
Variation in Response to Moisture Stress of Young Plants of Interspecific Hybrids between White Clover (T. repens L.) and Caucasian Clover (T. ambiguum M. Bieb.)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment 1
2.1.1. Plant Material and Experimental Treatments
2.1.2. Drought Tolerance
2.1.3. Plant Growth
2.2. Experiment 2
Root Depth Distribution
2.3. Data Analysis
3. Results
3.1. Experiment 1
3.1.1. Overall analysis

{kind=link}
| Treatment | LWP | Leaf RWC | SGR | LDR | DM Yield | Leaf Area | ||
|---|---|---|---|---|---|---|---|---|
| 21 Day | 35 Day | 21 Day | 35 Day | |||||
| Drought (D) | NS | *** | ** | * | * | * | ** | * |
| Population (P) | NS | *** | *** | *** | *** | *** | *** | *** |
| D × P | *** | *** | *** | *** | NS | NS | *** | *** |
3.1.2. Plant Water Status
| Population | Days after Start of Drought | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 21 | 35 | |||||||||
| C | M | S | C | M | S | |||||
| Caucasian Clover | −0.47 | −0.58 | −0.76 | −0.50 | −0.73 | −0.76 | ||||
| White Clover | −0.27 | −0.51 | −1.35 | −0.39 | −0.85 | −2.00 | ||||
| BC1 | −0.32 | −0.57 | −0.80 | −0.30 | −0.69 | −1.69 | ||||
| BC2 | −0.26 | −0.41 | −0.94 | −0.32 | −0.62 | −1.49 | ||||
| S.e.d. | ||||||||||
| Drought (D) | 0.270 NS | 0.037 *** | ||||||||
| Population (P) | 0.072 NS | 0.059 *** | ||||||||
| D × P | 0.291 *** (0.124 ***) | 0.092 *** (0.097 ***) | ||||||||
| Population | Days after Start of Drought | |||||
|---|---|---|---|---|---|---|
| 21 | 35 | |||||
| C | M | S | C | M | S | |
| Caucasian Clover | 93.1 | 94.3 | 93.1 | 92.2 | 92.9 | 92.0 |
| White Clover | 91.2 | 90.9 | 69.3 | 91.6 | 92.9 | 71.3 |
| BC1 | 94.1 | 93.2 | 76.5 | 92.7 | 92.5 | 76.4 |
| BC2 | 93.1 | 93.1 | 69.9 | 92.3 | 93.1 | 68.4 |
| S.e.d. | ||||||
| Drought (D) | 1.23 ** | 1.96 * | ||||
| Population (P) | 1.61 *** | 1.63 *** | ||||
| D × P | 2.72 *** (2.79 ***) | 3.13 *** (2.82 ***) | ||||
3.1.3. Plant Growth
| Population | Stolon Growth Rate | Leaf Development Rate | Leaf Area (mm2) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| C | M | S | C | M | S | C | M | S | |
| Caucasian Clover | - | - | - | - | - | - | 358.2 | 303.0 | 184.9 |
| White Clover | 3.8 | 4.0 | 1.1 | 9.9 | 10.0 | 5.6 | 280.4 | 241.9 | 178.6 |
| BC1 | 2.1 | 1.6 | 0.4 | 6.1 | 7.0 | 4.8 | 309.8 | 265.4 | 257.0 |
| BC2 | 3.1 | 2.3 | 0.6 | 6.4 | 7.0 | 4.3 | 322.2 | 248.3 | 220.8 |
| Drought (D) | 0.39 * | 0.38 * | 10.90 * | ||||||
| Population (P) | 0.32 *** | 0.43 *** | 10.19 *** | ||||||
| D × P | 0.65 NS (0.55 NS) | 0.73 NS (0.75 NS) | 18.77 *** (17.64 ***) | ||||||
3.2. Experiment 2
| Population | Moisture Level | ||
|---|---|---|---|
| C | M | S | |
| Caucasian Clover | 3.3 | 2.1 | 1.5 |
| White Clover | 30.8 | 20.2 | 4.4 |
| BC1 | 22.7 | 17.7 | 4.9 |
| BC2 | 26.9 | 16.1 | 4.4 |
| Drought (D) | 0.98 ** | ||
| Population (P) | 0.75 *** | ||
| P × S | 1.49 *** (1.30 ***) | ||
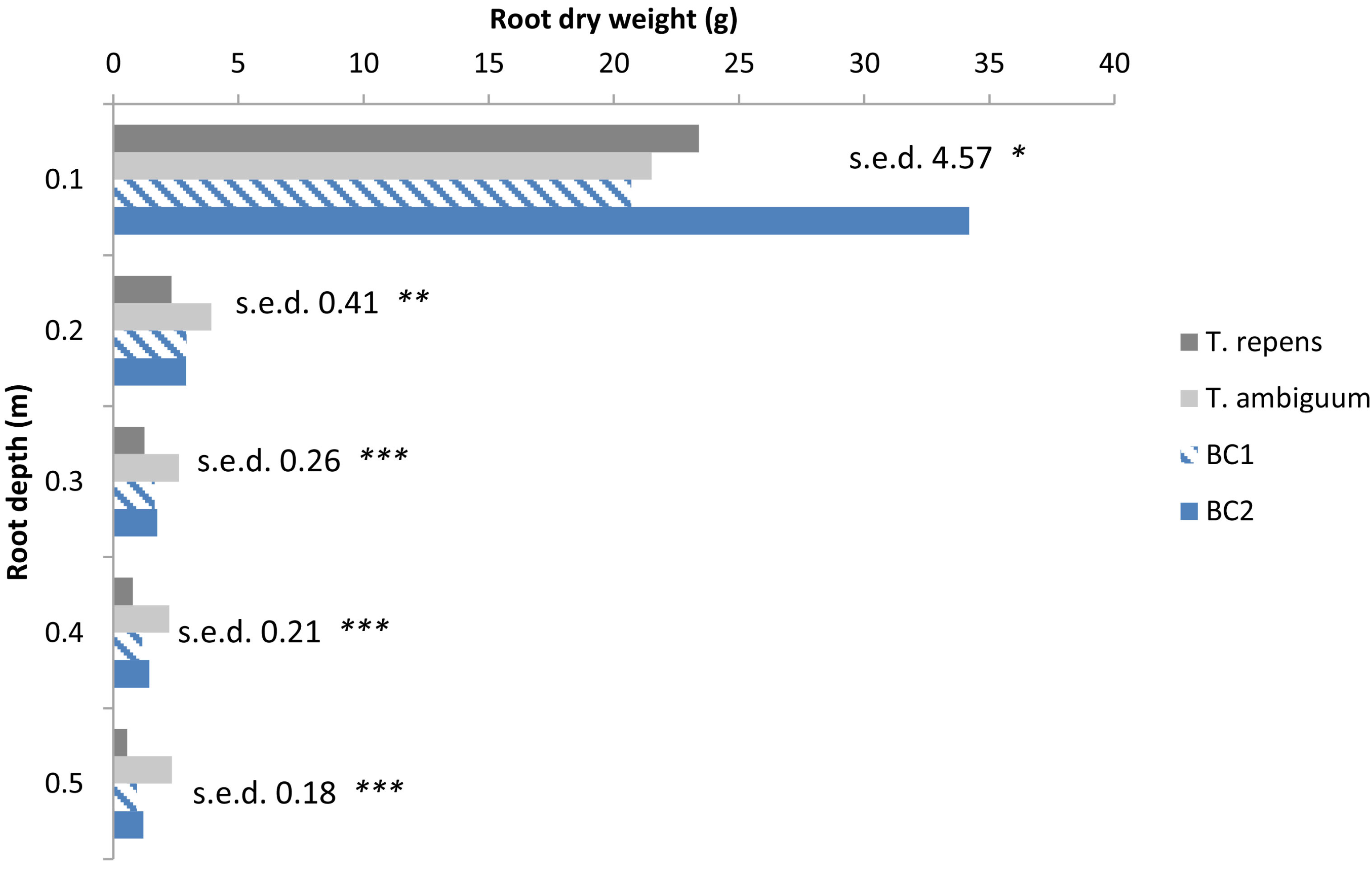
| Significance | Root Depth (m) | ||||
|---|---|---|---|---|---|
| 0.1 | 0.2 | 0.3 | 0.4 | 0.5 | |
| Population | * | ** | *** | *** | *** |
| Genotypes within Population | *** | ** | ** | ** | NS |

4. Discussion
4.1. Plant Water Status in Response to Drought
4.2. Root Depth Distribution
4.3. Plant Growth and Development
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Foresight. The Future of Food and Farming. Final Project Report; The Government Office for Science: London, UK, 2011.
- Hopkins, A.; Del Prado, A. Implications of climate change for grassland in Europe: Impacts, adaptations and mitigation options: A review. Grass Forage Sci. 2007, 62, 118–126. [Google Scholar] [CrossRef]
- Abberton, M.T.; MacDuff, J.H.; Marshall, A.H.; Humphreys, M.W. The genetic improvement offorage grasses and legumes to enhance adaptation of grasslands to climate change. In Proceedings of the United National Climate Change Conference, Nusa Dua, Indonesia, 3–14 December 2007.
- Foulkes, M.J.; Sylvester-Bradley, R.; Weightman, R.; Snape, J.W. Identifying physiological traitsassociated with improved drought resistance in winter wheat. Field Crops Res. 2007, 103, 11–14. [Google Scholar] [CrossRef]
- Dodd, I.C.; Whalley, W.R.; Ober, E.S.; Parry, M.A.J. Genetic and management approaches to boost UK winter wheat yields by ameliorating water deficits. J. Exp. Bot. 2011, 62, 5241–5248. [Google Scholar] [CrossRef] [PubMed]
- Lizana, C.; Wentworth, M.; Martinez, J.P.; Villegas, D.; Meneses, R.; Murchie, E.H.; Pastenes, C.; Lercari, B.; Vernieri, P.; Horton, P.; et al. Differential adaptation of two varieties of common bean to abiotic stress. I. Effects of drought on yield and photosynthesis. J. Exp. Bot. 2006, 57, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.P.; Silva, H.; Ledent, J.F.; Pinto, M. Effect of drought stress on the osmotic adjustment, cell wall elasticity and cell volume of six cultivars of common beans (Phaseolus vulgaris L.). Eur. J. Agron. 2007, 6, 30–38. [Google Scholar] [CrossRef]
- Abberton, M.T.; Michaelson-Yeates, T.P.T.; Marshall, A.H.; Holdbrook-Smith, K.; Rhodes, I. Morphological characteristics of hybrids between white clover, Trifolium. repens L. and Caucasian clover, Trifolium. ambiguum M. Bieb. Plant Breed. 1998, 117, 494–496. [Google Scholar] [CrossRef]
- Abberton, M.T.; Marshall, A.H. Progress in breeding perennial clovers for temperate agriculture. J. Agric. Sci. 2005, 143, 117–135. [Google Scholar] [CrossRef]
- Barbour, M.; Caradus, J.R.; Woodfield, D.R.; Silvester, W.B. Water stress and water use efficiency of ten white clover cultivars. In White Clover: New Zealand’s Competitive Edge; Woodfield, D.R., Ed.; Grassland Research and Practice Series No. 6; New Zealand Grassland Association: Palmerston North, New Zealand, 1996; pp. 159–162. [Google Scholar]
- Brink, G.E.; Pederson, G.A. White clover response to a water application gradient. Crop Sci. 1998, 38, 771–775. [Google Scholar] [CrossRef]
- Collins, R.P. The effect of drought stress and winter stress on the persistence of white clover. In Lowland Grasslands of Europe: Utilization and Development; Fisher, G., Frankow-Lindberg, B.E., Eds.; REUR Technical Series No. 64; FAO: Rome, Italy, 2002; pp. 17–32. [Google Scholar]
- Annicchiarico, P.; Piano, E. Indirect selection for root development of white clover and implications for drought tolerance. J. Agron. Crop Sci. 2004, 190, 28–34. [Google Scholar] [CrossRef]
- Caradus, J.R.; Woodfield, D.R. Genetic control of adaptive root characteristics in white clover. Plant Soil 1998, 200, 63–69. [Google Scholar] [CrossRef]
- Hussain, S.W.; Williams, W.M.; Mercer, C.F.; White, D.W.R. Transfer of clover cyst nematode resistance from Trifolium. nigrescens Viv. to T. repens by interspecific hybridisation. Theor. Appl. Genet. 1997, 95, 1274–1281. [Google Scholar] [CrossRef]
- Marshall, A.H.; Rascle, C.; Abberton, M.T.; Michaelson-Yeates, T.P.T.; Rhodes, I. Introgression as a route to improved drought tolerance in white clover (Trifolium. repens L.). J. Agron. Crop Sci. 2001, 187, 11–18. [Google Scholar] [CrossRef]
- Williams, W.M.; Hussain, S.W. Development of a breeding strategy for interspecific hybrids between Caucasian clover and white clover. NZ J. Agric. Res. 2008, 51, 115–126. [Google Scholar] [CrossRef]
- Williams, W.M. Trifolium interspecific hybridisation: Widening the white clover gene pool. Crop Pasture Sci. 2014, 65, 1091–1106. [Google Scholar] [CrossRef]
- Nichols, S.N.; Hofman, R.W.; Williams, W.M. Drought resistance of Trifolium repens × Trifolium uniflorum interspecific hybrids. Crop Pasture Sci. 2014, 65, 911–921. [Google Scholar] [CrossRef]
- Coolbear, P.; Hill, M.J.; Efendi, F. Relationships between vegetative and reproductive growth in a four year old stand of Caucasian clover (Trifolium. ambiguum M Bieb.) cv. Monaro. Proc. Agron. Soc. N. Z. 1994, 24, 77–82. [Google Scholar]
- Taylor, N.L.; Smith, R.R. Kura clover (Trifolium. ambiguum M.B.) breeding, culture and utilization. Adv. Agron. 1998, 63, 153–178. [Google Scholar]
- Black, A.D.; Moot, D.J.; Lucas, R.J. Development and growth characteristics of Caucasian and white clover seedlings, compared with perennial ryegrass. Grass Forage Sci. 2006, 61, 442–453. [Google Scholar] [CrossRef]
- Fu, S.M.; Hill, M.J.; Hampton, J.G. Root system development in Caucasian clover cv. Monaro and its contribution to seed yield. N. Z. J. Agric. Res. 2001, 44, 23–29. [Google Scholar] [CrossRef]
- Turner, L.B. The effect of water stress on the vegetative growth of white clover (T. repens L.): comparison of long-term water deficit and a short-term developing drought. J. Exp. Bot. 1991, 42, 311–316. [Google Scholar] [CrossRef]
- Belaygue, C.; Wery, J.; Cowan, A.A.; Tardieu, F. Contribution of leaf expansion, rate of leaf appearance and stolon branching to growth of plant leaf area under water deficit in white clover. Crop Sci. 1996, 36, 1240–1246. [Google Scholar] [CrossRef]
- Carlson, G.E. Growth of clover leaves developmental morphology at ten stages. Crop Sci. 1996, 6, 293–294. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Snyder, W.C. Nutrition of strawberry under controlled conditions: (a) Effects of deficiencies of boron and certain other elements: (b) Susceptibility to injury from sodium salts. Proc. Am. Soc. Hortic. Sci. 1933, 30, 288–296. [Google Scholar]
- Payne, R.W.; Murray, D.A.; Harding, S.A.; Baird, D.B.; Soutar, D.M. Introduction to GenStat® for WindowsTM; VSN International: Hemel Hempstead, UK, 2010. [Google Scholar]
- Humphreys, M.W.; Yadav, R.S.; Cairns, A.J.; Turner, L.B.; Humphreys, J.; Skøt, L. A changing climate for grassland research. New Phytol. 2006, 169, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Humphreys, M.O. Grass roots for improved soil structure and hydrology. IBERS Knowl.-Based Innov. 2011, 2011, 21–25. [Google Scholar]
- Humphreys, M.O. Genetic improvement of forage crops––Past, present and future. J. Agric. Sci. 2005, 143, 441–448. [Google Scholar] [CrossRef]
- Turner, N.C.; Begg, J.E. Responses of pasture plants to water deficits. In Plant Relations in Pastures; Wilson, J.R., Ed.; CSIRO: Melbourne, Australia, 1978; pp. 50–66. [Google Scholar]
- Widdup, K.H.; Knight, T.L.; Waters, C.J. Genetic variation for rate of establishment in Caucasian clover. Proc. Agron. Soc. N. Z. 1998, 60, 213–217. [Google Scholar]
- Marshall, A.H.; Williams, T.A.; Abberton, M.T.; Michaelson-Yeates, T.P.T.; Powell, H.G. Dry matter production of white clover (Trifolium. repens L.), Caucasian clover (T. ambiguum M. Bieb) and their associated hybrids when grown with a grass companion over three harvest years. Grass Forage Sci. 2003, 59, 91–99. [Google Scholar]
- Marshall, A.H.; Williams, T.A.; Abberton, M.T.; Michaelson-Yeates, T.P.T.; Olyott, P.; Powell, H.G. Forage quality of white clover (Trifolium. repens L.) × Caucasian clover (T. ambiguum M. Bieb) hybrids when grown with a grass companion over three harvest years. Grass Forage Sci. 2004, 59, 91–99. [Google Scholar] [CrossRef]
- Blum, A. Drought resistance, water-use efficiency, and yield potential––Are they compatible, dissonant or mutually exclusive? Aust. J. Agric. Res. 2005, 56, 1159–1168. [Google Scholar] [CrossRef]
- Sponchiado, B.N.; White, J.W.; Castillo, J.A.; Jones, P.G. Root growth of four common bean cultivars in relation to drought tolerance in environments with contrasting soil types. Exp. Agric. 1989, 25, 249–257. [Google Scholar] [CrossRef]
- Caradus , J.R. The structure and function of white clover root systems. Adv. Agron. 1990, 43, 1–46. [Google Scholar]
- Ennos, R.A. The significance of genetic variation for root growth within a natural population of white clover (Trifolium. repens L.). J. Ecol. 1985, 73, 615–624. [Google Scholar] [CrossRef]
- Clarke, J.M.; McCaig, T.N. Breeding for efficient root systems. In Plant breeding––Principles and Prospectsi; Hayward, M.D., Bosemark, N.O., Romagosa, I., Eds.; Chapman & Hal: London, UK, 1993; pp. 485–499. [Google Scholar]
- Wilson, D. Breeding for morphological and physiological traits. In Plant Breeding II; Frey, K.J, Ed.; Iowa State University Press: Ames, IA, USA, 1981; pp. 233–290. [Google Scholar]
- Thomas, H. Effects of drought on growth and competitive ability of perennial ryegrass and white clover. J. Appl. Ecol. 1984, 21, 591–602. [Google Scholar] [CrossRef]
- Stevenson, C.A.; Laidlaw, A.S. The effect of moisture stress on stolon and adventitious root development in white clover (Trifolium. repens L.). Plant Soil 1985, 85, 249–257. [Google Scholar] [CrossRef]
- Fouldes, W. Response to soil moisture supply in three leguminous species. I. Growth, reproduction and mortality. New Phytol. 1978, 80, 535–545. [Google Scholar] [CrossRef]
- Elgersma, A.; Fengrui, L. Effects of cultivar and cutting frequency on dynamics of stolon growth and leaf appearance rate in white clover grown in mixed swards. Grass Forage Sci. 1997, 3, 370–380. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marshall, A.H.; Lowe, M.; Collins, R.P. Variation in Response to Moisture Stress of Young Plants of Interspecific Hybrids between White Clover (T. repens L.) and Caucasian Clover (T. ambiguum M. Bieb.). Agriculture 2015, 5, 353-366. https://doi.org/10.3390/agriculture5020353
Marshall AH, Lowe M, Collins RP. Variation in Response to Moisture Stress of Young Plants of Interspecific Hybrids between White Clover (T. repens L.) and Caucasian Clover (T. ambiguum M. Bieb.). Agriculture. 2015; 5(2):353-366. https://doi.org/10.3390/agriculture5020353
Chicago/Turabian StyleMarshall, Athole H., Matthew Lowe, and Rosemary P. Collins. 2015. "Variation in Response to Moisture Stress of Young Plants of Interspecific Hybrids between White Clover (T. repens L.) and Caucasian Clover (T. ambiguum M. Bieb.)" Agriculture 5, no. 2: 353-366. https://doi.org/10.3390/agriculture5020353
APA StyleMarshall, A. H., Lowe, M., & Collins, R. P. (2015). Variation in Response to Moisture Stress of Young Plants of Interspecific Hybrids between White Clover (T. repens L.) and Caucasian Clover (T. ambiguum M. Bieb.). Agriculture, 5(2), 353-366. https://doi.org/10.3390/agriculture5020353