
Temperature Impact on the Forage Quality of Two Wheat Cultivars with Contrasting Capacity to Accumulate Sugars

Abstract
:1. Introduction
2. Experimental Section
2.1. Plant Material
2.2. Experimental Layout
2.2.1. Field Enclosure Experiments

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean air temperature (°C) | DLI (mol photons m−2·day−1) | |||||
|---|---|---|---|---|---|---|
| Field Enclosure | Cool | C-W | Warm | Cool | C-W | Warm |
| 2005 | 8.5 ± 2.8 | – | 16.5 ± 1.7 | 12.7 ± 5.7 | – | 17.8 ± 7.8 |
| 2006 | 9.3 ± 3.2 | – | 15.7 ± 2.2 | 12.7 ± 3.4 | – | 11.1 ± 4.1 |
| 2008 | 8.7 ± 3.3 | 16.6 ± 2.3 | 16.3 ± 1.9 | 14.0 ± 5.3 | 12.2 ± 3.9 | 11.9 ± 3.6 |
2.2.2. Growth Chamber Experiments
2.3. Determinations
2.3.1. Plant Development
2.3.2. Relative Water Content
2.3.3. Dry matter content and sugar concentration
2.3.4. Cell Wall Components
2.3.5. Crude Protein
2.3.6. True in Vitro Dry Matter Digestibility (IVDMD)
2.3.7. In Vitro Rumen Gas Production
2.4. Experimental Design and Statistics
3. Results
3.1. Forage Composition
3.1.1. Forage Dry Matter Content
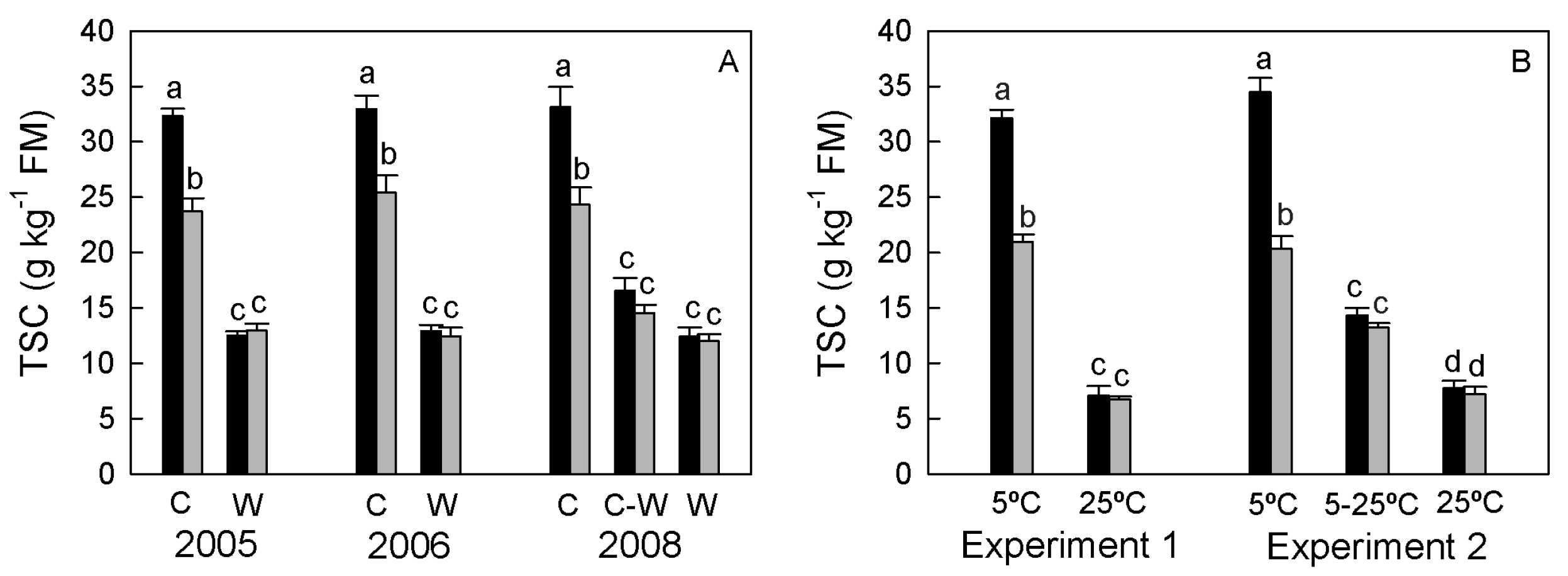
3.1.2. Total Sugar Concentration (TSC)
| Pincén | Patacón | |||||
|---|---|---|---|---|---|---|
| Field enclosure | Cool | Cool-Warm | Warm | Cool | Cool-Warm | Warm |
| 2005 | 179 ± 6.9 a | N.D. | 146 ± 7.3 c | 161 ± 8.3 b | N.D. | 140 ± 6.7 c |
| 2006 | 176 ± 7.1 a | N.D. | 141 ± 3.6 c | 159 ± 8.5 b | N.D. | 140 ± 6.0 c |
| 2008 | 175 ± 2.3 a | 162 ± 6.7 b | 136 ± 2.9 c | 159 ± 6.1 b | 155 ± 4.6 b | 134 ± 8.9 c |
| Growth Chamber | 5 °C | 5 °C–25 °C | 25 °C | 5 °C | 5 °C–25 °C | 25 °C |
| Experiment 1 | 189 ± 10.3 a | N.D. | 128 ± 7.3 c | 141 ± 8.7 b | N.D. | 123 ± 8.6 c |
| Experiment 2 | 183 ± 9.0 a | 156 ± 5.6 b | 125 ± 6.1 d | 142 ± 2.3 c | 133 ± 8.9 cd | 121 ± 8.9 d |

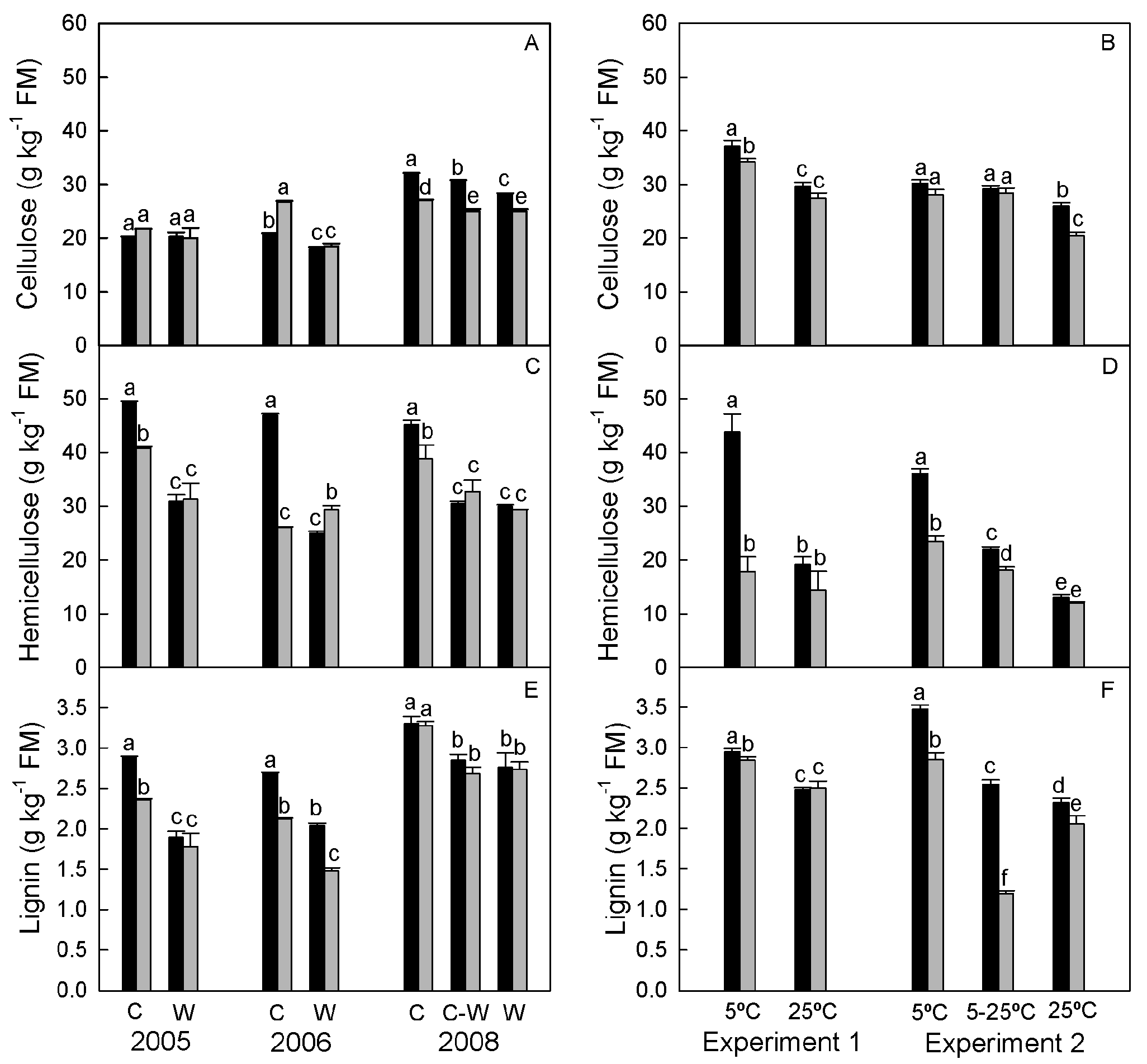
3.1.3. Structural Carbohydrates and Lignin

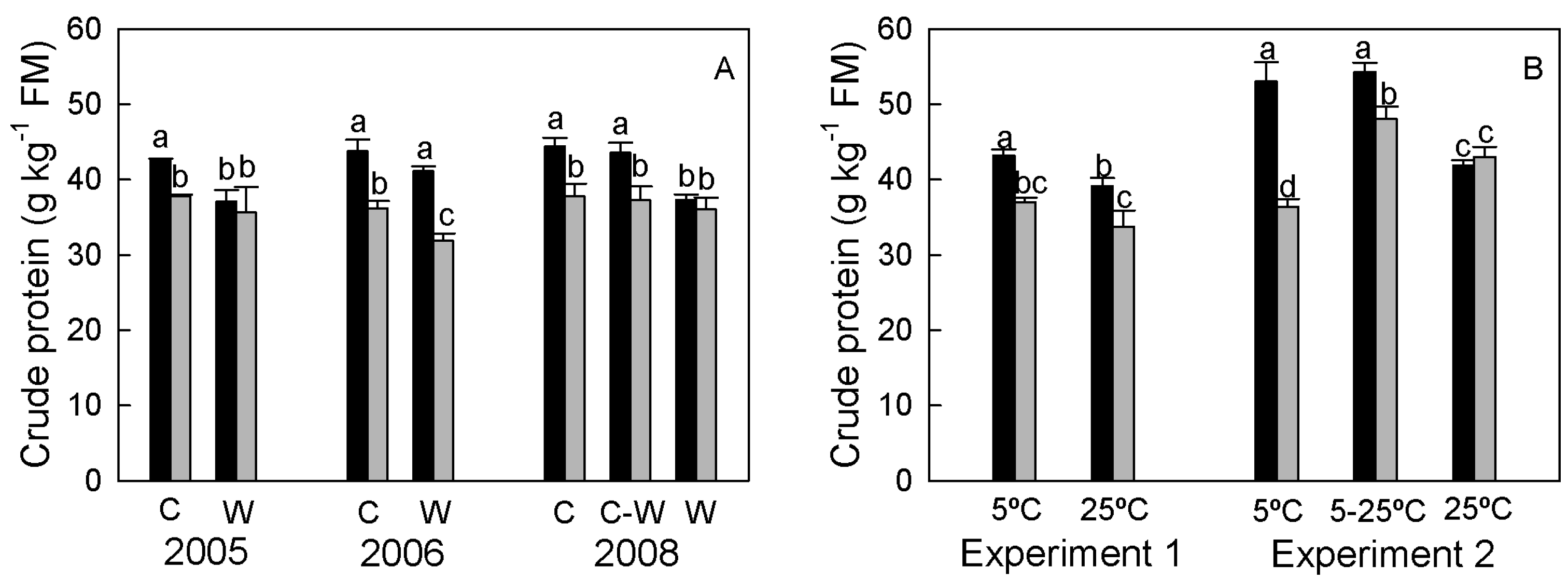
3.1.4. Crude Protein Concentration

3.2. Forage Quality
3.2.1. True in Vitro Dry Matter Digestibility (IVDMD)
| Pincén | Patacón | |||||
|---|---|---|---|---|---|---|
| Field Enclosure | Cool | Cool-Warm | Warm | Cool | Cool-Warm | Warm |
| 2005 | 913 ± 1.6 a | N.D. | 917 ± 13.4 a | 894 ± 4.9 a | N.D. | 912 ± 14.0 a |
| 2006 | 951 ± 33.2 a | N.D. | 868 ± 13.4 a | 942 ± 28.4 a | N.D. | 905 ± 23.4 a |
| 2008 | 754 ± 38.4 b | 783 ± 4.3 ab | 820 ± 3.9 ab | 842 ± 7.5 a | 830±2.6 ab | 838 ± 2.9 ab |
| Growth chamber | 5 °C | 5 °C–25 °C | 25 °C | 5 °C | 5 °C–25 °C | 25 °C |
| Experiment 1 | 941 ± 1.6 c | N.D. | 972 ± 2.1 a | 953 ± 1.8 bc | N.D. | 962 ± 3.4 ab |
| Experiment 2 | 975 ± 4.6 a | 963±1.4 ab | 936 ± 2.3 d | 954 ± 2.1 bc | 910±3.5 e | 946 ± 3.1 cd |
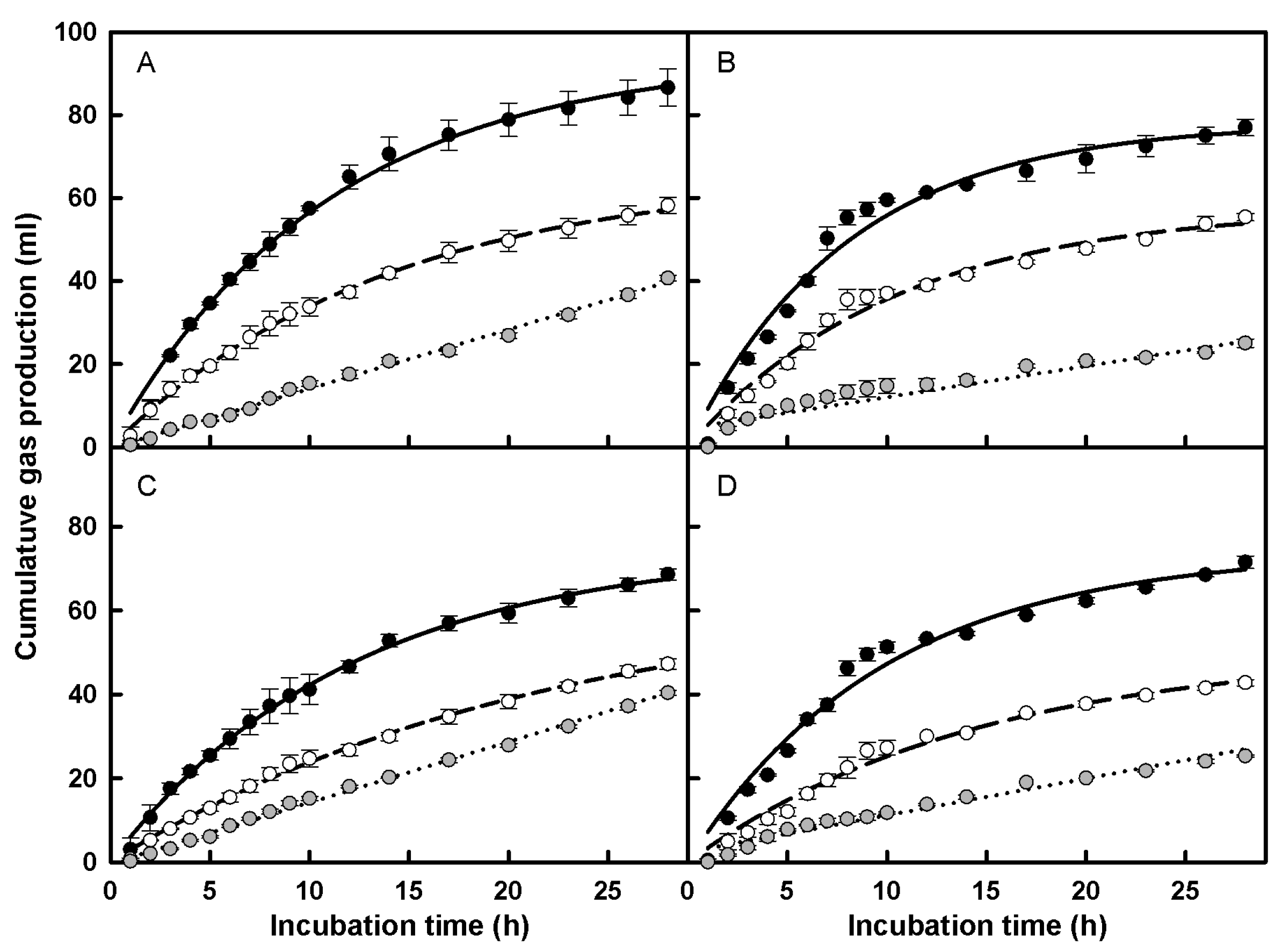
3.2.2. In Vitro Rumen Gas Production

4. Discussion
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hossaina, I.; Epplin, F.M.; Krenzer, E.G. Planting date influence on dual-purpose winter wheat forage yield, grain yield, and test weight. Agron. J. 2003, 95, 1179–1188. [Google Scholar] [CrossRef]
- Butchee, J.D.; Edwards, J.T. Dual-purpose wheat grain yield as affected by growth habit and simulated grazing intensity. Crop Sci. 2013, 53, 1686–1692. [Google Scholar] [CrossRef]
- Dove, H.; McMullen, G. Diet selection, herbage intake and liveweight gain in young sheep grazing dual-purpose wheats and sheep responses to mineral supplements. Anim. Prod. Sci. 2009, 49, 749–758. [Google Scholar] [CrossRef]
- Kelman, W.M.; Dove, H. Growth and phenology of winter wheat and oats in a dual-purpose management system. Crop Pasture Sci. 2009, 60, 921–932. [Google Scholar] [CrossRef]
- Tian, L.H.; Bell, L.W.; Shen, Y.Y.; Whish, J.P.M.; Nan, Z.B. Dual-purpose use of winter wheat in western China: Cutting time effects on forage production and grain yield. Crop Pasture Sci. 2012, 63, 520–528. [Google Scholar] [CrossRef]
- Arzadun, M.J.; Arroquy, J.I.; Laborde, H.E.; Brevedan, R.E. Grazing pressure on beef and grain production of dual-purpose wheat in Argentina. Agron. J. 2003, 95, 1157–1162. [Google Scholar] [CrossRef]
- Arzadun, M.J.; Arroquy, J.I.; Laborde, H.E.; Brevedan, R.E. Effect of planting date, clipping height, and cultivar on forage and grain yield of winter wheat in Argentinean Pampas. Agron. J. 2006, 98, 1274–1279. [Google Scholar] [CrossRef]
- Peralta, N.; Abbate, P.E.; Marino, A. Effect of the defoliation regime on grain production in dual purpose wheat. Agriscientia 2011, 28, 1–11. [Google Scholar]
- Morant, A.E.; Merchán, H.D.; Lutz, E.E. Comparación de la producción forrajera de cultivares de trigo para doble propósito. Rev. Argent. Prod. Anim. 1998, 18, 213–214. [Google Scholar]
- Lutz, E.E.; Merchán, H.D.; Morant, A.E. Carne y grano de un trigo doble propósito en condiciones semiá-ridas. Phyton (Buenos Aires) 2000, 67, 195–200. [Google Scholar]
- Lutz, E.E.; Merchán, H.D.; Morant, A.E. Estado de desarrollo de la planta de trigo (var. ProINTA Pincén) al momento de la última defoliación y su rendimiento en grano. Phyton (Buenos Aires) 2000, 68, 83–87. [Google Scholar]
- Bainotti, C.T.; Gomes, D.; Masiero, B.; Salines, J.; Fraschina, J.; Bertram, N.; Navarro, C. Evaluación de Cultivares de trigo como Doble Propósito. Available online: http://agrolluvia.com/wp-content/uploads/2010/05/INTA-Marcos-Ju%C3%A1rez-Evaluaci%C3%B3n-de-cultivares-de-trigo-como-doble-prop%C3%B3sito1.pdf (accessed on 18 May 2015).
- Morant, A.E.; Merchán, H.D.; Lutz, E.E. Evaluación de genotipos de trigos para doble propósito. Fecha de siembra y producción de grano. Rev. Argic. Prod. Anim. 2003, 23, 222–223. [Google Scholar]
- Lutz, E.; Merchán, H.; Morant, A. Mezcla de variedades de trigo para doble propósito. Phyton (Buenos Aires) 2008, 77, 217–223. [Google Scholar]
- Bell, L.W.; Moore, A.D. Mixed Crop-livestock Businesses Reduce Price- and Climate-induced Variability in Farm Returns: A Model-derived Case Study. Available online: http://aciar.gov.au/files/node/13992/mixed_crop_livestock_businesses_reduce_price_and__20972.pdf (accessed on 3 August 2015).
- Walker, D.W.; West, C.P.; Bacon, R.K.; Longer, D.E.; Turner, K.E. Changes in forage yield and composition of wheat and wheat-ryegrass mixtures with maturity. J. Dairy Sci. 1990, 73, 1296–1303. [Google Scholar] [CrossRef]
- Paterson, J.A.; Bowman, J.P.; Belyea, R.L.; Kerley, M.S.; Williams, J.E. The impact of forage quality and supplementation regimen on ruminant animal intake and performance. In Forage Quality, Evaluation, and Utilization; Fahey, G.C., Ed.; American Society of Agronomy, Crop Science Society of America, Soil Science Society of America: Madison, WI, USA, 1994; pp. 59–114. [Google Scholar]
- Kelman, W.M.; Dove, H.; Flint, P. The Potential of Winter Wheat Cultivars and Breeding Lines for Use in Dual-purpose (Grain and Graze) Systems. Available online: http://www.regional.org.au/au/asa/2006/poster/systems/4613_kelmanw.htm (accessed on 18 May 2015).
- Howarth, R.E.; Horn, G.W. Wheat pasture bloat of stocker cattle: A comparison with legume pasture bloat. In Proceedings of the National Wheat Pasture Symposium; Division of Agriculture, Oklahoma State University: Stillwater, OK, USA, 1984; pp. 24–25. [Google Scholar]
- Horn, G.W. Growing cattle on winter wheat pasture: Management and herd health considerations. Vet. Clin. North Am. Food A 2006, 22, 335–356. [Google Scholar] [CrossRef] [PubMed]
- Min, B.R.; Pinchak, W.E.; Mathews, D.; Fulford, J.D. In vitro rumen fermentation and in vivo bloat dynamics of steers grazing winter wheat to corn oil supplementation. Anim. Feed Sci. Technol. 2007, 133, 192–205. [Google Scholar] [CrossRef]
- Mayland, H.F.; Cheeke, P.R.; Majak, W.; Goff, J.P. Forage-induced animal disorders. In Forages, 6th ed.; Nelson, C.J., Moore, K.M., Collins, M., Eds.; Blackwell Publication: Ames, IA, USA, 2007; Volume 2, pp. 687–707. [Google Scholar]
- Malinowski, D.P.; Pitta, D.W.; Pinchak, W.E.; Min, B.R.; Emendack, Y.Y. Effect of nitrogen fertilisation on diurnal phenolic concentration and foam strength in forage of hard red wheat (Triticum aestivum L.) cv. Cutter. Crop Pasture Sci. 2011, 62, 656–665. [Google Scholar] [CrossRef]
- Min, B.R.; Pinchak, W.E.; Fulford, J.D.; Puchala, R. Wheat pasture bloat dynamics, in vitro ruminal gas production, and potential bloat mitigation with condensed tannins. J. Anim. Sci. 2005, 83, 1322–1331. [Google Scholar] [PubMed]
- Tognetti, J.A.; Calderón, P.L.; Pontis, H.G. Fructan metabolism: Reversal of cold acclimation. J. Plant Physiol. 1989, 134, 232–236. [Google Scholar] [CrossRef]
- Tognetti, J.A.; Salerno, C.L.; Crespi, M.D.; Pontis, H.G. Sucrose and fructan metabolism of different wheat cultivars at chilling temperatures. Physiol. Plant. 1990, 78, 554–559. [Google Scholar] [CrossRef]
- Equiza, M.A.; Miravé, J.P.; Tognetti, J.A. Differential root versus shoot growth inhibition and its relationship with carbohydrate accumulation at low temperature in different wheat cultivars. Ann. Bot. 1997, 80, 657–663. [Google Scholar] [CrossRef]
- Lorenzo, M.; Assuero, S.G.; Tognetti, J.A. Low temperature differentially affects tillering in spring and winter wheat in association with changes in plant carbon status. Ann. App. Biol. 2015, 166, 236–248. [Google Scholar] [CrossRef]
- Equiza, M.A.; Tognetti, J.A. Morphological plasticity of spring and winter wheats under changing temperatures. Funct. Plant Biol. 2002, 29, 1427–1436. [Google Scholar] [CrossRef]
- Blair, R.M.; Alcaniz, R.; Harrell, A. Shade intensity influences the nutrient quality and digestibility of southern deer browse leaves. J. Range Manag. 1983, 36, 257–264. [Google Scholar] [CrossRef]
- Moura, J.C.; Bonine, C.A.; de Oliveira Fernandes Viana, J.; Dornelas, M.C.; Mazzafera, P. Abiotic and biotic stresses and changes in the lignin content and composition in plants. J. Integr. Plant Biol. 2010, 52, 360–376. [Google Scholar] [CrossRef] [PubMed]
- Equiza, M.A.; Miravé, J.P.; Tognetti, J.A. Morphological, anatomical and physiological responses related to differential shoot vs. root growth inhibition at low temperature in spring and winter wheat. Ann. Bot. 2001, 87, 67–76. [Google Scholar] [CrossRef]
- Assuero, S.G.; Lorenzo, M.; Pérez, N.M.; Velázquez, L.; Tognetti, J.A. Tillering promotion by paclobutrazol in wheat and its relationship with plant carbohydrate status. N. Z. J. Agric. Res. 2012, 55, 347–358. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Calif. Agric. Exp. Stn. Circ. 1950, 347, 1–32. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Komareck, A.R.; Robertson, J.B.; van Soest, P.J. Comparison of the filter bag technique to conventional filtration in the Van Soest NDF analysis of 21 feeds. In Proceedings of the National Conference on Forage Quality, Evaluation and Utilization, Lincoln, NE, USA, 13–15 April 1994; Fahey, G.C., Ed.; Nebraska University: Lincoln, NE, USA, 1994. [Google Scholar]
- Komareck, A.R.; Robertson, J.B.; Van Soest, P.J. A comparison of methods for determining ADF using the filter bag technique versus conventional filtration. J Dairy Sci. 1993, 77, 24–26. [Google Scholar]
- Goering, H.K.; van Soest, P.J. Forage fiber analyses (Apparatus, Reagents, Procedures and Some Applications). In USDA-ARS Agricultural Handbook 379; US Government Printing Office: Washington, DC, USA, 1970; p. 20. [Google Scholar]
- Horneck, D.A.; Miller, R.O. Determination of total nitrogen in plant tissue. In Handbook of Reference Methods for Plant Analysis; Kalra, Y.P., Ed.; CRC Press: London, UK, 1998; pp. 75–83. [Google Scholar]
- ANKOM Tecnology. Analytical Methods in vitro True Digestibility Method (IVTD-Daisy). Available online: https://ankom.com/sites/default/files/document-files/Method_3_Invitro_0805_D200%2CD200I.pdf (accessed on 3 August 2015).
- Fay, J.P.; Cheng, K.-J.; Hanna, M.R.; Howarth, R.E.; Costerton, J.W. In vitro digestion of boat-safe and boat-causing legumes by rumen microorganisms: Gas and foam production. J Dairy Sci. 1980, 63, 1273–1281. [Google Scholar] [CrossRef]
- McDougall, E.I. Studies on ruminant saliva. 1. The composition and output of sheep’s saliva. Biochem. J. 1948, 43, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Tanino, K.; Weiser, C.J.; Fuchigami, L.H.; Chen, T.H. Water content during abscisic acid induced freezing tolerance in bromegrass cells. Plant Physiol. 1990, 93, 460–464. [Google Scholar] [CrossRef] [PubMed]
- Wanner, L.A.; Junttila, O. Cold-induced freezing tolerance in Arabidopsis. Plant Physiol. 1999, 120, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Ørskov, E.R.; McDonald, I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agric. Sci. 1979, 92, 499–503. [Google Scholar] [CrossRef]
- Pollock, C.J. The response of plants to temperature change. J. Agric. Sci. 1990, 115, 1–5. [Google Scholar] [CrossRef]
- Levitt, J. Responses of plants to environmental stress. In Chilling, Freezing, and High Temperature Stresses, 2nd ed.; Academic Press: New York, NY, USA, 1980; p. 447. [Google Scholar]
- Tarkowski, Ł.P.; van den Ende, W. Cold tolerance triggered by soluble sugars: A multifaceted countermeasure. Front. Plant Sci. 2015, 6, 203. [Google Scholar]
- Van den Ende, W. Multifunctional fructans and raffinose family oligosaccharides. Front. Plant Sci. 2013, 4, 247. [Google Scholar] [PubMed]
- Panelo, J.S.; Redi, W.I.; Lorenzo, M.; Tognetti, J. Efecto del Incremento de la Temperatura Sobre la Fotosíntesis y la Respiración en Plantas de Trigo Aclimatadas a Bajas Temperaturas. Available online: http://fisiologiavegetal.org/fv2014/abstract-index/abstracts/#905 (accessed on 21 September 2014).
- Huner, N.P.A.; Palta, J.P.; Li, P.H.; Carter, J.V. Anatomical changes in leaves of Puma rye in response to growth at cold-hardening temperatures. Bot. Gaz. 1981, 142, 55–62. [Google Scholar] [CrossRef]
- Huner, N.P.A.; Oquist, G.; Sarhan, F. Energy balance and acclimation to light and cold. Trends Plant Sci. 1998, 3, 224–230. [Google Scholar] [CrossRef]
- Ndong, C.; Danyluk, J.; Huner, N.P.; Sarhan, F. Survey of gene expression in winter rye during changes in growth temperature, irradiance or excitation pressure. Plant Mol. Biol. 2001, 45, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Gray, G.R.; Chauvin, L.P.; Sarhan, F.; Huner, N.P. Cold acclimation and freezing tolerance (A complex interaction of light and temperature). Plant Physiol. 1997, 114, 467–474. [Google Scholar] [PubMed]
- Getachew, G.; Robinson, P.H.; DePeters, E.J.; Taylor, S.J. Relationships between chemical composition, dry matter degradation and in vitro gas production of several ruminant feeds. Anim. Feed Sci. Technol. 2004, 111, 57–71. [Google Scholar] [CrossRef]
- Lopez-Guisa, J.M.; Satter, L.D. Effect of forage source on retention of digesta markers applied to corn gluten meal and brewers grains for heifers. J. Dairy Sci. 1991, 74, 4297–4304. [Google Scholar] [CrossRef]
- John, A.; Ulyatt, M.J. Importance of dry matter content to voluntary intake of fresh grass forages. Proc N. Z. Soc. Anim. Prod. 1987, 47, 13–16. [Google Scholar]
- Cabrera Estrada, J.I.; Delagarde, R.; Faverdin, P.; Peyraud, J.L. Dry matter intake and eating rate of grass by dairy cows is restricted by internal, but not external water. Anim. Feed Sci. Technol. 2004, 114, 59–74. [Google Scholar] [CrossRef]
- Allinson, D.W. Influence of photoperiod and thermoperiod on the IVDMD and cell wall components of tall fescue. Crop Sci. 1971, 11, 456–458. [Google Scholar] [CrossRef]
- Bertrand, A.; Tremblay, G.F.; Pelletier, S.; Castonguay, Y.; Bélanger, G. Yield and nutritive value of timothy as affected by temperature, photoperiod and time of harvest. Grass Forage Sci. 2008, 63, 421–432. [Google Scholar] [CrossRef]
- Thorvaldsson, G.; Tremblay, G.F.; Tapani Kunelius, H. The effects of growth temperature on digestibility and fibre concentration of seven temperate grass species. Acta Agric. Scand. Sect. B 2007, 57, 322–328. [Google Scholar] [CrossRef]
- Crasta, O.R.; Cox, W.J.; Cherney, J.H. Factors affecting maize forage quality development in the northeastern USA. Agron. J. 1997, 89, 251–256. [Google Scholar] [CrossRef]
- Agnusdei, M.G.; di Marco, O.N.; Nenning, F.R.; Aello, M.S. Leaf blade nutritional quality of rhodes grass (Chloris gayana) as affected by leaf age and length. Crop Pasture Sci. 2012, 62, 1098–1105. [Google Scholar] [CrossRef]
- Di Marco, O.N.; Harkes, H.; Agnusdei, M.G. Calidad de agropiro alargado (Thinopyrum ponticum) en estado vegetativo en relación con la edad y longitud de las hojas. RIA 2013, 39, 105–110. [Google Scholar]
- Reynolds, S.G. Pasture-Cattle-Coconut Systems; FAO RAPA Publication: Bangkok, Thailand, 1995; p. 668. [Google Scholar]
- Samarakoon, S.P.; Wilson, J.R.; Shelton, H.M. Growth, morphology and nutritive quality of shaded Stenotaphrum secundatum, Axonopus compressus and Pennisetum clandestinum. J Agric. Sci. 1990, 114, 161–169. [Google Scholar] [CrossRef]
- Livingston, D.P., III; Hincha, D.K.; Heyer, A.G.; Norio, S.; Noureddine, B.; Shuichi, O. The relationship of fructan to abiotic stress tolerance in plants. In Recent Advances in Fructooligosaccharides Research; Norio, S., Noureddine, B., Shuichi, O., Eds.; Research Signpost: Kerala, India, 2007; pp. 181–199. [Google Scholar]
- Valluru, R.; van den Ende, W. Plant fructans in stress environments: Emerging concepts and future prospects. J. Exp. Bot. 2008, 59, 2905–2916. [Google Scholar] [CrossRef] [PubMed]
- Livingston, D.P., III; Hincha, D.K.; Heyer, A.G. Fructan and its relationship to abiotic stress tolerance in plants. Cell Mol. Life Sci. 2009, 66, 2007–2023. [Google Scholar] [CrossRef] [PubMed]
- Vágújfalvi, A.; Kerepesi, I.; Galiba, G.; Tischner, T.; Sutka, J. Frost hardiness depending on carbohydrate changes during cold acclimation in wheat. Plant Sci. 1999, 144, 85–92. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lorenzo, M.; Assuero, S.G.; Tognetti, J.A. Temperature Impact on the Forage Quality of Two Wheat Cultivars with Contrasting Capacity to Accumulate Sugars. Agriculture 2015, 5, 649-667. https://doi.org/10.3390/agriculture5030649
Lorenzo M, Assuero SG, Tognetti JA. Temperature Impact on the Forage Quality of Two Wheat Cultivars with Contrasting Capacity to Accumulate Sugars. Agriculture. 2015; 5(3):649-667. https://doi.org/10.3390/agriculture5030649
Chicago/Turabian StyleLorenzo, Máximo, Silvia G. Assuero, and Jorge A. Tognetti. 2015. "Temperature Impact on the Forage Quality of Two Wheat Cultivars with Contrasting Capacity to Accumulate Sugars" Agriculture 5, no. 3: 649-667. https://doi.org/10.3390/agriculture5030649
APA StyleLorenzo, M., Assuero, S. G., & Tognetti, J. A. (2015). Temperature Impact on the Forage Quality of Two Wheat Cultivars with Contrasting Capacity to Accumulate Sugars. Agriculture, 5(3), 649-667. https://doi.org/10.3390/agriculture5030649