Determining the Stability of Sugarcane Filtercake Biochar in Soils with Contrasting Levels of Organic Matter


Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Selection and Biochar Production
2.2. Laboratory Incubation and CO2 Analysis
2.3. Soil Fractionation and Isotope Analysis
2.4. Soil Analysis
2.5. Statistical Analysis
3. Results
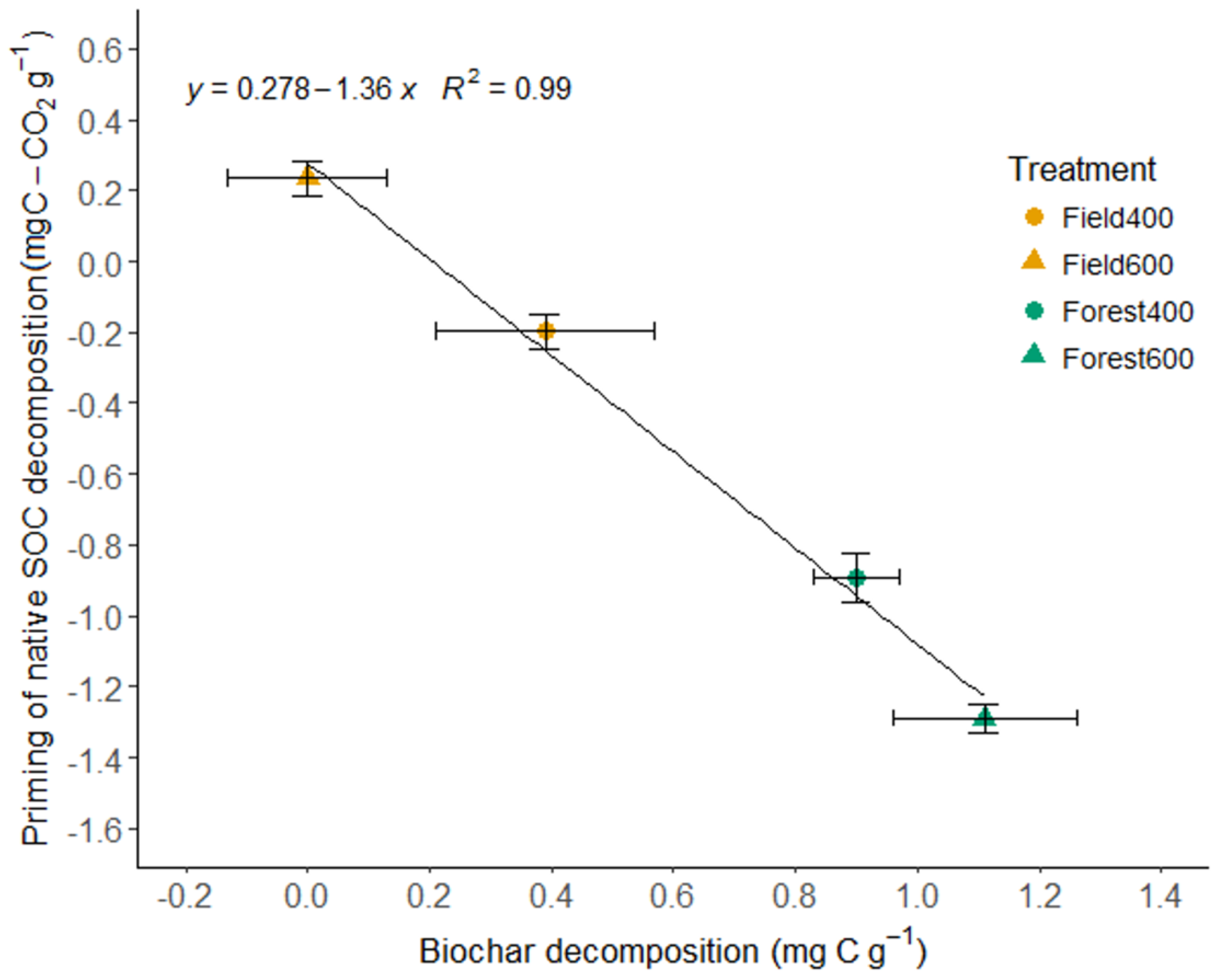
3.1. Biochar C in the Soils
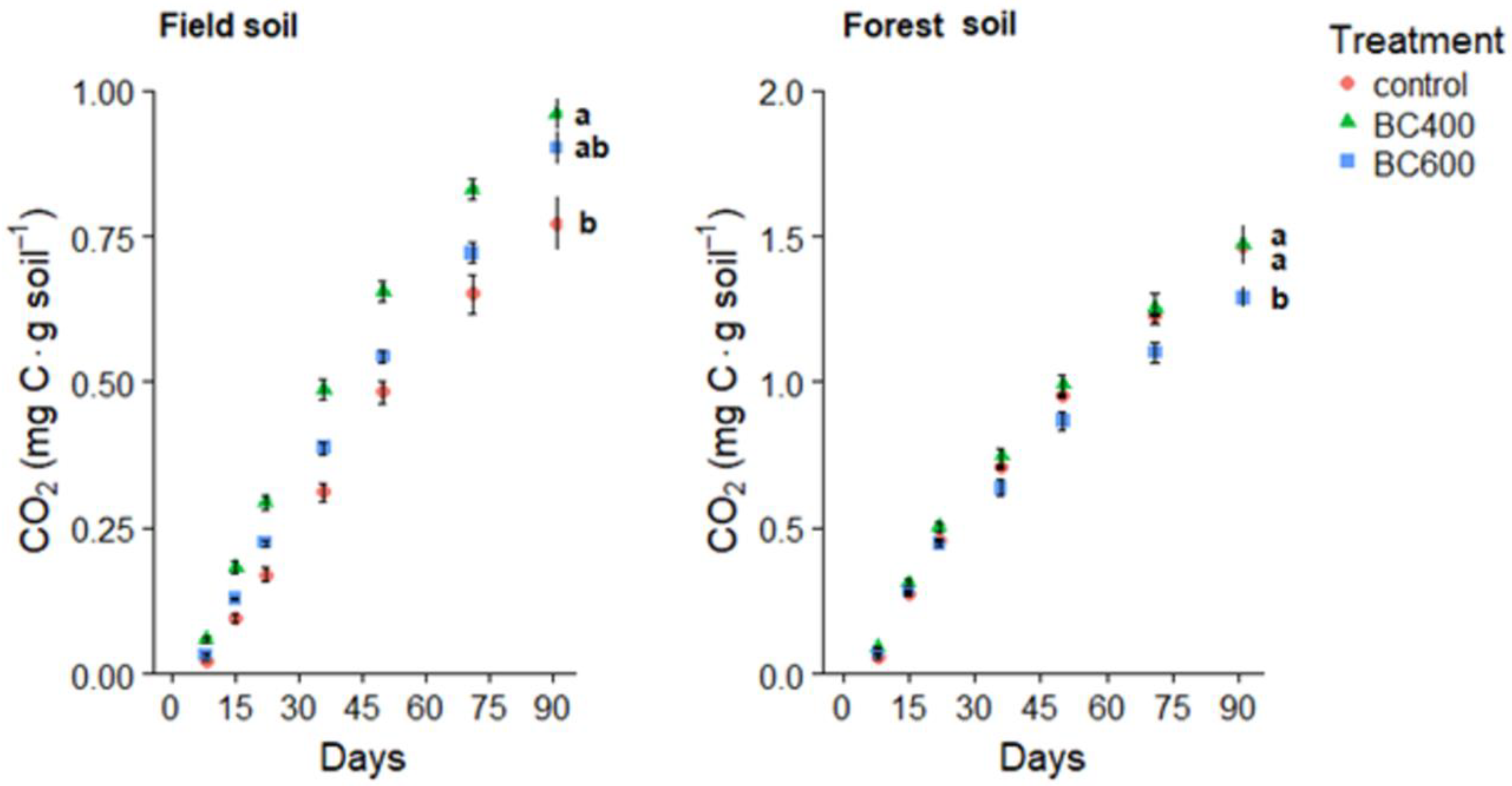
3.2. CO2 Fluxes
3.3. Soil Analysis
4. Discussion
4.1. Biochar C Stabilization
4.2. Biochar Effect on Respiration Rates
4.3. Biochar Influence on Soil Chemical Properties
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Leifeld, J.; Kögel-Knabner, I. Soil organic matter fractions as early indicators for carbon stock changes under different land-use? Geoderma 2005, 124, 143–155. [Google Scholar] [CrossRef]
- Lal, R. Challenges and opportunities in soil organic matter research. Eur. J. Soil Sci. 2009, 60, 158–169. [Google Scholar] [CrossRef]
- Angers, D.A.; Chantigny, M.H.; MacDonald, J.D.; Rochette, P.; Côté, D. Differential retention of carbon, nitrogen and phosphorus in grassland soil profiles with long-term manure application. Nutr. Cycl. Agroecosyst. 2010, 86, 225e229. [Google Scholar] [CrossRef]
- Lehmann, J. A handful of carbon. Nature 2007, 447, 10–11. [Google Scholar] [CrossRef] [PubMed]
- Clough, T.J.; Condron, L.M. Biochar and the nitrogen cycle: Introduction. J. Environ. Qual. 2010, 39, 1218–1223. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.; Joseph, S. Biochar for environmental management: An introduction. In Biochar for Environmental Management: Science and Technology; Lehmann, J., Joseph, S., Eds.; Earthscan: London, UK, 2009; pp. 1–12. [Google Scholar]
- Woolf, D.; Amonette, J.E.; Street-Perrott, F.A.; Lehmann, J.; Joseph, S. Sustainable biochar to mitigate global climate change. Nat. Commun. 2010, 1, 56. [Google Scholar] [CrossRef] [PubMed]
- Woolf, D.; Lehmann, J. Modelling the long-term response to positive and negative priming of soil organic carbon by black carbon. Biogeochemistry 2012, 111, 83–95. [Google Scholar] [CrossRef]
- Singh, B.P.; Cowie, A.L. Long-term influence of biochar on native organic carbon mineralisation in a low-carbon clayey soil. Sci. Rep. 2015, 4, 3687. [Google Scholar] [CrossRef] [PubMed]
- Ameloot, N.; Graber, E.R.; Verheijen, F.G.A.; De Neve, S. Interactions between biochar stability and soil organisms: Review and research needs. Eur. J. Soil Sci. 2013, 64, 379–390. [Google Scholar] [CrossRef]
- Lehmann, J.; Czimczik, C.; Laird, D.; Sohi, S. Stability of biochar in the soil. In Biochar for Environmental Management: Science and Technology; Lehmann, J., Joseph, S., Eds.; Earthscan: London, UK, 2009; pp. 183–205. [Google Scholar]
- Six, J.; Conant, R.T.; Paul, E.A.; Paustian, K. Stabilization mechanisms of soil organic matter: Implications for C-saturation of soils. Plant Soil 2002, 241, 155–176. [Google Scholar] [CrossRef]
- Stewart, C.; Paustian, K.; Conant, R.; Plante, A.; Six, J. Soil carbon saturation: Concept, evidence and evaluation. Biogeochemistry 2007, 86, 19–31. [Google Scholar] [CrossRef]
- Kimetu, J.M.; Lehmann, J. Stability and stabilization of biochar and green manure in soil with different organic carbon contents. Aust. J. Soil Res. 2010, 48, 577–585. [Google Scholar] [CrossRef]
- Eykelbosh, A.J.; Johnson, M.S.; Santos de Queiroz, E.; Dalmagro, H.J.; Guimarães Couto, E. Biochar from sugarcane filtercake reduces soil CO2 emissions relative to raw residue and improves water retention and nutrient availability in a highly-weathered tropical soil. PLoS ONE 2014, 9, e98523. [Google Scholar] [CrossRef] [PubMed]
- Abbruzzini, T.F.; Oliveira Zenero, M.D.; de Andrade, P.A.M.; Dini Andreote, F.; Campo, J.; Pellegrino Cerri, C.E. Effects of biochar on the emissions of greenhouse gases from sugarcane residues applied to soils. Agric. Sci. 2017, 8, 869–886. [Google Scholar] [CrossRef]
- Speratti, A.B.; Johnson, M.S.; Martins Sousa, H.; Nunes Torres, G.; Guimarães Couto, E. Impact of different agricultural waste biochars on maize biomass and soil water content in a Brazilian Cerrado Arenosol. Agronomy 2017, 7, 49. [Google Scholar] [CrossRef]
- Speratti, A.B.; Johnson, M.S.; Martins Sousa, H.; Dalmagro, H.J.; Guimarães Couto, E. Biochars from local agricultural waste residues contribute to soil quality and plant growth in a Cerrado region (Brazil) Arenosol. GCB Bioenergy 2017, 12, 3218–3221. [Google Scholar] [CrossRef]
- Speratti, A.B.; Johnson, M.S.; Martins Sousa, H.; Dalmagro, H.J.; Guimarães Couto, E. Biochar feedstock and pyrolysis temperature effects on leachate: DOC characteristics and nitrate losses from a Brazilian Cerrado Arenosol mixed with agricultural waste biochars. J. Environ. Manag. 2018, 211, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Eykelbosh, A.J.; Johnson, M.S.; Couto, E.G. Biochar decreases dissolved organic carbon but not nitrate leaching in relation to vinasse application in a Brazilian sugarcane soil. J. Environ. Manag. 2015, 149, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Kleber, M.; Hockaday, W.; Nico, P.S.; Chia, C.H.; Downie, A.; Munroe, P. Characteristics of biochar: Macro-molecular properties. In Biochar for Environmental Management: Science, Technology, and Implementation; Lehmann, J., Joseph, S., Eds.; Earthscan: New York, NY, USA, 2015; pp. 11–137. [Google Scholar]
- Anderson, J. Soil respiration. In Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties, 2nd ed.; Page, A.L., Ed.; American Society of Agronomy, Inc.; Soil Science Society of America, Inc.: Madison, WI, USA, 1982; pp. 831–871. [Google Scholar]
- Rovira, P.; Jorba, M.; Romanyà, J. Active and passive organic matter fractions in Mediterranean forest soils. Biol. Fertil. Soils 2010, 46, 355–369. [Google Scholar] [CrossRef]
- Edwards, A.P.; Bremner, J.M. Dispersion of soil particles by sonic vibration. J. Soil Water Conserv. 1967, 18, 47–63. [Google Scholar]
- Cerri, C.; Feller, C.; Balesdent, J.; Victoria, R.; Plenecassagne, A. Application du traçage isotopique naturel en 13C, à l’étude de la dynamique de la matière organique dans le sols. C. R. Acad. Sci. Ser. 2 1985, 9, 423–428. [Google Scholar]
- Nelson, D.; Sommers, L. Total carbon, organic carbon and organic matter. In Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties; Page, A.L., Miller, R., Keeney, D., Eds.; American Society of Agronomy, Inc.; Soil Science Society of America, Inc.: Madison, WI, USA, 1982; pp. 539–579. [Google Scholar]
- Cabrera, M.L.; Beare, M.H. Alkaline persulfate oxidation for determining total nitrogen in microbial biomass extracts. Soil Sci. Soc. Am. J. 1993, 57, 1007–1012. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Joseph, S.D.; Camps-Arbestain, M.; Lin, Y.; Munroe, P.; Chia, C.H.; Hook, J.; van Zwieten, L.; Kimber, S.; Cowie, A.; Singh, B.P.; et al. An investigation into the reactions of biochar in soil. Aust. J. Soil Res. 2010, 48, 501–515. [Google Scholar] [CrossRef]
- Whitman, T.; Singh, B.; Zimmerman, A. Priming effects in biochar-amended soils: Implications of biochar-soil organic matter interactions for carbon storage. In Biochar for Environmental Management: Science, Technology, and Implementation; Lehmann, J., Joseph, S., Eds.; Routledge: New York, NY, USA, 2015; pp. 455–487. [Google Scholar]
- Steinbeiss, S.; Gleixner, G.; Antonietti, M. Effect of biochar amendment on soil carbon balance and soil microbial activity. Soil Biol. Biochem. 2009, 41, 1301–1310. [Google Scholar] [CrossRef]
- Keith, A.; Singh, B.; Singh, B. Interactive priming of biochar and labile organic matter mineralization in a smectite-rich soil. Environ. Sci. Technol. 2011, 45, 9611–9618. [Google Scholar] [CrossRef] [PubMed]
- Whitman, T.; Enders, A.; Lehmann, J. Pyrogenic carbon additions to soil counteract positive priming of soil carbon mineralization by plants. Soil Biol. Biochem. 2014, 73, 33–41. [Google Scholar] [CrossRef]
- Zimmerman, A.R.; Gao, B.; Ahn, M.Y. Positive and negative carbon mineralization priming effects among a variety of biochar-amended soils. Soil Biol. Biochem. 2011, 43, 1169–1179. [Google Scholar] [CrossRef]
- St. Luce, M.; Whalen, J.K.; Ziadi, N.; Zebarth, B.J. Net nitrogen mineralization enhanced with the addition of nitrogen-rich particulate organic matter. Geoderma 2016, 262, 112–118. [Google Scholar] [CrossRef]
- Downie, A.; Crosky, A.; Munroe, P. Physical properties of biochar. In Biochar for Environmental Management: Science and Technology; Lehmann, J., Joseph, S., Eds.; Earthscan: London, UK, 2009; pp. 13–32. [Google Scholar]
- Ameloot, N.; De Neve, S.; Jegajeevagan, K.; Yildiz, G.; Buchan, D.; Funkuin, Y.N.; Prins, W.; Bouckaert, L.; Sleutel, S. Short-term CO2 and N2O emissions and microbial properties of biochar amended sandy loam soils. Soil Biol. Biochem. 2013, 57, 401–410. [Google Scholar] [CrossRef]
- Six, J.; Frey, S.D.; Thiet, R.K.; Batten, K.M. Bacterial and fungal contributions to carbon sequestration in agroecosystems. Soil Sci. Soc. Am. J. 2006, 70, 555–569. [Google Scholar] [CrossRef]
- van Leeuwen, J.P.; Djukic, I.; Bloem, J.; Lehtinen, T.; Hemerik, L.; de Ruiter, P.C.; Lair, G.J. Effects of land use on soil microbial biomass, activity and community structure at different soil depths in the Danube floodplain. Eur. J. Soil Biol. 2017, 79, 14–20. [Google Scholar] [CrossRef]
- Blagodatskaya, E.V.; Anderson, T.-H. Interactive effects of pH and substrate quality on the fungal-to-bacterial ratio and qCO2 of microbial communities in forest soils. Soil Biol. Biochem. 1998, 30, 1269–1274. [Google Scholar] [CrossRef]
- Thies, J.E.; Rillig, M.C.; Graber, E.R. Biochar effects on the abundance, activity and diversity of the soil biota. In Biochar for Environmental Management: Science, Technology, and Implementation; Lehmann, J., Joseph, S., Eds.; Earthscan: New York, NY, USA, 2015; pp. 327–389. [Google Scholar]
- Ameloot, N.; Sleutel, S.; Das, K.C.; Kanagaratnam, J.; de Neve, S. Biochar amendment to soils with contrasting organic matter level: Effects on N mineralization and biological soil properties. Glob. Chang. Biol. Bioenergy 2015, 7, 135–144. [Google Scholar] [CrossRef]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D. Biochar effects on soil biota—A review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Späth, A.; König, B. Molecular recognition of organic ammonium ions in solution using synthetic receptors. Beilstein J. Org. Chem. 2010, 6, 32. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Biochar C (mg C g−1) | Native SOC (mg C-CO2 g−1) | Priming (mg C-CO2 g−1) | Total CO2 (mg C-CO2 g−1) |
|---|---|---|---|---|
| Field400 | 0.39 ± 0.10 | 0.58 ± 0.10 | −0.19 ± 0.12 | 0.96 ± 0.03 |
| Field600 | 0.00 ± 0.10 | 0.89 ± 0.11 | 0.11 ± 0.12 | 0.90 ± 0.03 |
| Field0 | NA | 0.77 ± 0.05 | 0.77 ± 0.05 | |
| Forest400 | 0.90 ± 0.02 | 0.57 ± 0.07 | −0.89 ± 0.07 | 1.47 ± 0.07 |
| Forest600 | 1.11 ± 0.11 | 0.17 ± 0.11 | −1.29 ± 0.12 | 1.29 ± 0.03 |
| Forest0 | NA | 1.47 ± 0.01 | 1.47 ± 0.01 | |
| ANOVA p values | ||||
| Soil | 0.0001 | 0.0001 | 0.0001 | 0.0001 |
| Biochar | n.s. | n.s. | n.s. | 0.015 |
| Soil*Biochar | 0.025 | 0.0001 | 0.0001 | n.s. |
| Condition | Treatment | Total Organic C (%) | Total N (%) | NO3− (mg N g soil−1) | NH4+ (mg N g soil−1) | Mineral N (mg N g soil−1) | DOC (µg C g soil−1) | DON (µg N g soil−1) |
|---|---|---|---|---|---|---|---|---|
| Pre-incubation | Field0 | 1.2 ± 0.03 b | 0.11 ± 0.002 b | 39.8 ± 2.7 | 2.8 ± 0.3 b | 42.7 ± 3.0 | 95.8 ± 6.1 b | 53.1 ± 2.6 |
| Field400 | 1.5 ± 0.03 a | 0.13 ± 0.001 a | 36.3 ± 1.2 | 6.1 ± 1.0 a | 42.5 ± 0.5 | 105.8 ± 5.2 b | 46.7 ± 1.8 | |
| Field600 | 1.5 ± 0.03 a | 0.12 ± 0.002 a | 30.8 ± 3.6 | 5.3 ± 0.4 a,b | 36.1 ± 3.6 | 238.0 ± 14.3 a | 49.0 ± 1.9 | |
| Post-incubation | Field0 | 1.1 ± 0.07 b | 0.11 ± 0.004 | 70.4 ± 5.0 a,b,* | 1.1 ± 0.1 * | 71.3 ± 5.0 a,b,* | 101.4 ± 3.8 | 16.5 ± 2.0 * |
| Field400 | 1.5 ± 0.03 a | 0.12 ± 0.005 | 62.5 ± 7.4 b,* | 0.9 ± 0.6 * | 62.8 ± 7.7 b,* | 84.0 ± 24.7 | 20.6 ± 8.7 * | |
| Field600 | 1.4 ± 0.00 a | 0.12 ± 0.002 | 91.3 ± 6.6 a,* | 0.9 ± 0.3 * | 92.2 ± 7.0 a,* | 123.3 ± 13.3 * | 10.7 ± 4.2 * | |
| Pre-incubation | Forest0 | 2.7 ± 0.07 b | 0.19 ± 0.004 b | 0 | 12.0 ± 0.3 | 3.4 ± 0.3 | 180.4 ± 5.3 b | 34.6 ± 2.2 |
| Forest400 | 3.3 ± 0.13 a | 0.23 ± 0.008 a | 0 | 10.0 ± 0.6 | 6.5 ± 1.4 | 255.3 ± 7.2 a | 43.6 ± 0.3 | |
| Forest600 | 3.2 ± 0.03 a | 0.22 ± 0.005 a | 0 | 11.2 ± 0.6 | 5.4 ± 0.4 | 277.9 ± 11.1 a | 45.1 ± 4.3 | |
| Post-incubation | Forest0 | 2.8 ± 0.09 | 0.20 ± 0.004 | 129.2 ± 9.4 * | 54.2 ± 0.5 a,* | 183.4 ± 9.0 * | 135.9 ± 7.5 * | 0 * |
| Forest400 | 3.0 ± 0.12 | 0.21 ± 0.009 | 164.1 ± 6.3 * | 4.7 ± 1.8 b,* | 168.8 ± 5.2 * | 136.6 ± 3.7 * | 20.4 ± 12.0 | |
| Forest600 | 3.2 ± 0.18 | 0.22 ± 0.013 | 167.2 ± 13.8 * | 0.2 ± 0.1 c,* | 159.7 ± 12.3 * | 133.2 ± 13.0 * | 7.4 ± 7.4 * |
| Treatment | Net Nitrification (mg N g soil−1) | Net Ammonification (mg N g soil−1) | Net N Mineralization (mg N g soil−1) | Change in DOC (µg C g soil−1) | Change in DON (µg N g soil−1) |
|---|---|---|---|---|---|
| Field0 | 30.1 ± 8.7 a,b | −1.8 ± 0.4 a | 28.1 ± 5.7 a,b | 5.7 ± 7.3 a | −37.0 ± 5.3 a |
| Field400 | 25.1 ± 7.2 b | −5.2 ± 0.7 b | 20.3 ± 7.7 b | −30.4 ± 31.0 a | −26.1 ± 9.5 a |
| Field600 | 60.5 ± 7.0 a | −4.4 ± 0.6 b | 56.0 ± 7.4 a | −114.8 ± 4.3 b | −38.4 ± 4.4 a |
| Forest0 | 129.2 ± 9.4 a | 42.2 ± 0.4 a | 180.0 ± 9.2 a | −46.1 ± 9.0 a | −34.6 ± 2.2 a |
| Forest400 | 164.1 ± 6.3 a | −5.4 ± 2.1 b | 161.4 ± 4.4 a | −118.8 ± 6.0 b | −23.2 ± 12.1 a |
| Forest600 | 167.2 ± 13.8 a | −11.1 ± 0.9 c | 154.4 ± 12.3 a | −144.7 ± 15.6 b | −40.8 ± 10.3 a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Speratti, A.B.; Romanyà, J.; Garcia-Pausas, J.; Johnson, M.S. Determining the Stability of Sugarcane Filtercake Biochar in Soils with Contrasting Levels of Organic Matter. Agriculture 2018, 8, 71. https://doi.org/10.3390/agriculture8060071
Speratti AB, Romanyà J, Garcia-Pausas J, Johnson MS. Determining the Stability of Sugarcane Filtercake Biochar in Soils with Contrasting Levels of Organic Matter. Agriculture. 2018; 8(6):71. https://doi.org/10.3390/agriculture8060071
Chicago/Turabian StyleSperatti, Alicia B., Joan Romanyà, Jordi Garcia-Pausas, and Mark S. Johnson. 2018. "Determining the Stability of Sugarcane Filtercake Biochar in Soils with Contrasting Levels of Organic Matter" Agriculture 8, no. 6: 71. https://doi.org/10.3390/agriculture8060071
APA StyleSperatti, A. B., Romanyà, J., Garcia-Pausas, J., & Johnson, M. S. (2018). Determining the Stability of Sugarcane Filtercake Biochar in Soils with Contrasting Levels of Organic Matter. Agriculture, 8(6), 71. https://doi.org/10.3390/agriculture8060071