
Effects of Biochar Blends on Microbial Community Composition in Two Coastal Plain Soils



Abstract
:1. Introduction
2. Results
2.1. Soil Characteristics in Response to Biochar Amendment

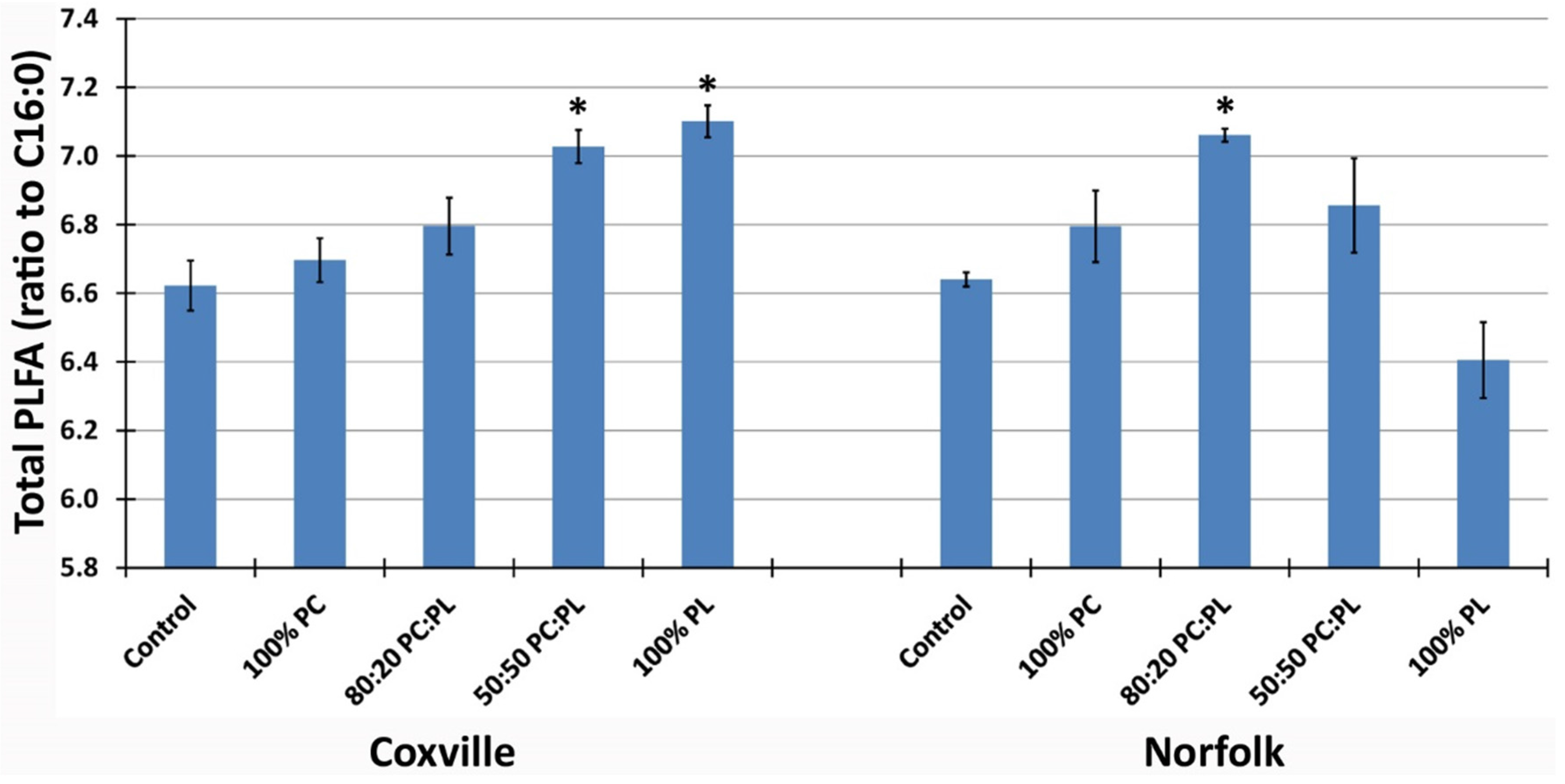{kind=link}
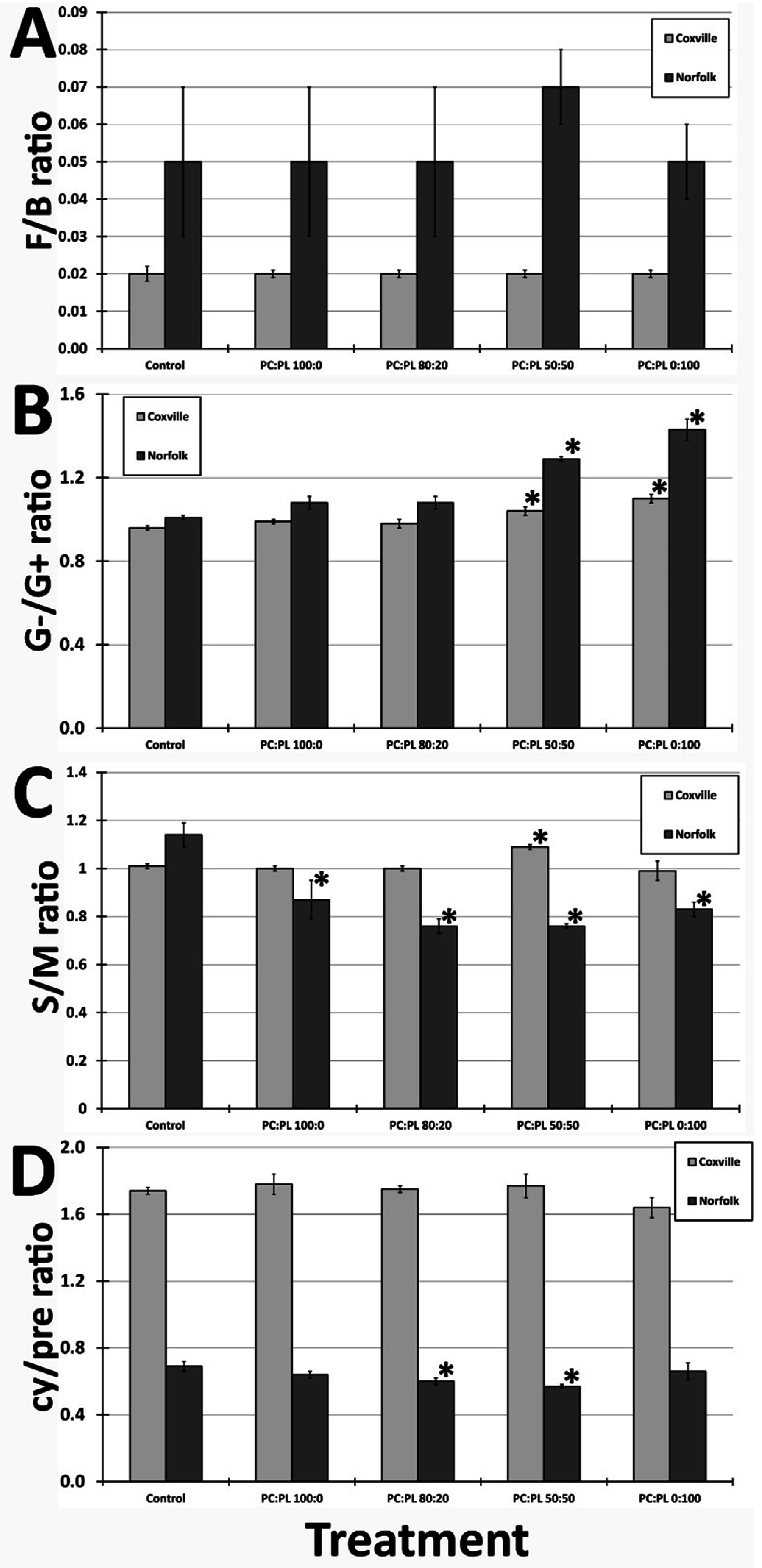{kind=link}
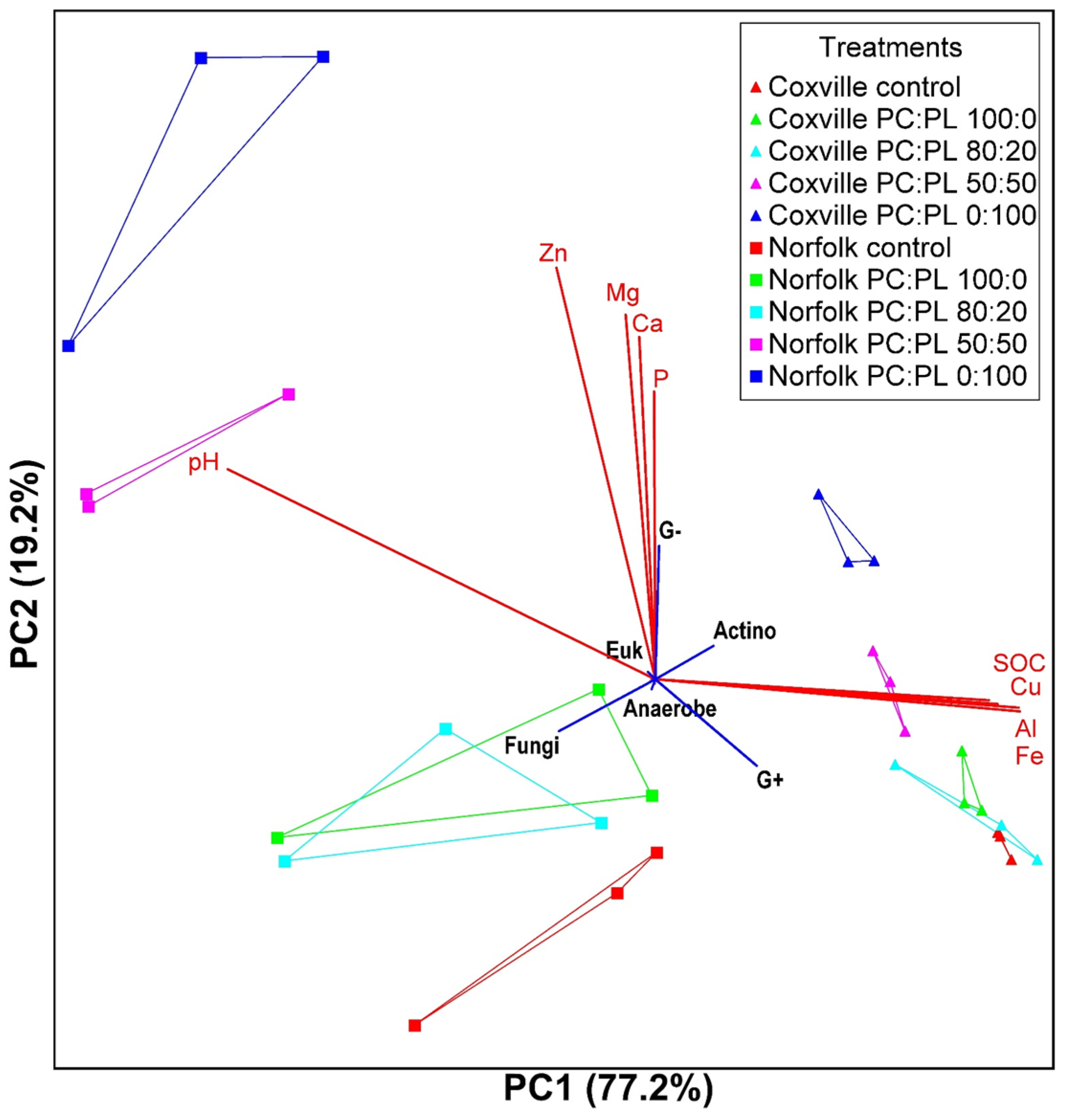{kind=link}
| pH | SOC (g·kg−1) | ||
|---|---|---|---|
| Coxville | Control | 5.1 ± 0.1 c,†,‡ | 47.8 ± 0.1 c |
| PC:PL 100:0 | 5.0 ± 0.1 c | 57.8 ± 0.9 a | |
| PC:PL 80:20 | 5.2 ± 0.1 b c | 55.9 ± 0.8 a b | |
| PC:PL 50:50 | 5.5 ± 0.1 a b | 54.1 ± 0.7 b | |
| PC:PL 0:100 | 5.7 ± 0.1 a | 54.6 ± 0.9 b | |
| Norfolk | Control | 6.0 ± 0.1 c | 12.6 ± 0.5 c |
| PC:PL 100:0 | 6.3 ± 0.1 c | 22.5 ± 0.5 a | |
| PC:PL 80:20 | 7.0 ± 0.1 b | 22.8 ± 0.3 a | |
| PC:PL 50:50 | 7.7 ± 0.1 a | 20.1 ± 0.3 a b | |
| PC:PL 0:100 | 7.9 ± 0.2 a | 17.7 ± 2.5 b |
2.2. Microbial Community Composition in Response to Biochar Amendment
| Mehlich-1 Extractable Nutrients (mg·kg−1) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Al | Ca | Cu | Fe | K | Mg | Mn | Na | P | Zn | ||
| Coxville | Control | 650.9 ± 60.2 a b,†,‡ | 235.8 ± 16.0 c | 0.7 ± 0.1 a | 48.3 ± 4.2 a b | 42.8 ± 3.8 d | 37.4 ± 1.3 c | 15.4 ± 1.6 a | 7.8 ± 1.2 c | 66.2 ± 7.7 d | 3.1 ± 0.1 c |
| PC:PL 100:0 | 675.8 ± 102.4 a b | 234.7 ± 34.5 c | 0.6 ± 0.1 a | 53.2 ± 7.7 a b | 45.7 ± 7.6 d | 36.2 ± 5.5 c | 18.4 ± 3.3 a | 7.6 ± 0.2 c | 68.7 ± 11.8 d | 3.1 ± 0.7 c | |
| PC:PL 80:20 | 759.8 ± 27.8 a | 304.1 ± 7.1 b c | 1.0 ± 0.2 a | 60.5 ± 3.9 a | 130.2 ± 3.4 c | 52.9 ± 1.1 c | 19.9 ± 1.3 a | 24.3 ± 2.0 b c | 110.7 ± 7.2 c | 3.9 ± 0.4 b c | |
| PC:PL 50:50 | 637.4 ± 70.9 a b | 381.6 ± 39.8 a b | 0.8 ± 0.1 a | 48.5 ± 6.1 a b | 271.8 ± 33.2 b | 84.0 ± 11.8 b | 17.0 ± 3.1 a | 60.2 ± 18.5 a b | 165.9 ± 25.8 b | 5.4 ± 0.9 a b | |
| PC:PL 0:100 | 587.9 ± 18.9 b | 416.6 ± 37.2 a | 0.9 ± 0.1 a | 44.3 ± 2.2 b | 388.3 ± 30.7 a | 108.4 ± 11.8 a | 17.1 ± 1.7 a | 95.9 ± 23.5 a | 228.6 ± 13.4 a | 6.9 ± 1.0 a | |
| Norfolk | Control | 89.3 ± 6.5 b | 176.5 ± 6.0 c | ND § | 8.4 ± 0.8 b c | 30.6 ± 10.2 c | 22.7 ± 0.7 d | 3.7 ± 0.2 d | 4.5 ± 0.4 c | 11.1 ± 1.5 d | 3.3 ± 0.1 c |
| PC:PL 100:0 | 86.2 ± 14.0 b | 193.7 ± 4.6 c | ND | 9.1 ± 0.5 a b | 25.3 ± 2.5 c | 23.5 ± 0.8 d | 4.0 ± 0.8 d | 3.3 ± 0.3 c | 11.4 ± 1.6 d | 2.9 ± 0.2 d | |
| PC:PL 80:20 | 87.3 ± 9.5 b | 204.9 ± 13.4 c | ND | 6.7 ± 0.3 d | 64.6 ± 3.7 c | 32.9 ± 2.5 c | 6.4 ± 1.0 c | 11.3 ± 1.0 b c | 28.9 ± 1.7 c | 3.1 ± 0.1 c d | |
| PC:PL 50:50 | 101.0 ± 5.0 a b | 330.5 ± 6.2 b | ND | 7.4 ± 0.3 c d | 148.4 ± 2.8 b | 77.2 ± 2.9 b | 9.3 ± 0.7 b | 31.1 ± 1.9 b | 112.7 ± 2.6 b | 6.5 ± 0.1 b | |
| PC:PL 0:100 | 121.2 ± 3.8 a | 509.9 ± 17.5 a | ND | 9.6 ± 0.1 a | 244.9 ± 39.4 a | 130.0 ± 4.0 a | 13.5 ± 0.1a | 61.5 ± 19.3 a | 236.5 ± 7.0 a | 9.8 ± 0.1 a | |


| Soil | PLFA Group | Control | PC:PL 100:0 | PC:PL 80:20 | PC:PL 50:50 | PC:PL 0:100 |
|---|---|---|---|---|---|---|
| Coxville | Gram-positive | 37.39 ± 0.18 † | 36.82 ± 0.20 | 37.03 ± 0.88 | 35.86 ± 0.32 * | 34.99 ± 0.20 * |
| Gram-negative | 36.04 ± 0.03 | 36.35 ± 0.32 | 36.17 ± 0.22 | 37.15 ± 0.34 * | 38.44 ± 0.48 * | |
| Actinomycetes | 19.90 ± 0.27 | 20.10 ± 0.14 | 19.94 ± 0.35 | 19.61 ± 0.18 | 19.49 ± 0.31 | |
| Fungi | 1.52 ± 0.18 | 1.59 ± 0.08 | 1.62 ± 0.43 | 1.81 ± 0.04 | 1.74 ± 0.16 | |
| Eukarya | 0.58 ± 0.30 | 0.42 ± 0.08 | 0.55 ± 0.32 | 1.00 ± 0.02 | 0.96 ± 0.11 | |
| Norfolk | Gram-positive | 34.80 ± 0.44 | 33.91 ± 0.93 | 33.55 ± 1.17 | 29.93 ± 0.68 * | 28.63 ± 0.38 * |
| Gram-negative | 35.24 ± 0.74 | 36.58 ± 0.48 | 36.24 ± 0.89 | 38.68 ± 0.81 * | 41.00 ± 1.09 * | |
| Actinomycetes | 18.25 ± 0.37 | 17.69 ± 0.93 | 17.64 ± 0.44 | 17.22 ± 0.23 * | 18.36 ± 1.32 | |
| Fungi | 4.61 ± 1.55 | 4.19 ± 2.00 | 4.59 ± 1.56 | 5.61 ± 1.18 | 4.61 ± 1.18 | |
| Eukarya | 0.74 ± 0.02 | 0.97 ± 0.21 | 0.82 ± 0.10 | 1.35 ± 0.86 | 1.12 ± 0.04 * |

| Axis 1 | Axis 2 | |||||
|---|---|---|---|---|---|---|
| r | r2 | tau | r | r2 | tau | |
| pH | −0.957 | 0.917 | −0.795 | 0.018 | 0.000 | −0.181 |
| Al | 0.795 | 0.631 | 0.526 | 0.377 | 0.142 | 0.127 |
| Ca | −0.244 | 0.059 | −0.012 | 0.734 | 0.538 | −0.031 |
| Cu | 0.768 | 0.582 | 0.570 | 0.386 | 0.149 | 0.142 |
| Fe | 0.797 | 0.635 | 0.568 | 0.352 | 0.124 | 0.127 |
| K | −0.126 | 0.016 | −0.054 | 0.714 | 0.510 | 0.034 |
| Mg | −0.358 | 0.128 | −0.045 | 0.573 | 0.588 | 0.000 |
| Mn | 0.495 | 0.245 | 0.376 | 0.553 | 0.305 | 0.066 |
| Na | −0.152 | 0.023 | −0.020 | 0.721 | 0.520 | 0.050 |
| P | −0.187 | 0.035 | 0.041 | 0.819 | 0.670 | 0.066 |
| Zn | −0.506 | 0.256 | −0.149 | 0.658 | 0.432 | −0.062 |
| SOC | 0.727 | 0.529 | 0.529 | 0.415 | 0.172 | 0.065 |
3. Discussion
4. Experimental Section
4.1. Feedstocks and Pyrolysis
4.2. Soils and Biochar Incubation
4.3. Soil and Biochar Chemical Characterization and Phospholipid Fatty Acid (PLFA) Analysis
| Soil Series | pH | SOC † (g·kg−1) | TN † (g·kg−1) | |
| Coxville | 5.1 | 26.3 | 1.8 | |
| Norfolk | 5.9 | 3.9 | ND ‡ | |
| Feedstock | pH | C (g·kg−1) | N (g·kg−1) | C:N ratio |
| Pine chip | 5.3 | 787 | 3.7 | 213:1 |
| Poultry litter | 9.4 | 511 | 56.1 | 9:1 |
4.4. Data Analysis
5. Summary and Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Biederman, L.A.; Harpole, W.S. Biochar and its effects on plant productivity and nutrient cycling: A meta-analysis. GCB Bioenergy 2013, 5, 202–214. [Google Scholar] [CrossRef]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D. Biochar effects on soil biota—A review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Lehmann, J.; Joseph, S. Biochar for Environmental Management: Science and Technology; Earthscan: London, UK, 2009; p. 416. [Google Scholar]
- Busscher, W.J.; Novak, J.M.; Ahmedna, M. Physical effects of organic matter amendment of a southeastern US coastal loamy sand. Soil Sci. 2011, 176, 661–667. [Google Scholar] [CrossRef]
- Kelly, C.N.; Calderon, F.C.; Acosta-Martinez, V.; Mikha, M.M.; Benjamin, J.; Rutherford, D.W.; Rostad, C.E. Switchgrass biochar effects on plant biomass and microbial dynamics in two soils from different regions. Pedosphere 2015, 25, 329–342. [Google Scholar] [CrossRef]
- Deenik, J.L.; McClellan, T.; Uehara, G.; Antal, M.J.; Campbell, S. Charcoal volatile matter content influences plant growth and soil nitrogen transformations. Soil Sci. Soc. Am. J. 2010, 74, 1259–1270. [Google Scholar] [CrossRef]
- Novak, J.M.; Lima, I.; Xing, B.; Gaskin, J.W.; Steiner, C.; Das, K.C.; Ahmedna, M.; Rehrah, D.; Watts, D.W.; Busscher, W.J.; et al. Characterization of designer biochar produced at different temperatures and their effects on a loamy sand. Ann. Environ. Sci. 2009, 3, 195–206. [Google Scholar]
- Spokas, K.A.; Cantrell, K.B.; Novak, J.M.; Archer, D.W.; Ippolito, J.A.; Collins, H.P.; Boateng, A.A.; Lima, I.M.; Lamb, M.C.; McAloon, A.J.; et al. Biochar: A synthesis of its agronomic impact beyond carbon sequestration. J. Environ. Qual. 2012, 41, 973–989. [Google Scholar] [CrossRef] [PubMed]
- Sigua, G.C.; Novak, J.M.; Watts, D.W.; Cantrell, K.B.; Shumaker, P.D.; Szogi, A.A.; Johnson, M.G. Carbon mineralization in two ultisols amended with different sources and particle sizes of pyrolyzed biochar. Chemosphere 2014, 103, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Herath, H.M.S.K.; Camps-Arbestain, M.; Hedley, M. Effect of biochar on soil physical properties in two contrasting soils: An alfisol and an andisol. Geoderma 2013, 209, 188–197. [Google Scholar] [CrossRef]
- Kolb, S.E.; Fermanich, K.J.; Dornbush, M.E. Effect of charcoal quantity on microbial biomass and activity in temperate soils. Soil Sci. Soc. Am. J. 2009, 73, 1173–1181. [Google Scholar] [CrossRef]
- Ippolito, J.A.; Stromberger, M.E.; Lentz, R.D.; Dungan, R.S. Hardwood biochar and manure co-application to a calcareous soil. Chemosphere 2015. [Google Scholar] [CrossRef] [PubMed]
- Ducey, T.F.; Ippolito, J.A.; Cantrell, K.B.; Novak, J.M.; Lentz, R.D. Addition of activated switchgrass biochar to an aridic subsoil increases microbial nitrogen cycling gene abundances. Appl. Soil Ecol. 2013, 65, 65–72. [Google Scholar] [CrossRef]
- Ippolito, J.A.; Ducey, T.F.; Cantrell, K.B.; Novak, J.M.; Lentz, R.D. Designer, acidic biochar influences calcareous soil characteristics. Chemosphere 2015. [Google Scholar] [CrossRef] [PubMed]
- Fox, T.R.; Jokela, E.J.; Allen, H.L. The development of pine plantation silviculture in the southern United States. J. Forest. 2007, 105, 337–347. [Google Scholar]
- USDA-NASS. Poultry—Production and Value 2013 Summary; USDA-NASS: Washington, DC, USA, 2014; pp. 1–14.
- Novak, J.M.; Cantrell, K.B.; Watts, D.W.; Busscher, W.J.; Johnson, M.G. Designing relevant biochars as soil amendments using lignocellulosic-based and manure-based feedstocks. J. Soil Sediment. 2014, 14, 330–343. [Google Scholar] [CrossRef]
- Buyer, J.S.; Sasser, M. High throughput phospholipid fatty acid analysis of soils. Appl. Soil Ecol. 2012, 61, 127–130. [Google Scholar] [CrossRef]
- Bartelt-Ryser, J.; Joshi, J.; Schmid, B.; Brandl, H.; Balser, T. Soil feedbacks of plant diversity on soil microbial communities and subsequent plant growth. Perspect. Plant Ecol. 2005, 7, 27–49. [Google Scholar] [CrossRef]
- Bohn, H.L.; McNeal, B.L.; O’Connor, G.A. Soil Chemistry; Wiley and Sons Inc.: New York, NY, USA, 1979; p. 329. [Google Scholar]
- Sims, J.T.; Simard, R.R.; Joern, B.C. Phosphorus loss in agricultural drainage: Historical perspective and current research. J. Environ. Qual. 1998, 27, 277–293. [Google Scholar] [CrossRef]
- Novak, J.M.; Watts, D.W.; Hunt, P.G.; Stone, K.C. Phosphorus movement through a coastal plain soil after a decade of intensive swine manure application. J. Environ. Qual. 2000, 29, 1310–1315. [Google Scholar] [CrossRef]
- Safaya, N.M. Phosphorus-zinc interaction in relation to absorption rates of phosphorus, zinc, copper, manganese, and iron in corn. Soil Sci. Soc. Am. J. 1976, 40, 719–722. [Google Scholar] [CrossRef]
- Singh, J.P.; Karamanos, R.E.; Stewart, J.W.B. The mechanism of phosphorus-induced zinc-deficiency in bean (Phaseolus vulgaris L.). Can. J. Soil Sci. 1988, 68, 345–358. [Google Scholar] [CrossRef]
- Kieft, T.L.; Wilch, E.; OConnor, K.; Ringelberg, D.B.; White, D.C. Survival and phospholipid fatty acid profiles of surface and subsurface bacteria in natural sediment microcosms. Appl. Environ. Microbiol. 1997, 63, 1531–1542. [Google Scholar] [PubMed]
- Bossio, D.A.; Scow, K.M.; Gunapala, N.; Graham, K.J. Determinants of soil microbial communities: Effects of agricultural management, season, and soil type on phospholipid fatty acid profiles. Microb. Ecol. 1998, 36, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Gundersen, P.; Zhang, T.; Mo, J.M. Effects of phosphorus addition on soil microbial biomass and community composition in three forest types in tropical China. Soil Biol. Biochem. 2012, 44, 31–38. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Townsend, A.R.; Schmidt, S.K. Phosphorus limitation of microbial processes in moist tropical forests: Evidence from short-term laboratory incubations and field studies. Ecosystems 2002, 5, 680–691. [Google Scholar] [CrossRef]
- Treseder, K.K.; Allen, M.F. Direct nitrogen and phosphorus limitation of arbuscular mycorrhizal fungi: A model and field test. New Phytol. 2002, 155, 507–515. [Google Scholar] [CrossRef]
- Elzobair, K.A.; Stromberger, M.E.; Ippolito, J.A.; Lentz, R.D. Contrasting effects of biochar versus manure on soil microbial communities and enzyme activities in an aridisol. Chemosphere 2015. [Google Scholar] [CrossRef] [PubMed]
- Rutigliano, F.A.; Romano, M.; Marzaioli, R.; Baglivo, I.; Baronti, S.; Miglietta, F.; Castaldi, S. Effect of biochar addition on soil microbial community in a wheat crop. Eur. J. Soil Biol. 2014, 60, 9–15. [Google Scholar] [CrossRef]
- Castaldi, S.; Riondino, M.; Baronti, S.; Esposito, F.R.; Marzaioli, R.; Rutigliano, F.A.; Vaccari, F.P.; Miglietta, F. Impact of biochar application to a Mediterranean wheat crop on soil microbial activity and greenhouse gas fluxes. Chemosphere 2011, 85, 1464–1471. [Google Scholar] [CrossRef] [PubMed]
- Cantrell, K.B.; Martin, J.H. Stochastic state-space temperature regulation of biochar production. Part I: Theoretical development. J. Sci. Food Agric. 2012, 92, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Schimel, J.P.; Holden, P.A. Variations in microbial community composition through two soil depth profiles. Soil Biol. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- ASTM. Standard Practice for Proximate Analysis of Coal and Coke; ASTM International: West Conshohocken, PA, USA, 2013. [Google Scholar]
- ASTM. Standard Practice for Ultimate Analysis of Coal and Coke; ASTM International: West Conshohocken, PA, USA, 2015. [Google Scholar]
- Mehlich, A. Determination of P, K, Ca, Mg, Na and NH4; Department of Agriculture: Raleigh, NC, USA, 1953. [Google Scholar]
- ASTM. Standard Test Method for Ash in Biomass; ASTM International: West Conshohocken, PA, USA, 2015. [Google Scholar]
- Novak, J.M.; Watts, D.W. An alum-based water treatment residual can reduce extractable phosphorus concentrations in three phosphorus-enriched coastal plain soils. J. Environ. Qual. 2005, 34, 1820–1827. [Google Scholar] [CrossRef] [PubMed]
- Gomez, J.D.; Denef, K.; Stewart, C.E.; Zheng, J.; Cotrufo, M.F. Biochar addition rate influences soil microbial abundance and activity in temperate soils. Eur. J. Soil Sci. 2014, 65, 28–39. [Google Scholar] [CrossRef]
- Drijber, R.A.; Doran, J.W.; Parkhurst, A.M.; Lyon, D.J. Changes in soil microbial community structure with tillage under long-term wheat-fallow management. Soil Biol. Biochem. 2000, 32, 1419–1430. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ducey, T.F.; Novak, J.M.; Johnson, M.G. Effects of Biochar Blends on Microbial Community Composition in Two Coastal Plain Soils. Agriculture 2015, 5, 1060-1075. https://doi.org/10.3390/agriculture5041060
Ducey TF, Novak JM, Johnson MG. Effects of Biochar Blends on Microbial Community Composition in Two Coastal Plain Soils. Agriculture. 2015; 5(4):1060-1075. https://doi.org/10.3390/agriculture5041060
Chicago/Turabian StyleDucey, Thomas F., Jeffrey M. Novak, and Mark G. Johnson. 2015. "Effects of Biochar Blends on Microbial Community Composition in Two Coastal Plain Soils" Agriculture 5, no. 4: 1060-1075. https://doi.org/10.3390/agriculture5041060
APA StyleDucey, T. F., Novak, J. M., & Johnson, M. G. (2015). Effects of Biochar Blends on Microbial Community Composition in Two Coastal Plain Soils. Agriculture, 5(4), 1060-1075. https://doi.org/10.3390/agriculture5041060