
Leaf Length Variation in Perennial Forage Grasses



Abstract
:1. Introduction


2. Genetic Variation in Leaf Length
2.1. Heritability

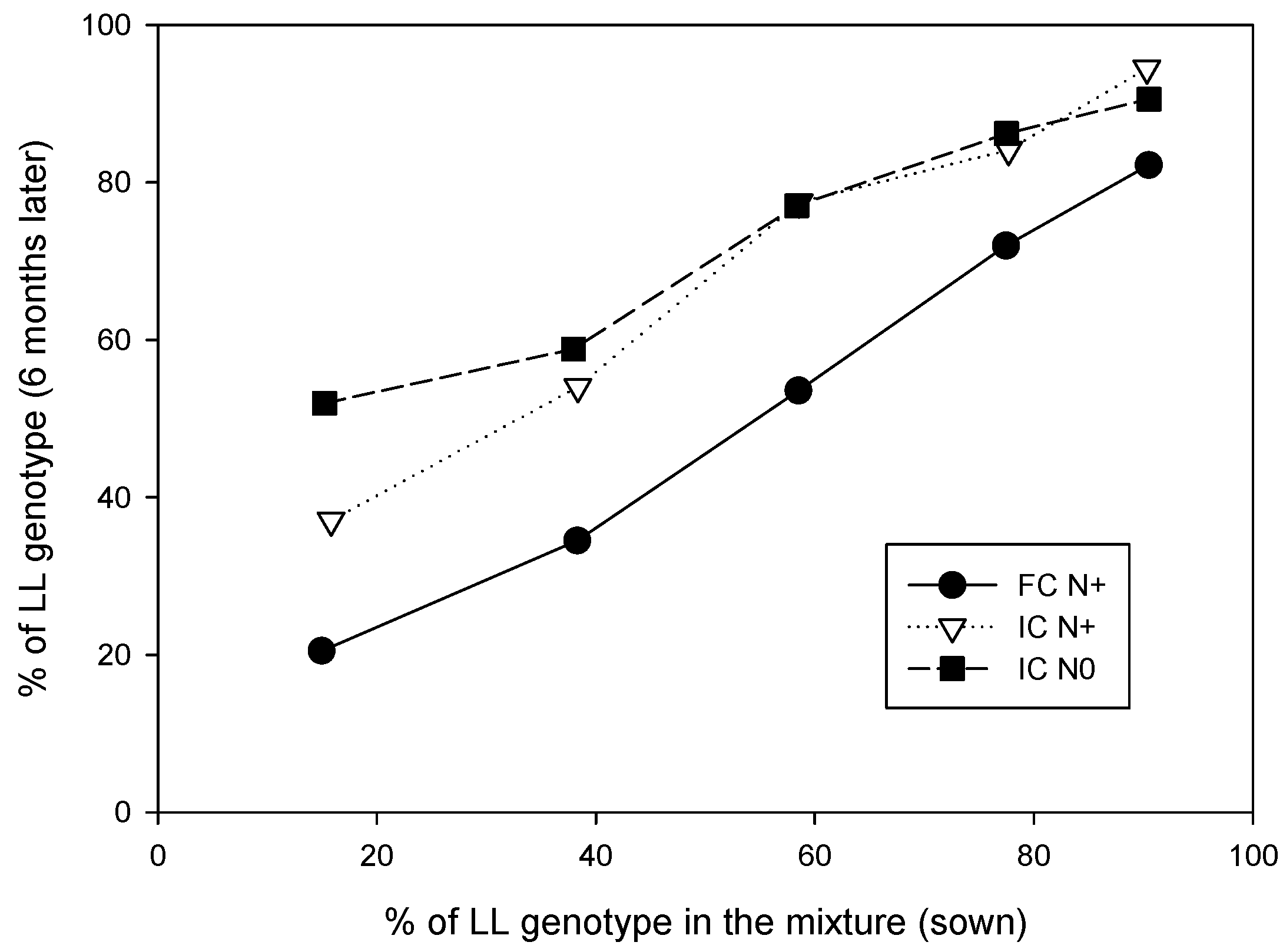
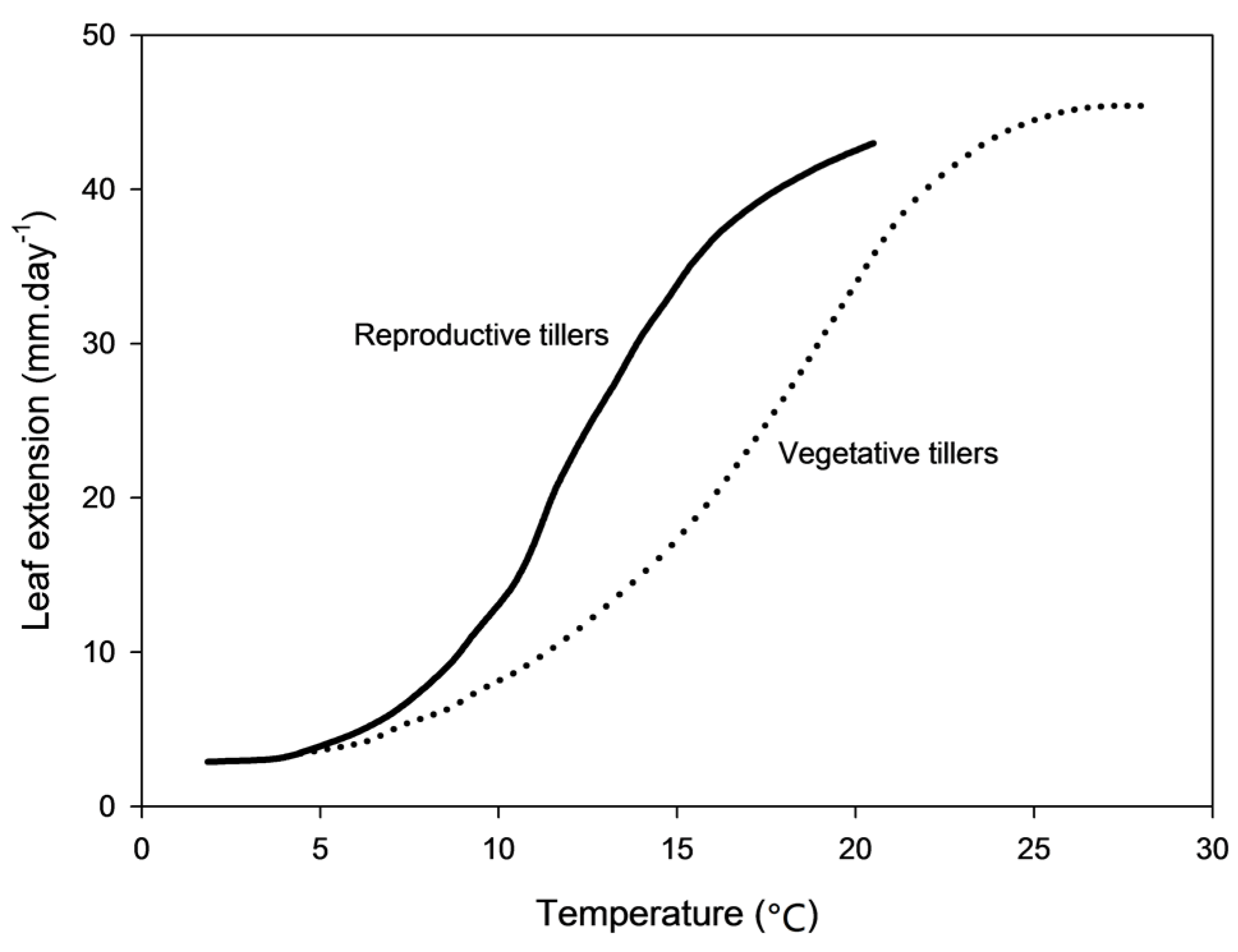
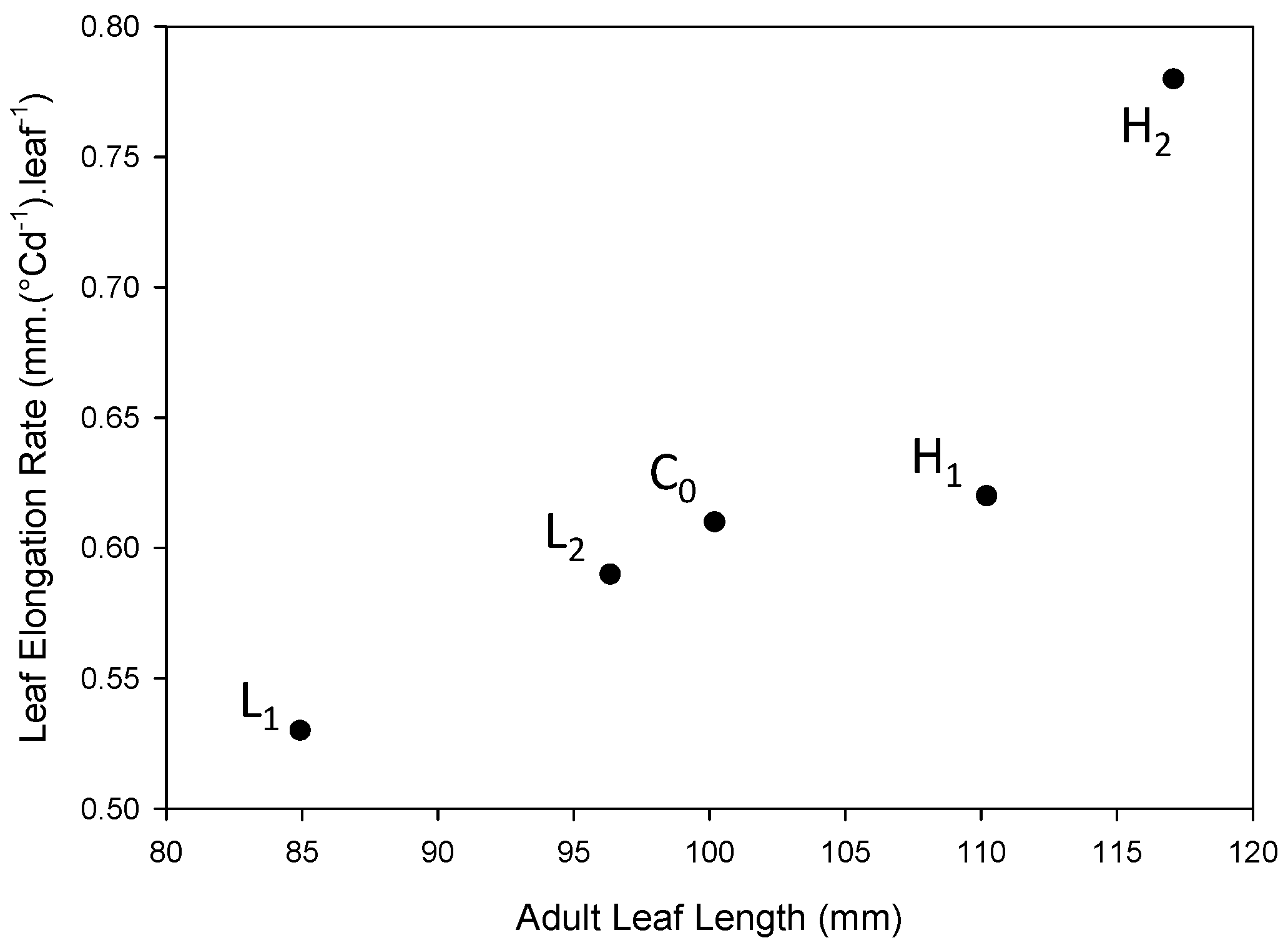
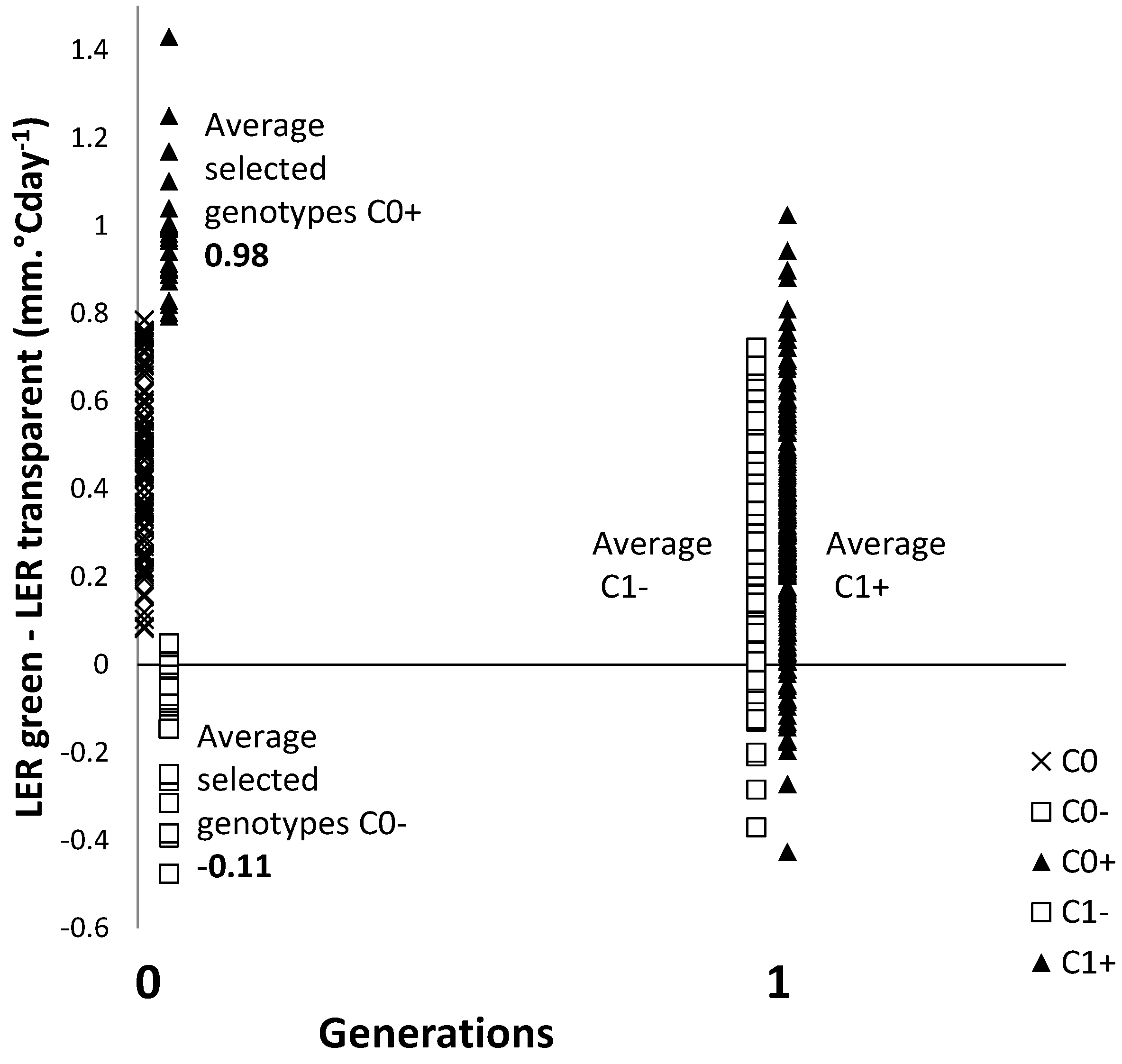
2.2. Genetic Architecture

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ref. | Parents | Traits | LG1 | LG2 | LG3 | LG4 | LG5 | LG6 | LG7 |
|---|---|---|---|---|---|---|---|---|---|
| [39] | Pop8490 | Leaf length | 13 | ||||||
| [41] | WSC F2 Perma × Aurora | 12 | 11 | 9 | 10 | 25 | 31–38 | 15 | |
| [54] | ILGI p152/112 mapping family | 6 | |||||||
| [42] | North African × Aurora F1 (NAx × AU6) | Leaf area | 12 | 6 | |||||
| [39] | Pop8490 (FL42 × FC61) | Lamina length in spring | 9 | 9 | 16 | ||||
| [39] | Pop8490 (FL42 × FC61) | Lamina length in autumn | 8–18 | 10 | 12 | 9 | |||
| [40] | Grasslands Impact × Grasslands Samson | Lamina length in autumn | 13 | 5 | 14–10 | 14–14 | 6 | ||
| [43] | WSC F2 Perma × Aurora | Leaf extension rate (LER) | 14 | 11 | |||||
| [40] | Grasslands Impact × Grasslands Samson | LER in spring | 13 | ||||||
| [39] | Pop8490 | LER in autumn | 11 | ||||||
| [40] | Grasslands Impact × Grasslands Samson | LER in autumn | 9 | 26–27 | 5–8 | ||||
| [55] | Three connected populations (elite material) | Vegetative plant height in spring | 6–5 | 5 | 4–4 | 4–4 | 4–9 | ||
| [55] | Three connected populations (elite material) | Vegetative plant height in autumn | 3 | 7–6 | 4 | 4 | |||
| [56] | WSC F2 Perma × Aurora | Flag lamina length * | 10 | ||||||
| [56] | ILGI p152/112 mapping family | 20 | |||||||
| [57] | Italian Veyo × Danish Falster | 11 | 13–12 | 17 |
3. Environmental and Genetic × Environmental Interaction Effects on Leaf Length

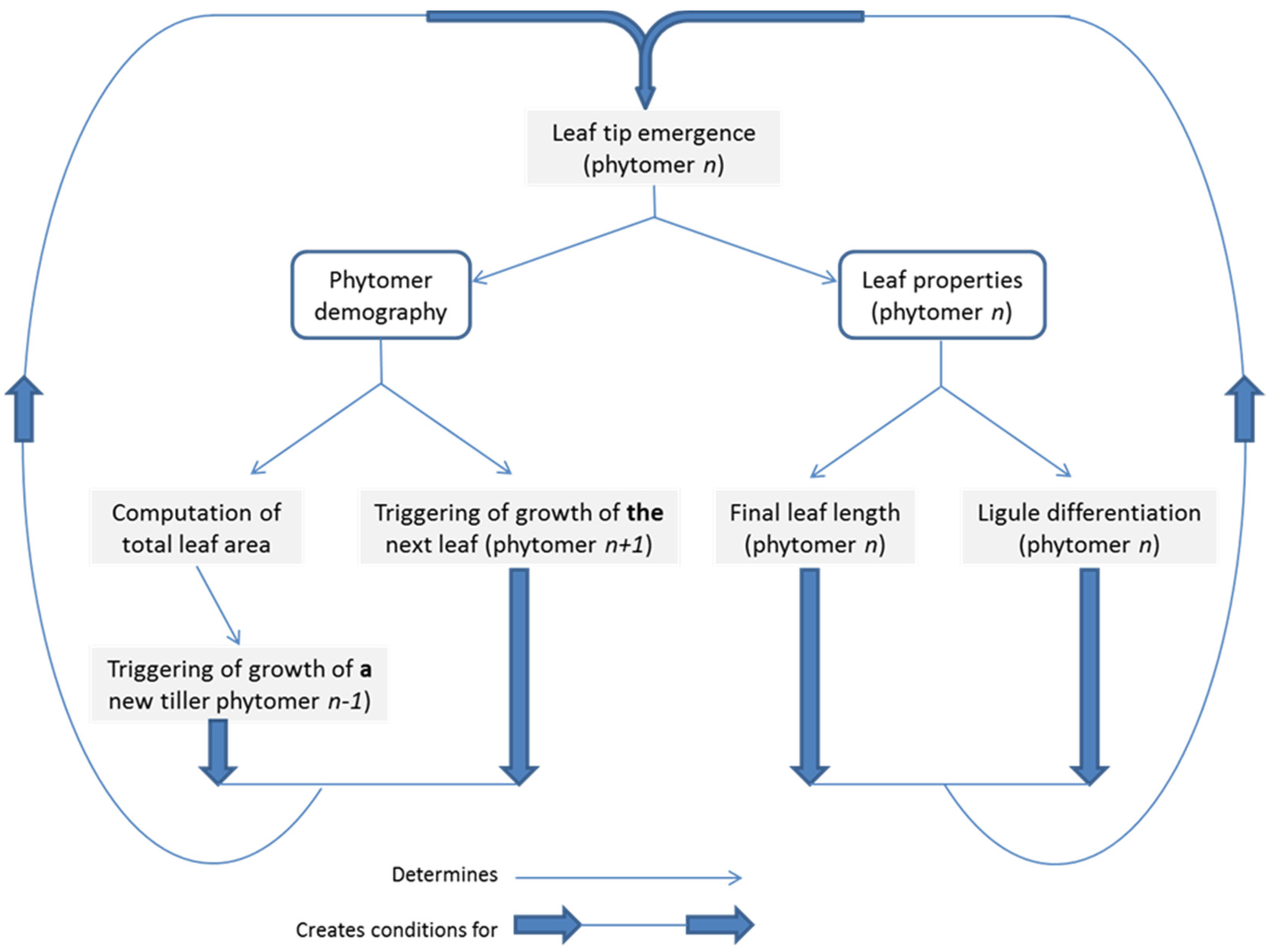
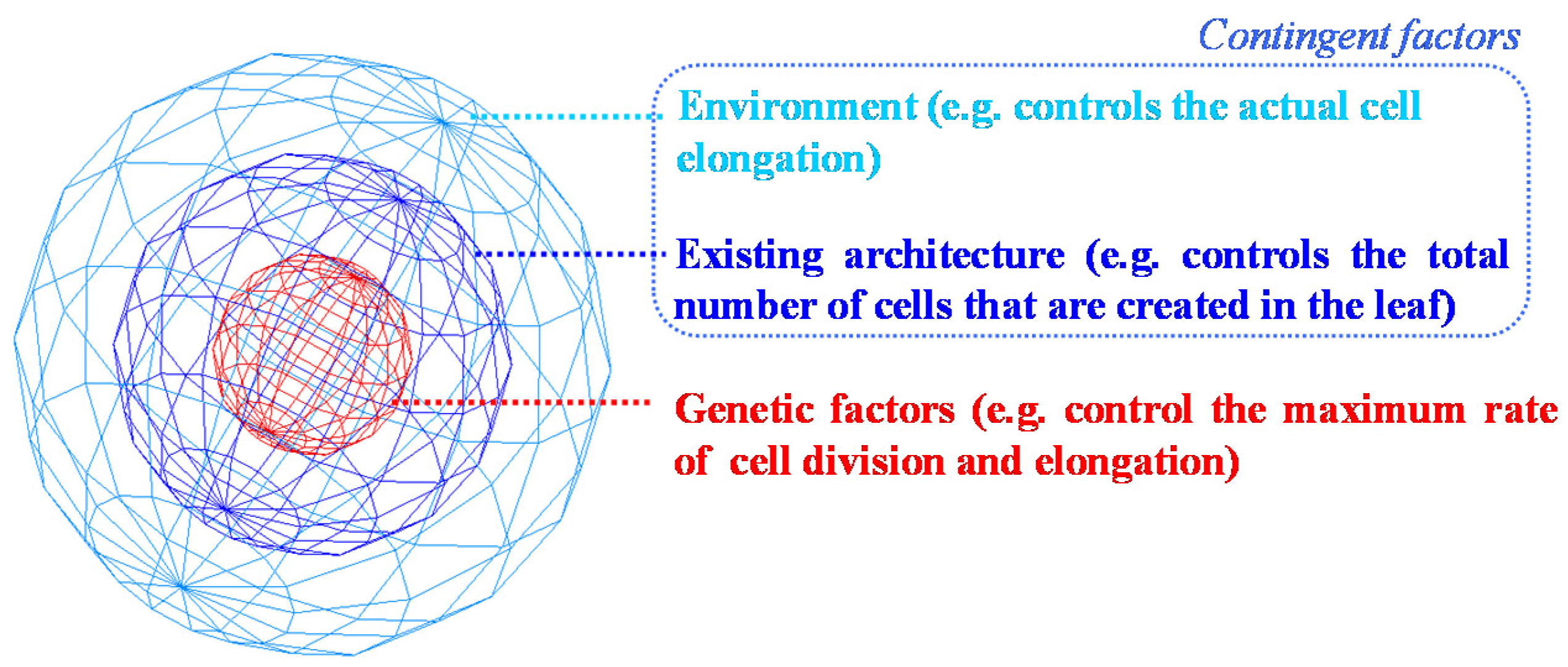
4. The Added Value of Plant Modelling


5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wilkins, P.W. Breeding perennial ryegrass for agriculture. Euphytica 1991, 52, 201–204. [Google Scholar] [CrossRef]
- Humphreys, M.O. Genetic improvement of forage crops—Past, present and future. Can. J. Agr. Sci. 2005, 143, 441–448. [Google Scholar] [CrossRef]
- Zarrough, K.M.; Nelson, C.J.; Coutts, J.H. Relationship between tillering and forage yield of tall fescue. I. Yield. Crop Sci. 1983, 23, 333–337. [Google Scholar] [CrossRef]
- Horst, G.L.; Nelson, C.J.; Asay, K.H. Relationship of leaf elongation to forage yield of tall fescue genotypes. Crop Sci. 1978, 18, 715–719. [Google Scholar] [CrossRef]
- Rhodes, I. The relationship between productivity and some components of canopy structure in ryegrass (Lolium spp.). I. Leaf length. Can. J. Agr. Sci. 1969, 73, 315–319. [Google Scholar] [CrossRef]
- Hazard, L.; Ghesquière, M. Productivity under contrasting cutting regimes of perennial ryegrass selected for short and long leaves. Euphytica 1997, 95, 295–299. [Google Scholar] [CrossRef]
- Zarrough, K.M.; Nelson, C.J.; Coutts, J.H. Relationship between tillering and forage yield of tall fescue. II. Pattern of tillering. Crop Sci. 1983, 23, 338–342. [Google Scholar] [CrossRef]
- Ceccarelli, S.; Falcinelli, M.; Damiani, F. Selection for dry matter yield in Lolium perenne L. II. Correlated responses under two cutting regimes. Can. J. Plant. Sci. 1980, 60, 501–508. [Google Scholar] [CrossRef]
- Ceccarelli, S.; Falcinelli, M.; Damiani, F. Selection for dry matter yield in Lolium perenne L. I. Direct response to selection. Can. J. Plant. Sci. 1980, 60, 491–500. [Google Scholar] [CrossRef]
- Hayward, M.D.; Vivero, J.L. Selection for yield in Lolium perenne. II. Performance of spaced plant selections under competitive conditions. Euphytica 1984, 33, 787–800. [Google Scholar] [CrossRef]
- Hazard, L.; Ghesquière, M.; Betin, M. Breeding for management adaptation in perennial ryegrass (Lolium perenne L.). I. Assessment of yield under contrasting cutting frequencies and relationships with leaf morphogenesis components. Agronomie 1994, 14, 259–266. [Google Scholar] [CrossRef]
- Hazard, L. Plasticity gives a greater flexibility to forage grass use. Fourrages 1996, 147, 293–302. [Google Scholar]
- Gastal, F.; Dawson, L.A.; Thornton, B. Responses of plant traits of four grasses from contrasting habitats to defoliation and n supply. Nutr. Cycling Agroecosyst. 2010, 88, 245–258. [Google Scholar] [CrossRef]
- Jones, R.J.; Nelson, C.J.; Sleper, D.A. Seedling selection for morphological characters associated with yield of tall fescue. Crop Sci. 1979, 19, 631–634. [Google Scholar] [CrossRef]
- Hazard, L.; Ghesquière, M.; Barraux, C. Genetic variability for leaf development in perennial ryegrass populations. Can. J. Plant. Sci. 1996, 76, 113–118. [Google Scholar] [CrossRef]
- Rhodes, I.; Mee, S. Changes in dry matter yield associated with selection for canopy characters in ryegrass. Grass Forage Sci. 1980, 35, 35–39. [Google Scholar] [CrossRef]
- Rhodes, I. The relationship between productivity and some components of canopy structure in ryegrass (Lolium spp.). III. Spaced plant characters, their heritabilities and relationship to sward yield. Can. J. Agr. Sci. 1973, 80, 171–176. [Google Scholar] [CrossRef]
- Barrett, P.D.; Laidlaw, A.S.; Mayne, C.S.; Christie, H. Pattern of herbage intake rate and bite dimensions of rotationally grazed dairy cows as sward height declines. Grass Forage Sci. 2001, 56, 362–373. [Google Scholar] [CrossRef]
- Barre, P.; Emile, J.C.; Betin, M.; Surault, F.; Ghesquière, M.; Hazard, L. Morphological characteristics of perennial ryegrass leaves that influence short-term intake in dairy cows. Agron. J. 2006, 98, 978–985. [Google Scholar] [CrossRef]
- Barre, P.; Gueye, B.; Gastal, F. Effect of light quality and quantity on leaf growth in Lolium perenne L. In Sustainable Use of Genetic Diversity in Forage and Turf breeding, Proceedings of the EUCARPIA 2009, La Rochelle, France, 11–14 May 2009; Huyghe, C., Ed.; Springer: Berlin, Germany, 2010; pp. 61–65. [Google Scholar]
- Kephart, K.D.; Buxton, D.R.; Taylor, S.E. Growth of C3 and C4 perennial grasses under reduced irradiance. Crop Sci. 1992, 32, 1033–1038. [Google Scholar] [CrossRef]
- Buxton, D.R.; Lentz, E.M. Performance of morphologically diverse orchardgrass clones in spaced and sward plantings. Grass Forage Sci. 1993, 48, 336–346. [Google Scholar] [CrossRef]
- Bahmani, I.; Hazard, L.; Varlet-Granchet, C.; Betin, M.; Lemaire, G.; Matthew, C.; Thom, E.R. Differences in tillering of long- and short-leaved perennial ryegrass genetic lines under full light and shade treatments. Crop Sci. 2000, 40, 1095–1102. [Google Scholar] [CrossRef]
- Gautier, H.; Varlet-Grancher, C. Regulation of leaf growth of grass by blue light. Physiol. Plant. 1996, 98, 424–430. [Google Scholar] [CrossRef]
- Hazard, L.; Ghesquière, M. Evidence from the use of isozyme markers of competition in swards between short-leaved and long-leaved perennial ryegrass. Grass Forage Sci. 1995, 50, 241–248. [Google Scholar] [CrossRef]
- Rhodes, I.; Stern, W.R. Competition for light. In Plant Relations in Pastures; CSIRO: East Melborne, Australia, 1978; pp. 175–189. [Google Scholar]
- Prieto, I.; Violle, C.; Barre, P.; Durand, J.L.; Ghesquière, M.; Litrico, I. Complementary effects of species and genetic diversity on productivity and stability of sown grasslands. Nat. Plant. 2015, 1, 1–5. [Google Scholar] [CrossRef]
- Kemp, D.R.; Eagles, C.F.; Humphreys, M.O. Leaf growth and apex development of perennial ryegrass during winter and spring. Ann. Bot. 1989, 63, 349–355. [Google Scholar]
- Gastal, F.; Belanger, G.; Lemaire, G. A model of the leaf extension rate of tall fescue in response to nitrogen and temperature. Ann. Bot. 1992, 70, 437–442. [Google Scholar]
- Parsons, A.J.; Robson, M.J. Seasonal changes in the physiology of S24 perennial ryegrass (Lolium perenne L.). I. Response of leaf extension to temperature during the transition from vegetative to reproductive growth. Ann. Bot. 1980, 46, 435–444. [Google Scholar]
- Ball, C.C.; Parsons, A.J.; Rasmussen, S.; Shaw, C.; Rowarth, J.S. Seasonal differences in the capacity of perennial ryegrass to respond to gibberellin explained. Proc. N. Z. Grassl. Assoc. 2012, 74, 183–188. [Google Scholar]
- Parsons, A.J.; Rasmussen, S.; Liu, Q.; Xue, H.; Ball, C.; Shaw, C. Plant growth—Resource or strategy limited: Insights from responses to gibberellin. Grass Forage Sci. 2013, 68, 577–588. [Google Scholar] [CrossRef]
- Bernier, G. The control of floral evocation and morphogenesis. Ann. Rev. Plant Phys. Plant Mol. Biol. 1988, 39, 175–219. [Google Scholar] [CrossRef]
- Wilkins, P.W. Genotype harvesting frequency and genotype nitrogen level interactions for annual dry-matter yield in Lolium perenne in relation to breeding. Euphytica 1989, 41, 207–214. [Google Scholar]
- Peacock, M. Temperature and leaf growth in Lolium perenne. II the site of temperature perception. J. Appl. Ecol. 1975, 12, 115–123. [Google Scholar] [CrossRef]
- Peacock, J.M. Temperature and leaf growth in Lolium perenne. III. Factors affecting seasonal differences. J. Appl. Ecol. 1975, 12, 685–697. [Google Scholar] [CrossRef]
- Cooper, J.P.; Edwards, D. The genetic control of leaf development in Lolium. I. Assessment of genetic variation. Heredity 1961, 16, 63–82. [Google Scholar] [CrossRef]
- Auzanneau, J.; Huyghe, C.; Escobar-Gutierrez, A.J.; Julier, B.; Gastal, F.; Barre, P. Association study between the gibberellic acid insensitive gene and leaf length in a Lolium perenne L. synthetic variety. BMC Plant Biol. 2011, 11, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Barre, P.; Moreau, L.; Mi, F.; Turner, L.; Gastal, F.; Julier, B.; Ghesquiere, M. Quantitative trait loci for leaf length in perennial ryegrass (Lolium perenne L.). Grass Forage Sci. 2009, 64, 310–321. [Google Scholar] [CrossRef]
- Sartie, A.M.; Matthew, C.; Easton, H.S.; Faville, M.J. Phenotypic and QTL analyses of herbage production-related traits in perennial ryegrass (Lolium perenne L.). Euphytica 2011, 182, 295–315. [Google Scholar] [CrossRef]
- Kobayashi, S.; Humphreys, M.O.; Tase, K.; Sanada, Y.; Yamada, T. Molecular marker dissection of ryegrass plant development and its response to growth environments and foliage cuts. Crop Sci. 2011, 51, 600–611. [Google Scholar] [CrossRef]
- Pearson, A.; Cogan, N.O.I.; Baillie, R.C.; Hand, M.L.; Bandaranayake, C.K.; Erb, S.; Wang, J.; Kearney, G.A.; Gendall, A.R.; Smith, K.F.; et al. Identification of QTLs for morphological traits influencing waterlogging tolerance in perennial ryegrass (Lolium perenne L.). Theor. Appl. Genet. 2011, 122, 609–622. [Google Scholar] [CrossRef] [PubMed]
- Turner, L.; Cairns, A.; Armstead, I.; Thomas, H.; Humphreys, M.W.; Humphreys, M.O. Does fructan have a functional role in physiological traits? Investigation by quantitative trait locus mapping. New Phytol. 2008, 179, 765–777. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.; Jensen, L.; Asp, T.; Lübberstedt, T. Vernalization response in perennial ryegrass (Lolium perenne L.) involves orthologues of diploid wheat (Triticum monococcum) Vrn1 and rice (Oryza sativa) HD1. Plant Mol. Biol. 2006, 60, 481–494. [Google Scholar] [CrossRef] [PubMed]
- Jokela, V.; Virkajarvi, P.; Tanskanen, J.; Seppanen, M.M. Vernalization, gibberellic acid and photoperiod are important signals of yield formation in timothy (Phleum pratense). Physiol. Plant. 2014, 152, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Fjellheim, S.; Boden, S.; Trevaskis, B. The role of seasonal flowering responses in adaptation of grasses to temperate climates. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghesquière, M.; Hazard, L.; Betin, M. Breeding for management adaptation in perennial ryegrass (Lolium perenne L.). II. Genetic variability and heritability of leaf morphogenesis components. Agronomie 1994, 14, 267–272. [Google Scholar] [CrossRef]
- Edwards, D.; Cooper, J.P. The genetic control of leaf development in Lolium. II. Response to selection. Heredity 1963, 18, 307–317. [Google Scholar] [CrossRef]
- Reeder, L.; Sleper, D.; Nelson, C. Response to selection for leaf area expansion rate of tall fescue. Crop Sci. 1984, 24, 97–100. [Google Scholar] [CrossRef]
- Shinozuka, H.; Cogan, N.O.I.; Spangenberg, G.C.; Forster, J.W. Quantitative trait locus (QTL) meta-analysis and comparative genomics for candidate gene prediction in perennial ryegrass (Lolium perenne L.). BMC Genet. 2012, 13, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Barre, P.; Cameleyre, I.; Huyghe, C.; Gastal, F.; Lübberstedt, T.; INRA, Lusignan, France. Unpublished work. 2007.
- Xu, S.Z. Theoretical basis of the beavis effect. Genetics 2003, 165, 2259–2268. [Google Scholar] [PubMed]
- Bernardo, R. Molecular markers and selection for complex traits in plants: Learning from the last 20 years. Crop Sci. 2008, 48, 1649–1664. [Google Scholar] [CrossRef]
- Yamada, T.; Jones, E.S.; Cogan, N.O.I.; Vecchies, A.C.; Nomura, T.; Hisano, H.; Shimamoto, Y.; Smith, K.F.; Hayward, M.D.; Forster, J.W. QTL analysis of morphological, developmental, and winter hardiness-associated traits in perennial ryegrass. Crop Sci. 2004, 44, 925–935. [Google Scholar] [CrossRef]
- Pauly, L.; Flajoulot, S.; Garon, J.; Julier, B.; Beguier, V.; Barre, P. Detection of favorable alleles for plant height and crown rust tolerance in three connected populations of perennial ryegrass (Lolium perenne L.). Theor. Appl. Genet. 2012, 124, 1139–1153. [Google Scholar] [CrossRef] [PubMed]
- Armstead, I.; Turner, L.; Marshall, A.; Humphreys, M.; King, I.; Thorogood, D. Identifying genetic components controlling fertility in the outcrossing grass species perennial ryegrass ( Lolium perenne L.) by quantitative trait loci analysis and comparative genetics. New Phytol. 2008, 178, 559–571. [Google Scholar] [CrossRef] [PubMed]
- Studer, B.; Jensen, L.B.; Hentrup, S.; Brazauskas, G.; Kölliker, R.; Lübberstedt, T. Genetic characterisation of seed yield and fertility traits in perennial ryegrass (Lolium perenne L.). Theor. Appl. Genet. 2008, 117, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Barillot, R.; Frak, E.; Combes, D.; Durand, J.-L.; Escobar-Gutierrez, A.J. What determines the complex kinetics of stomatal conductance under blueless par in Festuca arundinacea? Subsequent effects on leaf transpiration. J. Exp. Bot. 2010, 61, 2795–2806. [Google Scholar] [CrossRef] [PubMed]
- Forde, B.J. Effect of various environments on the anatomy and growth of perennial ryegrass and cocksfoot. N. Z. J. Bot. 1966, 4, 13. [Google Scholar]
- Duru, M.; Ducrocq, H. Growth and senescence of the successive grass leaves on a tiller. Ontogenic development and effect of temperature. Ann. Bot. 2000, 85, 635–643. [Google Scholar] [CrossRef]
- Verdenal, A.; Combes, D.; Escobar-Gutierrez, A.J. A study of ryegrass architecture as a self-regulated system, using functional-structural plant modelling. Funct. Plant Biol. 2008, 35, 911–924. [Google Scholar] [CrossRef]
- Gastal, F.; Barre, P.; Carré, S. What are the cellular components underlying the genetic diversity of leaf growth in Lolium perenne L.? In Sustainable Use of Genetic Diversity in Forage and Turf Breeding, Proceedings of the EUCARPIA 2009, La Rochelle, France, 11–14 May 2009; Huyghe, C., Ed.; Springer: Berlin, Germany, 2010; pp. 95–100. [Google Scholar]
- Sugiyama, S. Developmental basis of interspecific differences in leaf size and specific leaf area among C3 grass species. Funct. Ecol. 2005, 19, 916–924. [Google Scholar] [CrossRef]
- Casey, I.; Brereton, A.; Laidlaw, A.; McGilloway, D. Effects of sheath tube length on leaf development in perennial ryegrass (Lolium perenne L.). Ann. Appl. Biol. 1999, 134, 251–257. [Google Scholar] [CrossRef]
- Wilson, R.E.; Laidlaw, A.S. The role of the sheath tube in the development of expanding leaves in perennial ryegrass. Ann. Appl. Biol. 1985, 106, 385–391. [Google Scholar] [CrossRef]
- Andrieu, B.; Hillier, J.; Birch, C. Onset of sheath extension and duration of lamina extension are major determinants of the response of maize lamina length to plant density. Ann. Bot. 2006, 98, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Verdenal, A.; Combes, D.; Escobar-Gutiérrez, A.J. Programmable and self-organised processes in plant morphogenesis: The architectural development of ryegrass. In Studies on Complexity: Morphogenetic Engineering; Doursa, R.S., Michel, O., Eds.; Springer: Berlin, Germany, 2012; pp. 501–517. [Google Scholar]
- Verdenal, A. De la Simulation de la Morphogénèse de L’appareil Aérien du Ray-Grass Anglais (Lolium Perenne L.). Exploration d’un Schéma Cybernétique Inspiré du Concept D’auto-Organisation et Applications. Ph.D. Thesis, The Université de Poitiers, Poitiers, France, December 2009; p. 190. [Google Scholar]
- Migault, V.; Combes, D.; Barre, P.; Gueye, B.; Louarn, G.; Escobar-Gutiérrez, A.J. Improved modelling of ryegrass foliar growth. In Proceedings of the Plant Growth Modeling, Simulation, Visualization and Applications—PMA12, Shanghai, China, 31 October–3 November 2012; IEEE: Beijing, China, 2012; pp. 282–288. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barre, P.; Turner, L.B.; Escobar-Gutiérrez, A.J. Leaf Length Variation in Perennial Forage Grasses. Agriculture 2015, 5, 682-696. https://doi.org/10.3390/agriculture5030682
Barre P, Turner LB, Escobar-Gutiérrez AJ. Leaf Length Variation in Perennial Forage Grasses. Agriculture. 2015; 5(3):682-696. https://doi.org/10.3390/agriculture5030682
Chicago/Turabian StyleBarre, Philippe, Lesley B. Turner, and Abraham J. Escobar-Gutiérrez. 2015. "Leaf Length Variation in Perennial Forage Grasses" Agriculture 5, no. 3: 682-696. https://doi.org/10.3390/agriculture5030682
APA StyleBarre, P., Turner, L. B., & Escobar-Gutiérrez, A. J. (2015). Leaf Length Variation in Perennial Forage Grasses. Agriculture, 5(3), 682-696. https://doi.org/10.3390/agriculture5030682