resour. Crop. Evol. 2021, 68, 1425–1437. [Google Scholar] [CrossRef]
- Karapetsi, L.G.; Pantelidis, E.D.; Pratsinakis, P.; Drogoudi, P. Madesis Fruit Quality Traits and Genotypic Characterization in a Pomegranate Ex Situ (Punica granatum L.) Collection in Greece. Agriculture 2021, 11, 482. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Ioannidou, S.; Nikoloudakis, N.; Seraphides, N.; Papayiannis, L.C.; Kyratzis, A.C. Physicochemical Characterization and Trait Stability in a Genetically Diverse Ex Situ Collection of Pomegranate (Punica granatum L.) Germplasm from Cyprus. Sci. Hortic. 2020, 263, 109116. [Google Scholar] [CrossRef]
- Ganopoulos, I.V.; Kazantzis, K.; Chatzicharisis, I.; Karayiannis, I.; Tsaftaris, A.S. Genetic diversity, structure and fruit trait associations in Greek sweet cherry cultivars using microsatellite based (SSR/ISSR) and morpho-physiological markers. Euphytica 2011, 181, 237–251. [Google Scholar] [CrossRef]
- Pitt, J.; Hocking, A. Fungi and Food Spoilage; Blackie Academic and Professional: London, UK, 2009. [Google Scholar] [CrossRef]
- Barnett, H.L.; Hunter, B.B. Illustrated Genera of Imperfect Fungi, 4th ed.; APS Press: St. Paul, MN, USA, 1998; 218p. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A Rapid DNA Isolation Procedure for Small Quantities of Fresh Leaf Tissues. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic Analysis in Excel. Population Genetic Software for Teaching and Research—An Update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Amiryousefi, A.; Hyvönen, J.; Poczai, P. IMEC: Online Marker Efficiency Calculator. Appl. Plant Sci. 2018, 6, e01159. [Google Scholar] [CrossRef]
- The European Peach Catalogue. Peach accessions except cultivars. In The European Cooperative Programme for the Conservation and Exchange of Crop Genetic Resources; Nordic Gene Bank: Alnarp, Sweden, 1989; pp. 15–37. ISBN 91-87814-04-8. [Google Scholar]
- Douma, K.; Koutis, K.; Thanopoulos, R.; Tsigou, R.; Galanidis, A.; Bebeli, P. Diversity of agricultural plants on Lesvos Island (Northeast Aegean, Greece) with emphasis on fruit trees. Sci. Hortic. 2016, 210, 65–84. [Google Scholar] [CrossRef]
- Reig, G.; Alegre, S.; Gatius, F. Adaptability of peach cultivars [Prunus persica (L.) Batsch] to the climatic conditions of the Ebro Valley, with special focus on fruit quality. Sci. Hortic. 2015, 190, 149–160. [Google Scholar] [CrossRef]
- Byrne, D.H.; Raseira, M.B.; Bassi, D.; Piagnani, M.C.; Gasic, K.; Reighard, G.L.; Moreno, M.A.; Pérez, S. The peach. In Fruit Breeding, Handbook of Plant Breeding 8; Badenes, M.L., Byrne, D.H., Eds.; Springer: Berlin, Germany, 2012; pp. 505–569. [Google Scholar]
- Quilot, B.; Kervella, J.; Génard, M. Shape, mass and dry matter content of peaches of varieties with different domestication levels. Sci. Hortic. 1997, 99, 387–393. [Google Scholar] [CrossRef]
- Crisosto, C.H.; Crisosto, G.M. Relationship between ripe soluble solids concentration (RSSC) and consumer acceptance of high and low acid melting flesh peach and nectarine (Prunus persica (L.) Batsch) cultivars. Postharvest Biol. Technol. 2005, 38, 239–246. [Google Scholar] [CrossRef]
- Traquair, J.A.; White, G.P. Cylindrocarpon rot of fruit trees in cold storage. Can. J. Plant Pathol. 1992, 14, 310–314. [Google Scholar] [CrossRef]
- Obi, V.I.; Barriuso, J.J.; Moreno, M.A.; Giménez, R.; Gogorcena, Y. Optimizing protocols to evaluate brown rot (Monilinia laxa) susceptibility in peach and nectarine fruits. Australas. Plant Pathol. 2017, 46, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Obi, V.I.; Barriuso, J.J.; Gogorcena, Y. Peach brown rot: Still in search of an ideal management option. Agriculture 2018, 8, 125. [Google Scholar] [CrossRef] [Green Version]
- Villarino, M.; Egüen, B.; Lamarca, N.; De Cal, A. Occurrence of Monilinia laxa and M. fructigena after introduction of M. fructicola in peach orchards in Spain. Eur. J. Plant Pathol. 2013, 137, 835–845. [Google Scholar] [CrossRef]
- Pascal, T.; Kervella, J.; Pfeiffer, F.G.; Sauge, M.H.; Esmenjaud, D. Evaluation of the interspecific progeny Prunus persica cv Summergrand × Prunus davidiana for disease resistance and some agronomic features. Acta Hortic. 1998, 465, 185–191. [Google Scholar] [CrossRef]
- Bradshaw, A.D. Evolutionary significance of phenotypic plasticity in plants. Adv. Genet. 1965, 13, 115–155. [Google Scholar]
- Cirilli, M.; Baccichet, I.; Chiozzotto, R.; Silvestri, V.; Rossini, L.; Bassi, D. Genetic and phenotypic analyses reveal major quantitative loci associated to fruit size and shape traits in a non-flat peach collection (P. persica L. Batsch). Hortic. Res. 2021, 8, 232. [Google Scholar] [CrossRef] [PubMed]
- Cantín, C.M.; Gogorcena, Y.; Moreno, M.A. Phenotypic diversity and relationships of fruit quality traits in peach and nectarine [Prunus persica (L.) Batsch] breeding progenies. Euphytica 2010, 171, 211–226. [Google Scholar] [CrossRef] [Green Version]
- Dirlewanger, E.; Moing, A.; Rothan, C.; Svanella, L.; Pronier, V.; Guye, A.; Plomion, C.; Monet, R. Mapping QTLs controlling fruit quality in peach (Prunus persica (L.) Batsch). Theor. Appl. Genet. 1999, 98, 18–31. [Google Scholar] [CrossRef]
- Drogoudi, P.D.; Vemmos, S.; Pantelidis, G.; Petri, E.; Tzoutzoukou, C.; Karayiannis, I. Physical characters and antioxidant, sugar and mineral nutrient contents in fruit from 29 apricot (Prunus armeniaca L.) cultivars and hybrids. J. Agric. Food Chem. 2008, 56, 10754–10760. [Google Scholar] [CrossRef]
- Ruiz, D.; Egea, J. Phenotypic diversity and relationships of fruit quality traits in apricot (Prunus armeniaca L.) germplasm. Euphytica 2008, 163, 143–158. [Google Scholar] [CrossRef]
- Drogoudi, P.; Pantelidis, G. Phenotypic variation and peel contribution to fruit antioxidant contents in European and Japanese plums. Plants 2022, 11, 1338. [Google Scholar] [CrossRef]
- Bakić, I.V.; Rakonjac, V.S.; Čolić, S.D.; Fotirić Akšić, M.M.; Nikolić, D.T.; Radović, A.R.; Rahović, D.D. Agro-morphological characterization and evaluation of a Serbian vineyard peach [Prunus persica (L.) Batsch] germplasm collection. Sci. Hortic. 2017, 225, 668–675. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Location | Cultivar/Accession | Flesh Type | Flesh Color |
|---|---|---|---|---|
| Psychakis | Andros | Accession | MPE | YE |
| Mikros giarmas (July peach) | Ikaria | Accession | MPE | YE |
| Petrorodakino Kataphygi | Ikaria | Accession | MPE | YE |
| Andross late | Imathia | Selection | NMPE | YE |
| Catherina late | Imathia | Selection | NMPE | YE |
| Evert selection | Imathia | Selection | NMPE | YE |
| PI-A37 | Imathia | Selection | NMPE | YE |
| PI-A39 | Imathia | Selection | NMPE | YE |
| PI-IB42 | Imathia | Selection | NMPE | YE |
| Daggalakou | Imathia | Cultivar | MPE | YE |
| Opsimo Naoussas | Imathia | Cultivar | MPE | YE |
| Princess Anne | Imathia | Cultivar | MPE | YE |
| Prophet Ilias | Imathia | Cultivar | MPE | YE |
| Kaliga | Imathia | Cultivar | MPE | W |
| Papagianni | Imathia | Cultivar | MPE | W |
| Queen of October | Imathia | Cultivar | MPE | W |
| Strantza | Imathia | Accession | MPE | YE |
| Afrodite’s breast | Kythira | Cultivar | MPE | W |
| Kythiron yellow | Kythira | Accession | MPE | YE |
| Charazani | Lesvos | Accession | MPE | YE |
| Lemonato Afrodite | Magnesia | Accession | MPE | W |
| Lemonato Andrea | Magnisia | Accession | MPE | W |
| Lemonato August-1 | Magnisia | Accession | MPE | W |
| Lemonato August-2 | Magnisia | Accession | MPE | W |
| Lemonato Kountoupi | Magnisia | Accession | MPE | W |
| Lemonato late | Magnisia | Accession | MPE | W |
| Lemonato lemon | Magnisia | Accession | MPE | W |
| Lemonato medium | Magnisia | Accession | MPE | W |
| Lemonato very early | Magnisia | Accession | MPE | W |
| Magiatiko | Samos | Accession | MPE | YE |
| Pandrosou | Samos | Accession | MPE | YE |
| Sourliotiki | Samos | Accession | MPE | YE |
| Fantasia | Foreign | Cultivar | NE | YE |
| Andross | Foreign | Cultivar | NMPE | YE |
| Catherina | Foreign | Cultivar | NMPE | YE |
| Evert | Foreign | Cultivar | NMPE | YE |
| Romea | Foreign | Cultivar | NMPE | YE |
| Early May Crest | Foreign | Cultivar | MPE | YE |
| Fayette | Foreign | Cultivar | MPE | YE |
| Golden Jubilee | Foreign | Cultivar | MPE | YE |
| H.D. Hale | Foreign | Cultivar | MPE | W |
| June Gold | Foreign | Cultivar | MPE | YE |
| Maria Bianca | Foreign | Cultivar | MPE | W |
| Red Haven | Foreign | Cultivar | MPE | YE |
| Spring crest | Foreign | Cultivar | MPE | YE |
| Cultivar | Ripening Day | Fruit Fresh Weight | Soluble Solid Content | Titratable Acidity | Ripening Index |
|---|---|---|---|---|---|
| Local | |||||
| Lemonato very early | 184 | 178.7 | 12.3 | 0.7 | 18.3 |
| Lemonato Andrea | 202 | 208.7 | 11.6 | 1.2 | 9.8 |
| Kalliga | 205 | 224.7 | 12.3 | 1.5 | 8.0 |
| PI-A37 | 206 | 173.1 | 10.7 | 1.1 | 9.5 |
| Papagianni | 209 | 359.2 | 10.8 | 1.1 | 9.5 |
| Lemonato medium | 213 | 303.6 | 11.6 | 0.8 | 14.2 |
| PI-A39 | 218 | 174.6 | 12.0 | 0.7 | 16.6 |
| PI-IB42 | 223 | 203.2 | 10.5 | 0.5 | 19.2 |
| Daggalakou | 225 | 337.1 | 14.0 | 0.9 | 14.9 |
| Lemonato August-2 | 229 | 177.0 | 15.2 | 0.7 | 22.6 |
| Princess Anne | 231 | 238.6 | 14.9 | 0.7 | 20.6 |
| Andross late | 234 | 192.3 | 10.8 | 0.6 | 17.6 |
| Lemonato Kountoupi | 238 | 203.7 | 15.6 | 0.7 | 22.6 |
| Opsimo Naoussas | 238 | 274.5 | 12.5 | 0.6 | 20.6 |
| Lemonato late | 241 | 235.3 | 13.4 | 0.6 | 23.5 |
| Evert selection | 242 | 196.5 | 12.5 | 0.6 | 19.6 |
| Prophet Ilias | 245 | 268.1 | 16.5 | 0.9 | 17.7 |
| Lemonato Afrodite | 247 | 171.7 | 15.8 | 0.6 | 25.1 |
| Queen of October | 265 | 222.9 | 11.9 | 0.6 | 21.0 |
| Foreign | |||||
| Spring crest | 160 | 137.5 | 9.1 | 1.2 | 7.9 |
| June Gold | 176 | 255.0 | 12.9 | 1.2 | 10.8 |
| Red Haven | 182 | 252.0 | 10.3 | 1.2 | 8.9 |
| Maria Bianca | 189 | 275.0 | 11.3 | 1.0 | 10.8 |
| Romea | 189 | 158.6 | 11.5 | 0.5 | 21.3 |
| Golden Jubilee | 194 | 253.3 | 11.6 | 1.2 | 9.6 |
| Catherina | 195 | 179.1 | 13.1 | 0.4 | 30.3 |
| Andross | 218 | 292.8 | 11.8 | 0.7 | 16.1 |
| Fantasia | 221 | 184.3 | 16.6 | 0.6 | 27.1 |
| H.D. Hale | 225 | 198.9 | 14.3 | 0.7 | 19.1 |
| Fayette | 233 | 147.6 | 11.5 | 0.8 | 15.0 |
| Evert | 243 | 171.7 | 10.0 | 0.7 | 14.6 |
| Mean | 217 | 220.9 | 12.5 | 0.8 | 16.9 |
| CV% | 11 | 25 | 16 | 32 | 35 |
| LSD | 48.8 | 1.9 | 0.3 | 6.1 |
| Local | Foreign | |||||
|---|---|---|---|---|---|---|
| Mean | CV% | Mean | CV% | p | ||
| Leaf blade: Length | 3—short; 5—medium; 7—long | 5.8 | 24 | 4.2 | 25 | 0.001 |
| Leaf blade: Width | 3—narrow; 5—medium; 7—large | 5.6 | 27 | 5.5 | 23 | 0.802 |
| Leaf blade: Ratio length/width | 3—small; 5—medium; 7—large | 5.1 | 9 | 3.8 | 27 | 0.000 |
| Leaf blade: Shape in cross section | 1—concave; 2—flat; 3—convex | 1.5 | 52 | 2.1 | 43 | 0.054 |
| Leaf blade: Angle at base | 1—acute; 2—approximately right angle 3—obtuse | 1.4 | 36 | 1.4 | 36 | 0.982 |
| Leaf blade: Angle at apex | 3—small; 5—medium; 7—large | 4.3 | 28 | 3.7 | 27 | 0.159 |
| Leaf: Red mid-vein on the lower side | 1—absent; 9—present | 1.8 | 137 | 3.0 | 121 | 0.302 |
| Petiole: Length | 3—short; 5—medium; 7—long | 5.5 | 27 | 5.5 | 23 | 0.959 |
| Petiole: Shape of nectaries | 1—round; 2—reniform | 1.6 | 32 | 1.6 | 42 | 0.984 |
| Petiole: Predominant number of nectaries | 1—two; 2—more than two | 1.7 | 26 | 1.2 | 49 | 0.005 |
| Fruit: Size | 1—very small; 3—small; 5—medium; 7—large; 9—very large | 5.9 | 23 | 6.5 | 19 | 0.272 |
| Fruit: Shape (in ventral view) | 1—broad; oblate; 2—oblate; 3—round; 4—ovate; 5—elliptic | 3.2 | 30 | 3.2 | 30 | 0.902 |
| Fruit: Shape of pistil end | 1—prominently; pointed; 2—weakly pointed; 3—flat; 4—weakly depressed; 5—strongly depressed | 2.1 | 57 | 2.7 | 49 | 0.185 |
| Fruit: Symmetry (viewed from pistil end) | 1—asymmetric; 2—symmetric | 1.4 | 36 | 1.4 | 36 | 0.982 |
| Fruit: Prominence of suture | 3—weak; 5—medium; 7—strong | 4.9 | 32 | 4.8 | 28 | 0.911 |
| Fruit: Depth of stalk cavity | 3—shallow; 5—medium; 7—deep | 6.6 | 13 | 5.7 | 17 | 0.010 |
| Fruit: Width of stalk cavity | 3—narrow; 5—medium; 7—broad | 5.0 | 27 | 5.3 | 27 | 0.515 |
| Fruit: Ground color | 1—green; 2—cream; green; 3—greenish white; 4—cream white; 5—cream; 6—pink white; 7—greenish yellow; 8—cream yellow; 9—yellow; 10—orange yellow | 6.3 | 54 | 7.9 | 33 | 0.159 |
| Fruit: Overcolor | 1—absent; 9—present | 6.9 | 52 | 7.7 | 41 | 0.547 |
| Fruit: Hue of overcolor | 1—orange red; 2—pink; 3—pink red; 4—light red; 5—medium red; 6—dark red; 7—blackish red | 3.5 | 73 | 4.8 | 48 | 0.190 |
| Fruit: Pattern of overcolor | 1—solid flush; 2—striped; 3—mottled; 4—marbled | 1.9 | 58 | 3.2 | 32 | 0.011 |
| Fruit: Extent of overcolor | 1—very small; 3—small; 5—medium; 7—large; 9—very large | 2.8 | 93 | 4.7 | 54 | 0.056 |
| Fruit: Density of pubescence | 1—very sparse; 3—sparse; 5—medium; 7—dense; 9—very dense | 6.9 | 15 | 6.6 | 40 | 0.644 |
| Fruit: Adherence of skin to flesh | 1—absent; or; very; weak; 3—weak; 5—medium; 7—strong; 9—very; strong | 5.3 | 26 | 5.3 | 22 | 0.971 |
| Fruit: Firmness of flesh | 1—very soft; 3—soft; 5—medium; 7—firm; 9—very firm | 6.3 | 22 | 6.8 | 26 | 0.326 |
| Fruit: Ground color of flesh | 1—greenish white; 2—white; 3—cream white; 4—light yellow; 5—yellow; 6—orange yellow; 7—orange; 8—red | 3.7 | 58 | 5.1 | 34 | 0.081 |
| Fruit: Anthocyanin directly under skin | 1—absent or very weakly expressed; 2—weakly expressed; 3—strongly expressed | 1.1 | 29 | 1.5 | 60 | 0.090 |
| Fruit: Anthocyanin in flesh | 1—absent or very weakly expressed; 2—weakly expressed; 3—strongly expressed | 1.3 | 44 | 1.4 | 56 | 0.686 |
| Fruit: Anthocyanin around stone | 1—absent or very weakly expressed; 2—weakly expressed; 3—strongly expressed | 1.7 | 50 | 1.5 | 53 | 0.453 |
| Fruit: Texture of the flesh | 1—not fibrous; 2—fibrous | 1.8 | 20 | 2.0 | 0 | 0.158 |
| Stone: Size compared to fruit | 3—small; 5—medium; 7—large | 4.9 | 25 | 4.8 | 28 | 0.897 |
| Stone: Shape (in lateral view) | 1—oblate; 2—round; 3—elliptic; 4—obovate | 3.3 | 20 | 2.8 | 31 | 0.050 |
| Stone: Intensity of brown color | 3—light; 5—medium; 7—dark | 6.3 | 16 | 5.7 | 27 | 0.201 |
| Stone: Relief of surface | 1—small pits; 2—large pits; 3—grooves; 4—pits and grooves | 3.9 | 10 | 3.8 | 10 | 0.444 |
| Stone: Degree of adherence to flesh | 3—weak; 5—medium; 7—strong | 5.7 | 29 | 5.3 | 35 | 0.536 |
| Ripening day | Julian day | 226 | 9 | 202 | 13 | 0.006 |
| Fruit fresh weight | g | 228.6 | 25 | 208.8 | 26 | 0.339 |
| Soluble solid content | °Brix | 12.9 | 15 | 12.0 | 17 | 0.260 |
| Titratable acidity | g 100 mL−1 | 0.8 | 33 | 0.9 | 33 | 0.590 |
| Ripening index | Soluble solid content/Titratable acidity | 17.4 | 30 | 16.0 | 45 | 0.540 |
| Cylindrocarpon destructans | Shoot lesion in mm | 32.5 | 16 | 37.9 | 20 | 0.005 |
| Monilinia laxa | Shoot lesion in mm | 11.6 | 29 | 11.9 | 22 | 0.687 |
| Primer | Annealing Temperature (°C) | Fragment Size Range (bp) | Na | Ne | Shannon Index (I) | Gene Diversity (GD) | PIC | Rp |
|---|---|---|---|---|---|---|---|---|
| ISSR (UBC) | ||||||||
| 811 | 58 | 350–1700 | 1.860 | 1.726 | 0.574 | 0.399 | 0.360 | 4.065 |
| 823 | 49 | 500–2500 | 0.930 | 1.233 | 0.238 | 0.154 | 0.356 | 5.721 |
| 826 | 54 | 500–1800 | 0.953 | 1.363 | 0.272 | 0.191 | 0.346 | 6.465 |
| 827 | 47.5 | 600–2000 | 0.977 | 1.426 | 0.318 | 0.225 | 0.350 | 7.116 |
| 834 | 47.5 | 380–1800 | 2.000 | 1.894 | 0.661 | 0.469 | 0.349 | 4.140 |
| 841 | 51.5 | 230–2050 | 2.000 | 1.865 | 0.643 | 0.454 | 0.363 | 6.605 |
| 873 | 51.5 | 650–1700 | 0.953 | 1.240 | 0.241 | 0.156 | 0.380 | 5.442 |
| 880 | 54.5 | 280–1700 | 2.000 | 1.905 | 0.663 | 0.471 | 0.404 | 3.860 |
| Mean | 1.459 | 1.582 | 0.451 | 0.315 | 0.364 | 5.427 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drogoudi, P.; Pantelidis, G.; Karapetsi, L.; Ziakou, K.; Kazantzis, K.; Madesis, P.; Thomidis, T. Exploring the Genetic and Morphological Variation and Disease Resistance in Local and Foreign Prunus persica (L.) Batsch Cultivars. Agriculture 2023, 13, 800. https://doi.org/10.3390/agriculture13040800
Drogoudi P, Pantelidis G, Karapetsi L, Ziakou K, Kazantzis K, Madesis P, Thomidis T. Exploring the Genetic and Morphological Variation and Disease Resistance in Local and Foreign Prunus persica (L.) Batsch Cultivars. Agriculture. 2023; 13(4):800. https://doi.org/10.3390/agriculture13040800
Chicago/Turabian StyleDrogoudi, Pavlina, Georgios Pantelidis, Lefkothea Karapetsi, Konstantina Ziakou, Konstantinos Kazantzis, Panagiotis Madesis, and Thomas Thomidis. 2023. "Exploring the Genetic and Morphological Variation and Disease Resistance in Local and Foreign Prunus persica (L.) Batsch Cultivars" Agriculture 13, no. 4: 800. https://doi.org/10.3390/agriculture13040800
APA StyleDrogoudi, P., Pantelidis, G., Karapetsi, L., Ziakou, K., Kazantzis, K., Madesis, P., & Thomidis, T. (2023). Exploring the Genetic and Morphological Variation and Disease Resistance in Local and Foreign Prunus persica (L.) Batsch Cultivars. Agriculture, 13(4), 800. https://doi.org/10.3390/agriculture13040800