
A Modified Thermal Time Model Quantifying Germination Response to Temperature for C3 and C4 Species in Temperate Grassland

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Habitats
2.2. Germination Tests

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P | Species | Family | Life Cycle | Seed Weight (mg) |
|---|---|---|---|---|
| C4 | Kochia prostrata | Amaranthaceae | Annual | 0.762 ± 0.013 |
| Chenopodium album | Amaranthaceae | Annual | 0.579 ± 0.006 | |
| Salsola collina | Amaranthaceae | Annual | 1.632 ± 0.064 | |
| Amaranthus blitoides | Amaranthaceae | Annual | 0.965 ± 0.019 | |
| Amaranthus retroflexus | Amaranthaceae | Annual | 0.502 ± 0.006 | |
| Setaria viridis | Poaceae | Annual | 0.815 ± 0.007 | |
| Chloris virgata | Poaceae | Annual | 0.629 ± 0.025 | |
| Echinochloa crusgalli | Poaceae | Annual | 1.836 ± 0.028 | |
| Eriochloa villosa | Poaceae | Annual | 3.549 ± 0.353 | |
| Portulaca oleracea | Portulacaceae | Annual | 0.134 ± 0.003 | |
| Euphorbia humifusa | Euphorbiaceae | Annual | 0.434 ± 0.007 | |
| Artemisia anethifolia | Compositae | Biennial | 1.019 ± 0.012 | |
| C3 | Lappula echinata | Boraginaceae | Annual | 2.170 ± 0.052 |
| Incarvillea sinensis | Bignoniaceae | Annual | 0.660 ± 0.010 | |
| Dracocephalum moldavica | Labiatae | Annual | 1.892 ± 0.031 | |
| Bidens parviflora | Compositae | Annual | 5.530 ± 0.139 | |
| Saussurea glomerata | Compositae | Perennial | 2.843 ± 0.077 | |
| Lactuca indica | Compositae | Perennial | 1.031 ± 0.028 | |
| Achillea mongolica | Compositae | Perennial | 0.030 ± 0.001 | |
| Allium odorum | Liliaceae | Perennial | 2.187 ± 0.017 | |
| Convolvulus arvensis | Convolvulaceae | Perennial | 31.82 ± 0.131 | |
| Pharbitis purpurea | Convolvulaceae | Perennial | 28.55 ± 0.442 | |
| Cynanchum sibiricum | Asclepiadaceae | Perennial | 5.973 ± 0.124 | |
| Cynanchum chinense | Asclepiadaceae | Perennial | 4.217 ± 0.070 | |
| Potentilla chinensis | Rosaceae | Perennial | 0.411 ± 0.010 | |
| Stipa baicalensis | Poaceae | Perennial | 7.980 ± 0.194 | |
| Plantago asiatica | Plantaginaceae | Perennial | 0.229 ± 0.002 |
2.3. Data Analysis
3. Results and Discussion
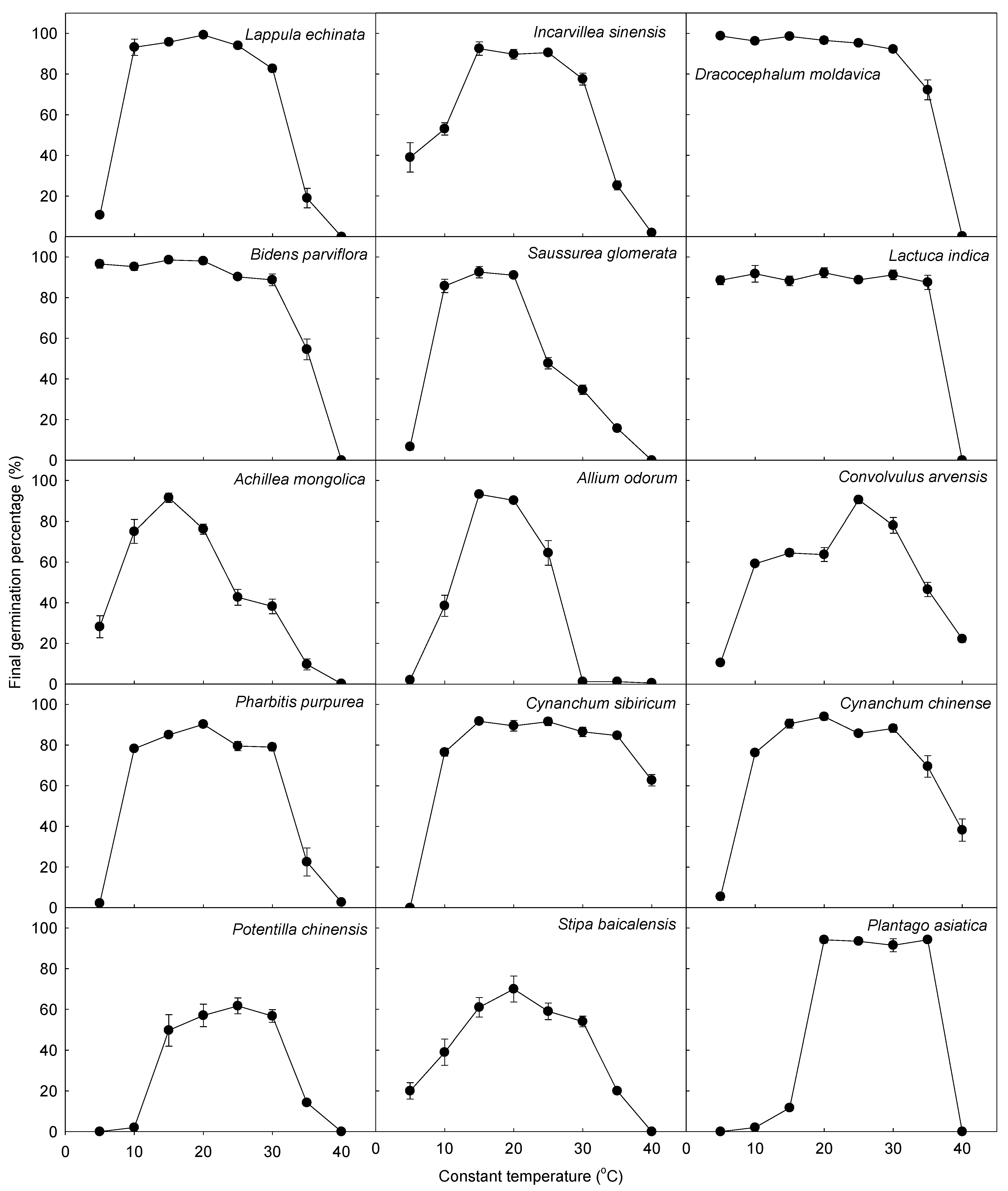
3.1. Germination Responses of C3 and C4 Species to Temperature




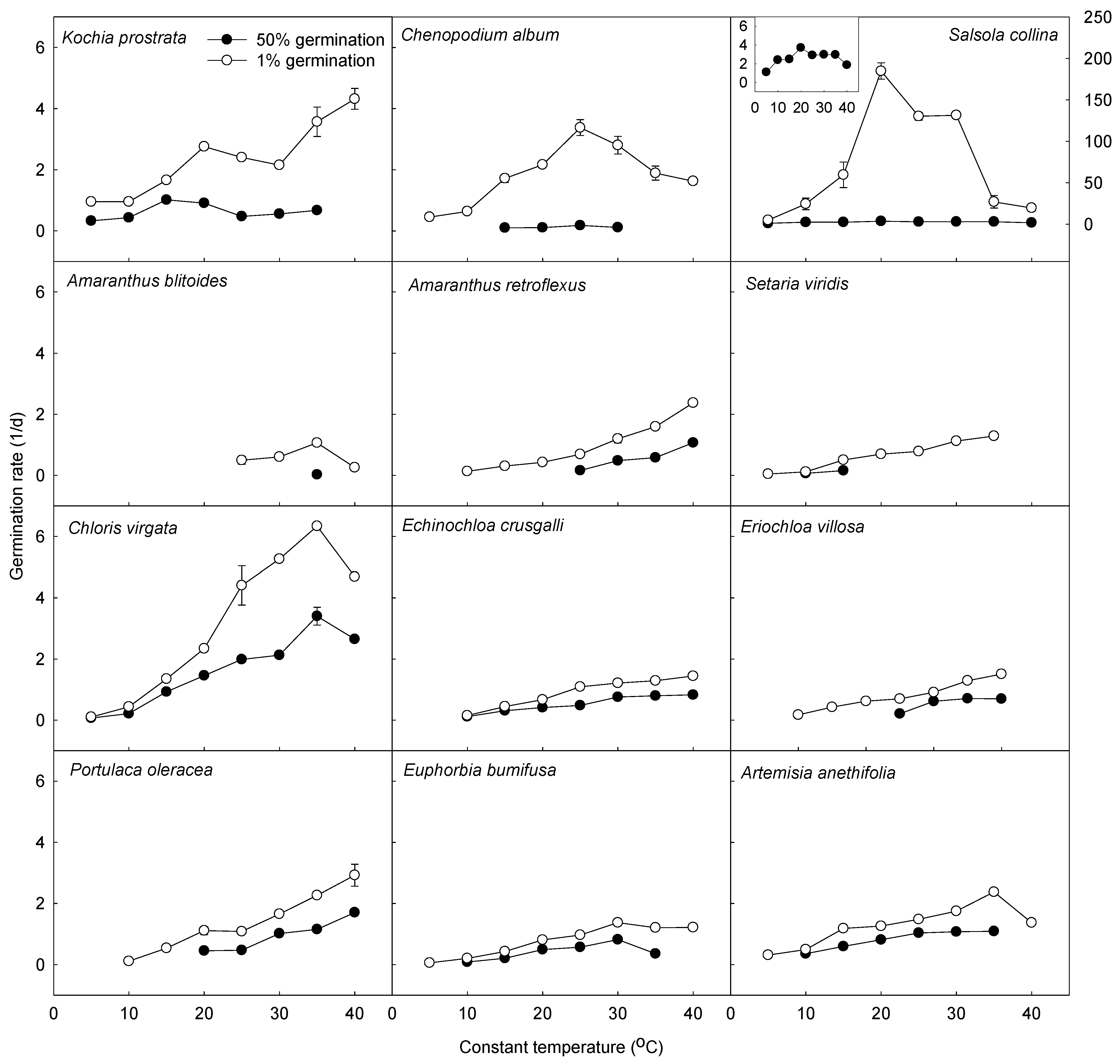
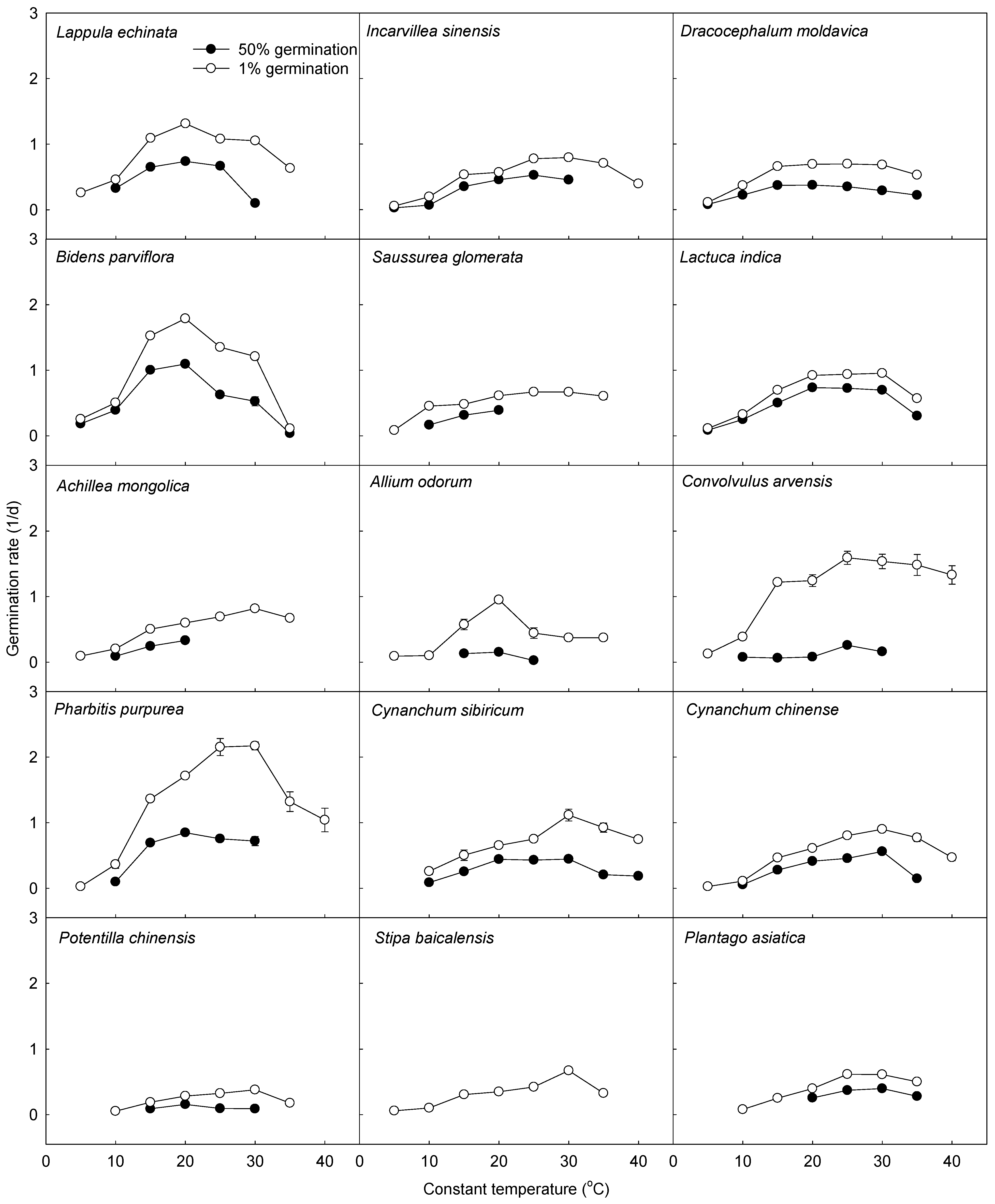
3.2. Comparison of Model Parameters between C3 and C4 Species
| Species | G | N | Tb (°C) | θ1 (°C·d) | R2 | p |
|---|---|---|---|---|---|---|
| Kochia prostrata | 1% | 8 | −3.6 | 11.1 | 0.85 | 0.001 |
| 50% | 3 | 1.4 | 14.6 | 0.86 | 0.24 | |
| Chenopodium album | 1% | 5 | 3.7 | 6.8 | 0.95 | 0.0044 |
| 50% | 3 | 1.7 | 126.6 | 0.81 | 0.28 | |
| Salsola collina | 1% | 4 | 6.5 | 0.09 | 0.84 | 0.08 |
| 50% | 4 | −2.8 | 6.3 | 0.92 | 0.04 | |
| Amaranthus blitoides | 1% | 3 | 17.3 | 17.6 | 0.89 | 0.21 |
| 50% | ||||||
| Amaranthus retroflexus | 1% | 7 | 11.6 | 13.9 | 0.92 | 0.0006 |
| 50% | 4 | 22.3 | 17.7 | 0.94 | 0.031 | |
| Setaria viridis | 1% | 7 | 4.8 | 23.3 | 0.98 | < 0.0001 |
| 50% | 2 | |||||
| Chloris virgata | 1% | 6 | 7.0 | 4.5 | 0.95 | 0.0008 |
| 50% | 7 | 6.3 | 9.4 | 0.96 | 0.0001 | |
| Echinochloa crusgalli | 1% | 7 | 4.4 | 22.9 | 0.95 | 0.0002 |
| 50% | 5 | 5.8 | 34.4 | 0.95 | 0.0042 | |
| Eriochloa villosa | 1% | 7 | 6.4 | 23.3 | 0.97 | < 0.0001 |
| 50% | 2 | |||||
| Portulaca oleracea | 1% | 7 | 9.4 | 11.3 | 0.96 | < 0.0001 |
| 50% | 5 | 14.9 | 15.7 | 0.93 | 0.008 | |
| Euphorbia humifusa | 1% | 6 | 5.4 | 18.9 | 0.98 | 0.0001 |
| 50% | 5 | 8.1 | 27.3 | 0.98 | 0.0016 | |
| Artemisia anethifolia | 1% | 7 | 0.3 | 15.6 | 0.96 | 0.0001 |
| 50% | 4 | 2.0 | 22.2 | 0.99 | 0.0002 |
| Species | G | N | Tb (°C) | θ1 (°C·d) | R2 | p |
|---|---|---|---|---|---|---|
| Lappula echinata | 1% | 4 | 2.2 | 13.2 | 0.95 | 0.0241 |
| 50% | 3 | 1.0 | 25 | 0.90 | 0.20 | |
| Incarvillea sinensis | 1% | 5 | 3.2 | 27.5 | 0.95 | 0.0041 |
| 50% | 5 | 4.6 | 36.1 | 0.94 | 0.0072 | |
| Dracocephalum moldavica | 1% | 3 | 3.1 | 18.2 | 0.99 | 0.0231 |
| 50% | 3 | 2.3 | 34.4 | 0.99 | 0.0045 | |
| Bidens parviflora | 1% | 4 | 3.4 | 8.9 | 0.93 | 0.0352 |
| 50% | 3 | 3.6 | 12.2 | 0.93 | 0.17 | |
| Saussurea glomerata | 1% | 4 | −0.2 | 31.1 | 0.84 | 0.08 |
| 50% | 3 | 2.0 | 44.6 | 0.96 | 0.11 | |
| Lactuca indica | 1% | 4 | 3.3 | 17.9 | 0.99 | 0.0057 |
| 50% | 4 | 3.6 | 22.7 | 0.99 | 0.0037 | |
| Achillea mongolica | 1% | 6 | 1.1 | 33.8 | 0.96 | 0.0006 |
| 50% | 3 | 5.7 | 41.8 | 0.98 | 0.09 | |
| Allium odorum | 1% | 4 | 5.5 | 16.4 | 0.90 | 0.0494 |
| 50% | 2 | |||||
| Convolvulus arvensis | 1% | 5 | 2.9 | 13.2 | 0.92 | 0.0099 |
| 50% | 4 | 6.7 | 89.3 | 0.61 | 0.21 | |
| Pharbitis purpurea | 1% | 5 | 5.0 | 8.9 | 0.97 | 0.0026 |
| 50% | 3 | 7.7 | 13.3 | 0.90 | 0.20 | |
| Cynanchum sibiricum | 1% | 5 | 3.2 | 25.5 | 0.96 | 0.0037 |
| 50% | 3 | 7.6 | 28.2 | 0.99 | 0.0196 | |
| Cynanchum chinense | 1% | 6 | 4.6 | 26.6 | 0.97 | 0.0004 |
| 50% | 5 | 5.1 | 42.0 | 0.93 | 0.0076 | |
| Potentilla chinensis | 1% | 5 | 4.3 | 63.3 | 0.94 | 0.0058 |
| 50% | 2 | |||||
| Stipa baicalensis | 1% | 6 | 3.8 | 43.1 | 0.94 | 0.0013 |
| 50% | ||||||
| Plantago asiatica | 1% | 4 | 7.9 | 28.6 | 0.96 | 0.0001 |
| 50% | 3 | 0.8 | 70.9 | 0.88 | 0.22 |
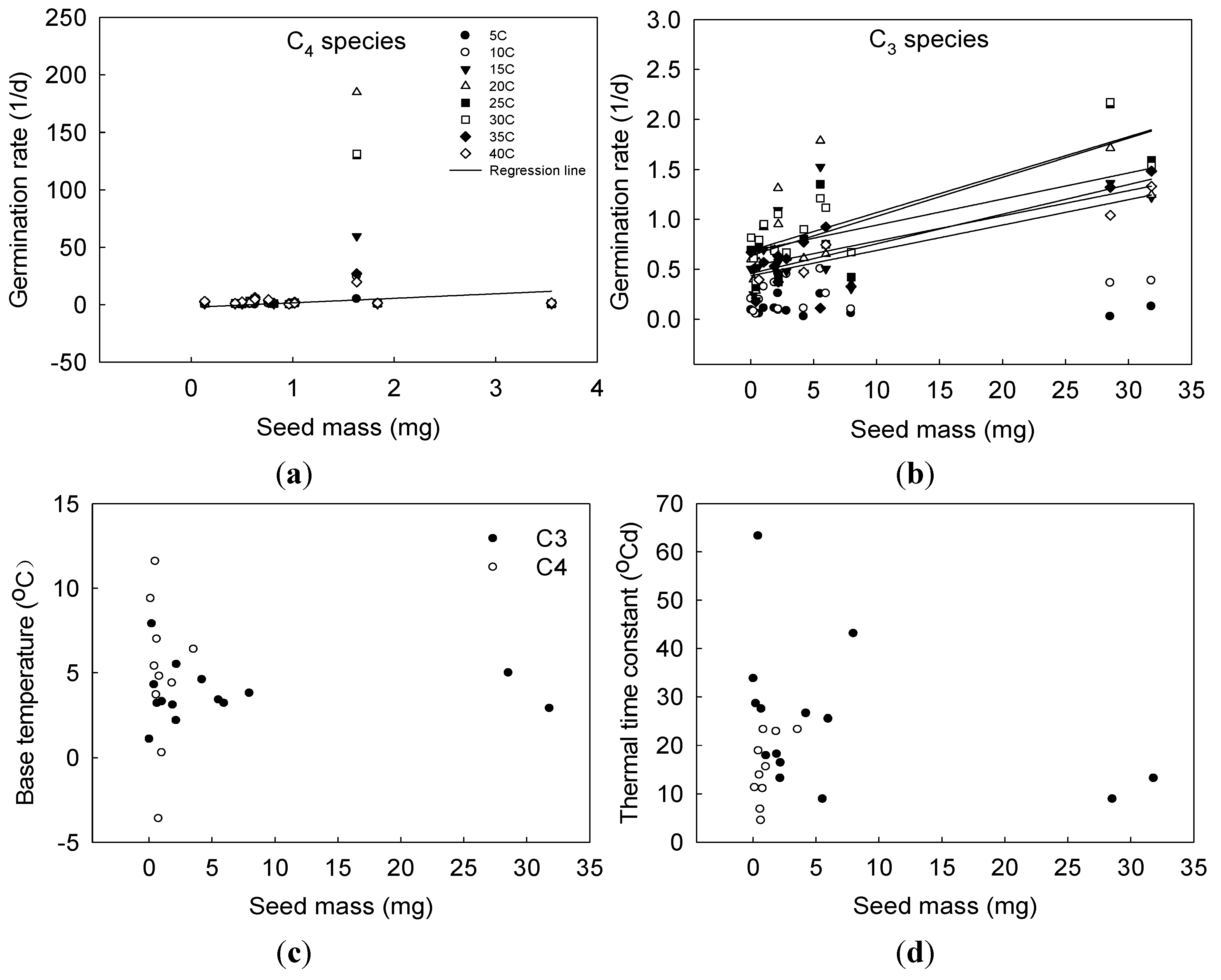
3.3. Relationship between Seed Mass and Germination Parameters

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Thompson, P.A. Characterisation of the germination response to temperature of species and ecotypes. Nature 1970, 225, 827–831. [Google Scholar] [CrossRef]
- Mott, J.J.; McKeon, G.M.; Moore, C.M. The effect of seed bed conditions on the germination of four Stylosanthes species in the Northern Territory. Aust. J. Agric. Res. 1976, 27, 811–823. [Google Scholar] [CrossRef]
- Alvarado, V.; Bradford, K.J. A hydrothermal time model explains the cardinal temperatures for seed germination. Plant Cell Environ. 2002, 25, 1061–1069. [Google Scholar] [CrossRef]
- Zhang, H.; McGill, C.R.; Irving, L.J.; Kemp, P.D.; Zhou, D. A modified thermal time model to predict germination rate of ryegrass and tall fescue at constant temperature. Crop Sci. 2013, 53, 240–249. [Google Scholar] [CrossRef]
- Mott, J.J.; Groves, R.H. Germination strategies. In The Biology of Australian Plants; Pate, J.S., McComb, A., Eds.; University of Western Australia Press: Perth, Australia, 1981; pp. 307–341. [Google Scholar]
- Murtagh, G.J. Effect of temperature on the germination of Glycine javanica. In Proceedings of the International Grassland Congress; Norman, M.J.T., Ed.; University of Queensland Press: Brisbane, Australia, 1970; pp. 574–578. [Google Scholar]
- Mayer, A.M.; Poljakoff-Mayber, A. The Germination of Seeds, 3rd ed.; Pergammon Press: Oxford, UK, 1982. [Google Scholar]
- Hegarty, T.W. Germination and other biological processes. Nature 1973, 243, 305–306. [Google Scholar] [CrossRef]
- Washitani, I.; Takenaka, A. Mathematical description of the seed germination dependency on time and temperature. Plant Cell Environ. 1984, 7, 359–362. [Google Scholar]
- Covell, S.; Ellis, R.H.; Roberts, E.H.; Summerfield, R.J. The influence of temperature on seed germination rate in grain legumes. 1. A comparison of chickpea, lentil, soyabean and cowpea at constant temperatures. J. Exp. Bot. 1986, 37, 705–715. [Google Scholar] [CrossRef]
- Garcia-Huidobro, J.; Monteith, J.L.; Squire, G.R. Time, temperature and germination of pearl millet (Pennisetum. typhoides S. and H.). 1. Constant temperature. J. Exp. Bot. 1982, 33, 288–296. [Google Scholar] [CrossRef]
- Trudgill, D.L.; Perry, J.N. Thermal time and ecological strategies––A unifying hypothesis. Ann. Appl. Biol. 1994, 125, 521–532. [Google Scholar] [CrossRef]
- Steinmaus, S.J.; Prather, T.S.; Holt, J.S. Estimation of base temperatures for nine weed species. J. Exp. Bot. 2000, 51, 275–286. [Google Scholar] [CrossRef]
- Ellis, R.H.; Covell, S.; Roberts, E.H.; Summerfield, R.J. The influence of temperature on seed germination rate in grain legumes. 2. Intraspecific variation in chickpea (Cicer. arietinum L.) at constant temperatures. J. Exp. Bot. 1986, 37, 1503–1515. [Google Scholar] [CrossRef]
- Larsen, S.U.; Bibby, B.M. Differences in thermal time requirement for germination of three turfgrass species. Crop. Sci. 2005, 45, 2030–2037. [Google Scholar] [CrossRef]
- Nori, H.; Moot, D.J.; Black, A.D. Thermal time requirements for germination of four annual clover species. N. Z. J. Agric. Res. 2014, 57, 30–37. [Google Scholar] [CrossRef]
- Trudgill, D.L.; Squire, G.R.; Thompson, K. A thermal time basis for comparing the germination requirements of some British herbaceous plants. New Phytol. 2000, 145, 107–114. [Google Scholar] [CrossRef]
- McDonald, C.K. Germination response to temperature in tropical and subtropical pasture legumes. 1. Constant temperature. Aust. J. Exp. Agric. 2002, 42, 407–419. [Google Scholar] [CrossRef]
- Murphy, B.P.; Bowman, D.M.J.S. Seasonal water availability predicts the relative abundance of C3 and C4 grasses in Australia. Global Ecol. Biogeogr. 2007, 16, 160–169. [Google Scholar] [CrossRef]
- Moles, A.T.; Ackerly, D.D.; Tweddle, J.C.; Dickie, J.B.; Smith, R.; Leishman, M.R.; Mayfield, M.M.; Pitman, A.; Wood, J.T.; Westoby, M. Global patterns in seed size. Global Ecol. Biogeogr. 2007, 16, 109–116. [Google Scholar] [CrossRef]
- Westoby, M.; Falster, D.S.; Moles, A.T.; Vest, P.A.; Wright, J.J. Plant ecological strategies: Some leading dimensions of variation between species. Ann. Rev. Ecol. Syst. 2002, 33, 125–159. [Google Scholar] [CrossRef]
- Foster, S.A.; Janson, C.H. The relationship between seed size and establishment conditions in tropical woody plants. Ecology 1985, 66, 773–780. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, G.; Lü, X.; Zhou, D.; Han, X. Salt tolerance during seed germination and early seedling stages of 12 halophytes. Plant Soil 2015, 388, 229–241. [Google Scholar] [CrossRef]
- Yin, L.; Li, M. A study on the geographic distribution and ecology of C4 plants in China. 1. C4 plant distribution in China and their relation with regional climatic condition. Acta Ecol. Sin. 1997, 17, 350–363. [Google Scholar]
- Tang, H.P.; Liu, S.R. The list of C4 plants in Nei Mongol area. Acta Sci. Nat. Univ. Nei Mong. 2001, 32, 431–438. [Google Scholar]
- Yin, L.; Zhu, L. C3 and C4 plants of forage resources in northeast grassland region of China. Inn. Mong. Pratac. 1990, 3, 32–40. [Google Scholar]
- Zhang, H.; Yu, Q.; Huang, Y.; Zheng, W.; Tian, Y.; Song, Y.; Li, G.; Zhou, D. Germination shifts of C3 and C4 species under simulated global warming scenario. PLoS ONE 2014, 9, e105139. [Google Scholar] [CrossRef]
- Trudgill, D.L. Why do tropical poikilothermic organisms tend to have higher threshold temperatures for development than temperate ones? Funct. Ecol. 1995, 9, 136–137. [Google Scholar]
- Grime, J.P.; Mason, G.; Curtis, A.V.; Rodman, J.; Band, S.R.; Mowforth, M.A.G.; Neal, A.M.; Shaw, S. A comparative study of germination characteristics in a local flora. J. Ecol. 1981, 64, 1017–1059. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Tian, Y.; Zhou, D. A Modified Thermal Time Model Quantifying Germination Response to Temperature for C3 and C4 Species in Temperate Grassland. Agriculture 2015, 5, 412-426. https://doi.org/10.3390/agriculture5030412
Zhang H, Tian Y, Zhou D. A Modified Thermal Time Model Quantifying Germination Response to Temperature for C3 and C4 Species in Temperate Grassland. Agriculture. 2015; 5(3):412-426. https://doi.org/10.3390/agriculture5030412
Chicago/Turabian StyleZhang, Hongxiang, Yu Tian, and Daowei Zhou. 2015. "A Modified Thermal Time Model Quantifying Germination Response to Temperature for C3 and C4 Species in Temperate Grassland" Agriculture 5, no. 3: 412-426. https://doi.org/10.3390/agriculture5030412
APA StyleZhang, H., Tian, Y., & Zhou, D. (2015). A Modified Thermal Time Model Quantifying Germination Response to Temperature for C3 and C4 Species in Temperate Grassland. Agriculture, 5(3), 412-426. https://doi.org/10.3390/agriculture5030412