Utilization of Molecular Marker Based Genetic Diversity Patterns in Hybrid Parents to Develop Better Forage Quality Multi-Cut Hybrids in Pearl Millet

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. DNA Isolation
2.3. Genotyping of Hybrid Parents
2.3.1. Simple Sequence Repeats
2.3.2. Polymerase Chain Reaction
2.3.3. Genotyping by Sequencing (GBS) and SNP Calling
2.4. Phenotyping of Hybrid Parents
2.5. Hybrid Development and Phenotypic Evaluation
2.6. Data Analysis
3. Results
3.1. Genetic Diversity Indicators Based on SSRs and GBS-Identified SNPs
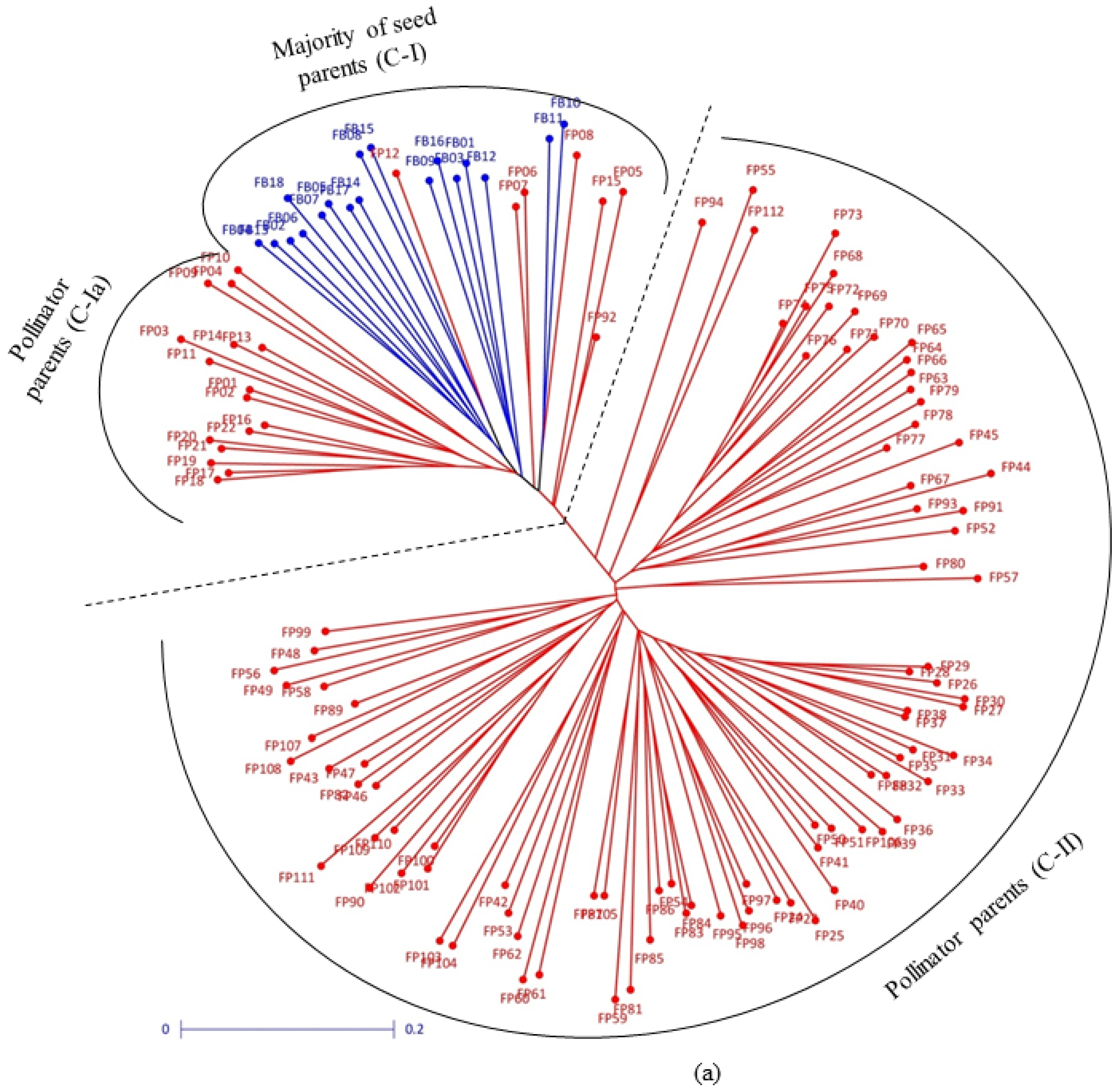
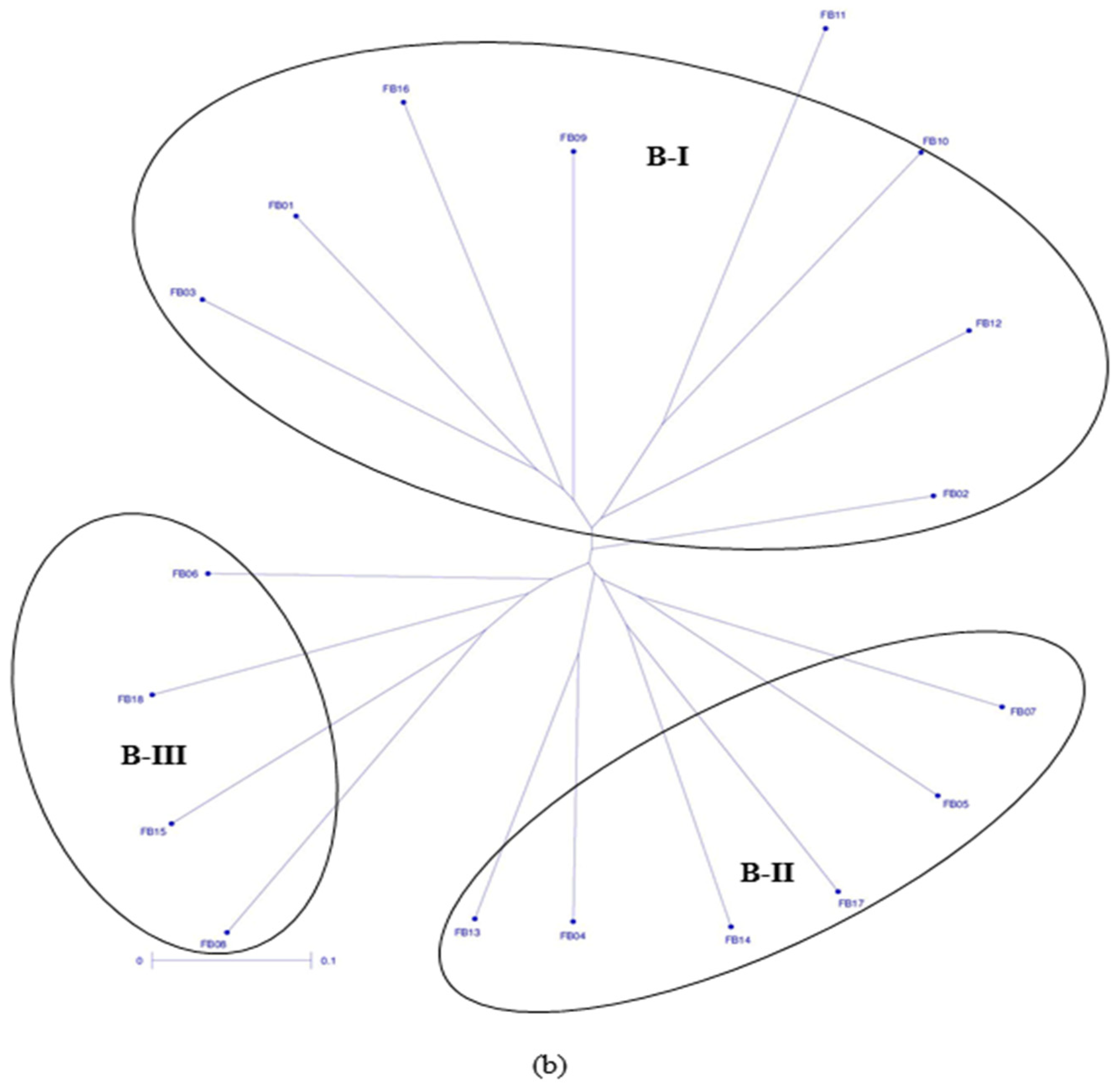
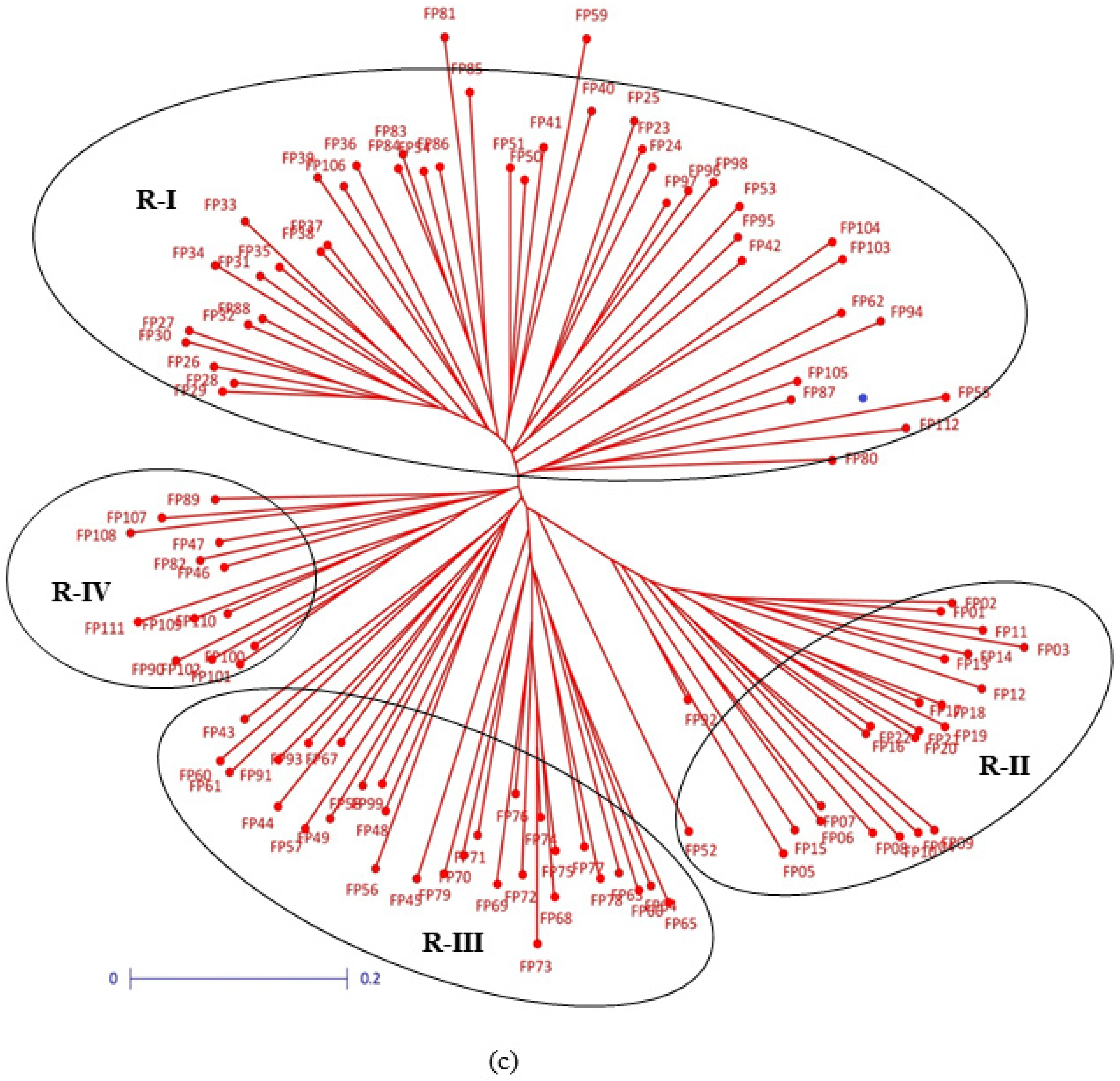
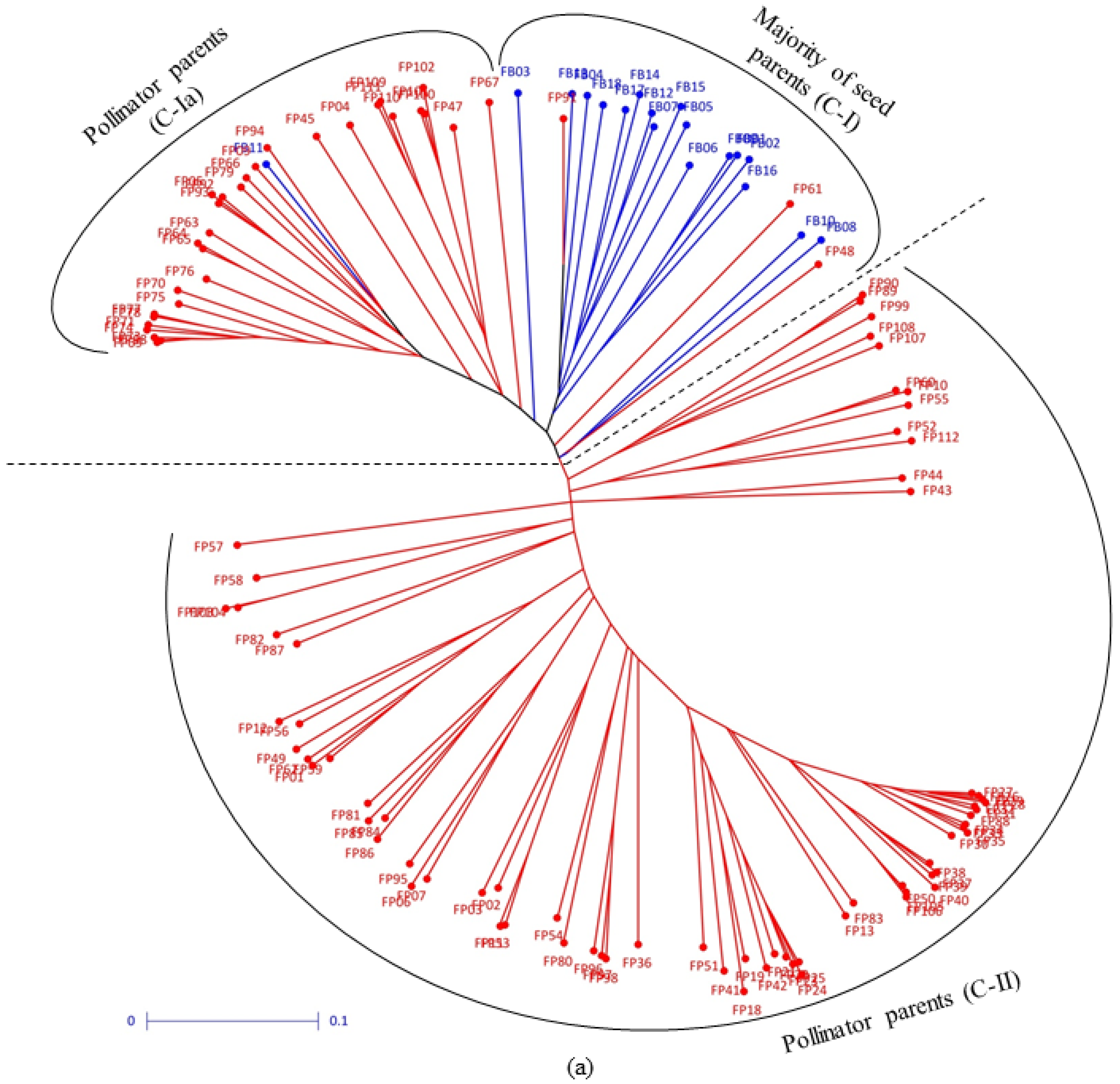
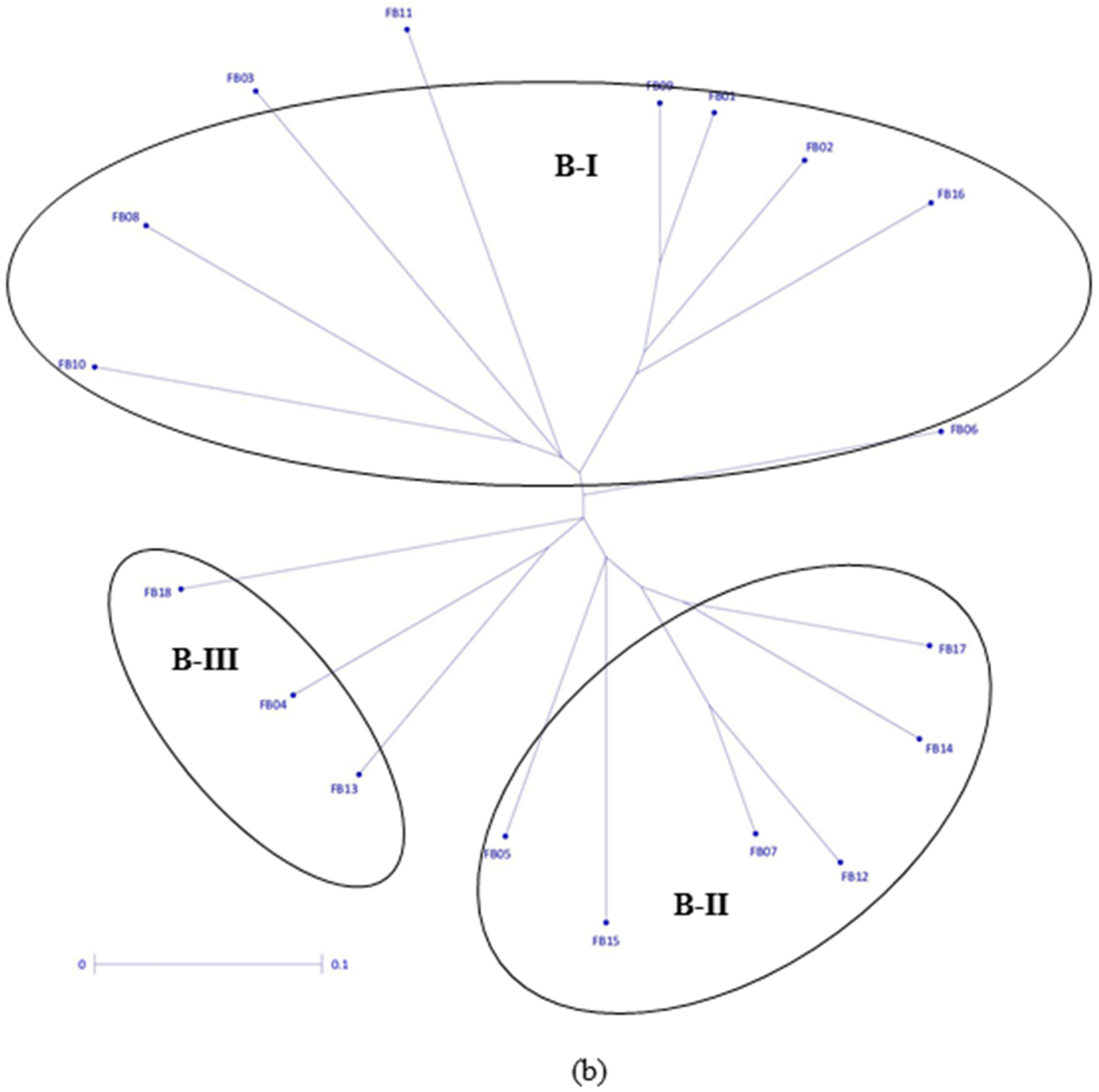
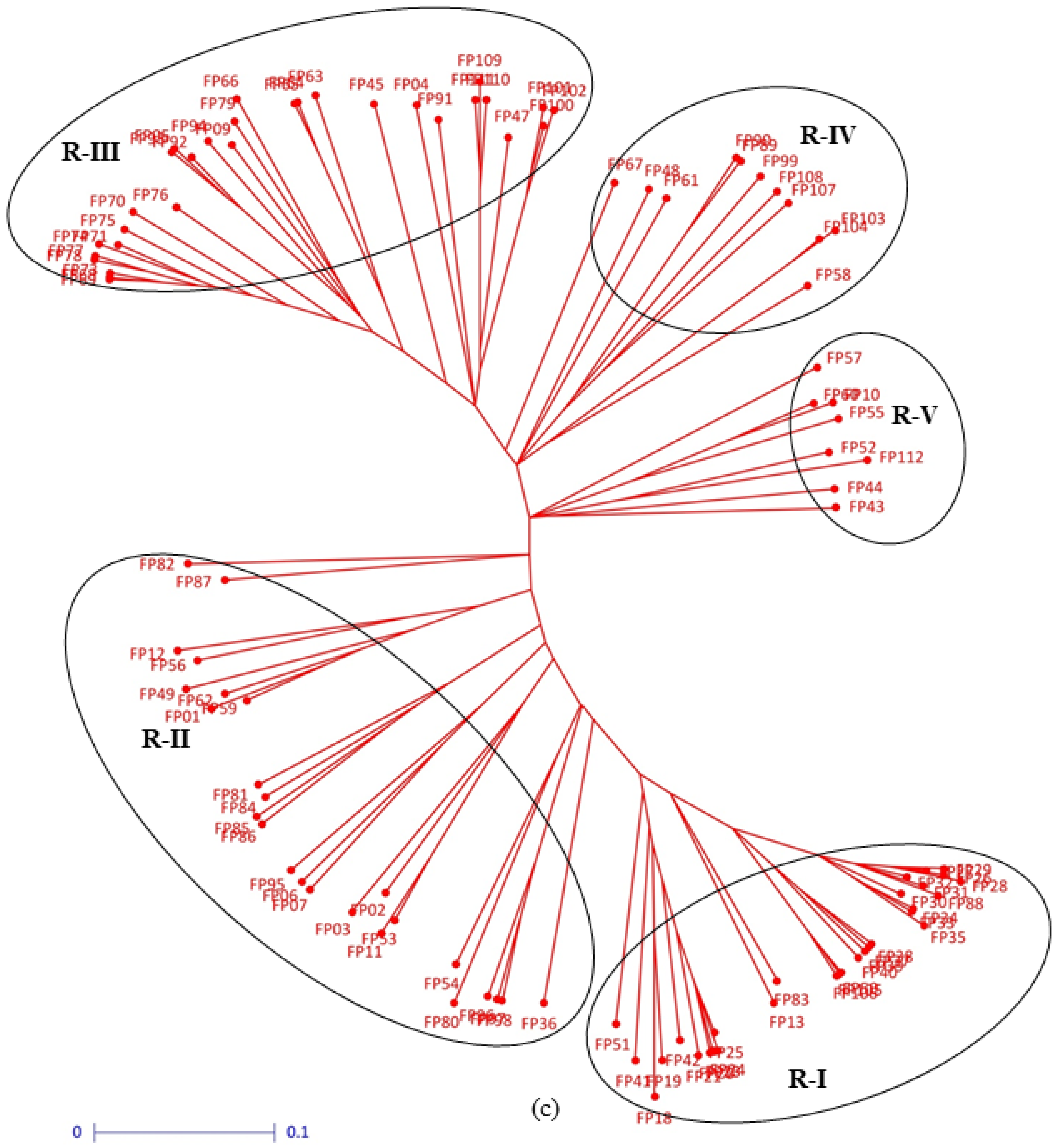
3.2. Clustering Pattern and Genetic Relatedness between Hybrid Parents Based on Markers
3.3. Variability for Forage Traits in Hybrid Parents
3.4. Trait Association with SSRs Based Clusters
3.5. ANOVA for Heterosis Estimation Trial
3.6. Magnitude of Heterosis and Its Association with GD
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| SSR | Simple sequence repeats |
| GBS | Genotyping by sequencing |
| SNP | Single nucleotide polymorphism |
| GFY | Green forage yield |
| CP | Crude protein |
| IVOMD | In vitro organic matter digestibility |
| NIRS | Near infra-red reflectance spectroscopy |
| GD | Genetic distance |
| DFY | Dry forage yield |
| ME | Metabolizable energy |
| ICRISAT | International Crops Research Institute for the Semi-Arid Tropics |
| FB | Forage B line |
| FP | Forage pollinator |
| CMS | Cytoplasmic male sterility |
| PCR | Polymerase chain reaction |
| UNEAK | Universal Network Enabled Analysis Kit |
| MAF | Minor allele frequency |
| TNAU | Tamil Nadu Agricultural University |
| AMOVA | Analysis of molecular variance |
| Fst | Fixation index |
| PCoA | Principal coordinate analysis |
| BLUP | Best Linear Unbiased Prediction |
| ANOVA | Analysis of variance |
| MPH | Mid-parent heterosis |
| BPH | Better parent heterosis |
| MP | Mid parent |
| BP | Better parent |
| F1 | First filial hybrid |
| PIC | Polymorphism information content |
| ICMB | ICRISAT millet B-line |
| ICMS | ICRISAT millet synthetic composite variety |
| MC | Medium composite |
| G × E | Genotype × Environment |
References
- Burton, G. History of hybrid development in pearl millet in Tifton. In Proceedings of the First Grain Pearl Millet Symposium, Tifton, GA, USA, 17–18 January 1995; pp. 5–8. [Google Scholar]
- Davis, A.J.; Dale, N.M.; Ferreira, F.J. Pearl millet as an alternative feed ingredient in broiler diets. J. Appl. Poultry Res. 2003, 12, 137–144. [Google Scholar] [CrossRef]
- Hanna, W. Improvement of millets: Emerging trends. In Proceedings of the 2nd International Crop Science Congress, New Delhi, India; Chopra, V.L., Singh, R.B., Varma, A., Eds.; Oxford & IBH Publishing Co. Pvt. Ltd.: New Delhi, India, 1996; pp. 139–146. [Google Scholar]
- Reddy, A.A.; Malik, D.; Singh, I.P.; Ardeshna, N.J.; Kundu, K.K.; Rao, P.; Gupta, S.K.; Sharma, R.; Gajanan, G.N. Demand and supply for pearl millet Grain and fodder by 2020 in Western India. Agric. Situat. India 2012, 68, 635–646. [Google Scholar]
- Amarender Reddy, A.; Yadav, O.; Dharm Pal Malik, S.I.; Ardeshna, N.; Kundu, K.; Gupta, S.K.; Sharma, R.; Sawarganokar, G.L.; Shyam, M.; Reddy, K.S. Utilization Pattern, Demand and Supply of Pearl Millet Grain and Fodder in Western India; Working Paper Series No. 37; ICRISAT: Patancheru, India, 2013. [Google Scholar]
- De Assis, R.L.; de Freitas, R.S.; Mason, S.C. Pearl millet production practices in Brazil: A review. Exp. Agric. 2018, 54, 699–718. [Google Scholar] [CrossRef]
- Dias-Martins, A.M.; Pessanha, K.L.; Pacheco, S.; Rodrigues, J.A.S.; Carvalho, C.W.P. Potential use of pearl millet (Pennisetum glaucum (L.) R. Br.) in Brazil: Food security, processing, health benefits and nutritional products. Food Res. Int. 2018, 109, 175–186. [Google Scholar] [CrossRef]
- IGFRI (Indian Grassland and Fodder Research Institute). Vision 2050. 2013. Available online: http://www.igfri.res.in/ (accessed on 28 August 2018).
- Rai, K.; Yadav, O.; Gupta, S.K.; Mahala, R. Emerging research priorities in pearl millet. J. SAT Agric. Res. 2012, 10, 1–5. [Google Scholar]
- Rai, K.N.; Reddy, B.V.S.; Dakheel, A.J. Forage potential of sorghum and pearl millet adapted to dry lands. In Proceedings of the Seventh International Conference on the Development of Drylands, Tehran, Iran, 14–17 September 2003; Beltagy, K.B., Saxena, Eds.; Sustainable Development and Management of Drylands in the Twenty-First Century; International Centre for Agricultural Research in the Dry Areas: Aleppo, Syria, 2005; pp. 243–251. [Google Scholar]
- Bidinger, F.; Blummel, M.; Hash, C.; Choudhary, S. Genetic enhancement for superior food-feed traits in a pearl millet (Pennisetum glaucum (L.) R. Br.) variety by recurrent selection. Anim. Nutr. Feed Technol. 2010, 10, 61–68. [Google Scholar]
- Gupta, S.; Ghouse, S.; Atkari, D.; Blümmel, M. Pearl millet with higher stover yield and better forage quality: Identification of new germplasm and cultivars. In Proceedings of the 3rd Conference of Cereal Biotechnology and Breeding/CBB3, Berlin, Germany, 2–4 November 2015. [Google Scholar]
- Stich, B.; Haussmann, B.I.; Pasam, R.; Bhosale, S.; Hash, C.T.; Melchinger, A.E.; Parzies, H.K. Patterns of molecular and phenotypic diversity in pearl millet [Pennisetum glaucum (L.) R. Br.] from West and Central Africa and their relation to geographical and environmental parameters. BMC Plant Biol. 2010, 10, 216. [Google Scholar] [CrossRef]
- Nepolean, T.; Gupta, S.; Dwivedi, S.; Bhattacharjee, R.; Rai, K.; Hash, C. Genetic diversity in maintainer and restorer lines of pearl millet. Crop Sci. 2012, 52, 2555–2563. [Google Scholar] [CrossRef]
- Gupta, S.K.; Nepolean, T.; Sankar, S.M.; Rathore, A.; Das, R.R.; Rai, K.N.; Hash, C.T. Patterns of molecular diversity in current and previously developed hybrid parents of pearl millet [Pennisetum glaucum (L.) R. Br.]. Am. J. Plant Sci. 2015, 6, 1697–1712. [Google Scholar] [CrossRef]
- Hu, Z.; Mbacké, B.; Perumal, R.; Guèye, M.C.; Sy, O.; Bouchet, S.; Vara Prasad, P.V.; Morris, G.P. Population genomics of pearl millet (Pennisetum glaucum (L.) R. Br.): Comparative analysis of global accessions and Senegalese landraces. BMC Genom. 2015, 16, 1048. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Nepolean, T.; Shaikh, C.G.; Rai, K.; Hash, C.T.; Das, R.R.; Rathore, A. Phenotypic and molecular diversity-based prediction of heterosis in pearl millet (Pennisetum glaucum L. (R.) Br.). Crop J. 2018, 6, 271–281. [Google Scholar] [CrossRef]
- Ramya, A.R.; Ahamed, M.; Satyavathi, C.T.; Rathore, A.; Katiyar, P.; Raj, A.G.B.; Kumar, S.; Gupta, R.; Mahendrakar, M.D.; Yadav, R.S.; et al. Towards defining heterotic gene pools in pearl millet [Pennisetum glaucum (L.) R. Br.]. Front. Plant Sci. 2018, 8, 1934. [Google Scholar] [CrossRef]
- Sattler, F.; Sanogo, M.; Kassari, I.; Angarawai, I.; Gwadi, K.; Dodo, H.; Haussmann, B.I.G. Characterization of West and Central African accessions from a pearl millet reference collection for agro-morphological traits and Striga resistance. Plant Genet. Resour.-C 2018, 16, 260–272. [Google Scholar] [CrossRef]
- Singh, S.; Gupta, S.; Thudi, M.; Das, R.R.; Vemula, A.; Garg, V.; Varshney, R.K.; Rathore, A.; Pahuja, S.K.; Yadav, D.V. Genetic Diversity Patterns and Heterosis Prediction Based on SSRs and SNPs in Hybrid Parents of Pearl Millet. Crop Sci. 2018, 58, 2379–2390. [Google Scholar] [CrossRef]
- Chowdari, K.; Venkatachalam, S.; Davierwala, A.; Gupta, V.; Ranjekar, P.; Govila, O. Hybrid performance and genetic distance as revealed by the (GATA) 4 microsatellite and RAPD markers in pearl millet. Theor. Appl. Genet. 1998, 97, 163–169. [Google Scholar] [CrossRef]
- Betran, F.; Beck, D.; Bänziger, M.; Edmeades, G. Genetic analysis of inbred and hybrid grain yield under stress and nonstress environments in tropical maize. Crop Sci. 2003, 43, 807–817. [Google Scholar] [CrossRef]
- Wegary, D.; Vivek, B.; Labuschagne, M. Association of parental genetic distance with heterosis and specific combining ability in quality protein maize. Euphytica 2013, 191, 205–216. [Google Scholar] [CrossRef]
- Cai, J.; Lan, W. Using of AFLP marker to predict the hybrid yield and yield heterosis in rice. Chin. Agric. Sci. Bull. 2005, 21, 39. [Google Scholar]
- Zhao, Q.Y.; Zhu, Z.; Zhang, Y.D.; Zhao, L.; Chen, T.; Zhang, Q.F.; Wang, C.L. Analysis on correlation between heterosis and genetic distance based on simple sequence repeat markers in japonica rice. Chin. J. Rice Sci. 2009, 23, 141–147. [Google Scholar]
- Darvishzadeh, R. Phenotypic and molecular marker distance as a tool for prediction of heterosis and F1 performance in sunflower (Helianthus annuus’ L.) under well-watered and water-stressed conditions. Aust. J. Crop Sci. 2012, 6, 732. [Google Scholar]
- Dreisigacker, S.; Melchinger, A.; Zhang, P.; Ammar, K.; Flachenecker, C.; Hoisington, D.; Warburton, M.L. Hybrid performance and heterosis in spring bread wheat, and their relations to SSR-based genetic distances and coefficients of parentage. Euphytica 2005, 144, 51–59. [Google Scholar] [CrossRef]
- Ndhlela, T.; Herselman, L.; Semagn, K.; Magorokosho, C.; Mutimaamba, C.; Labuschagne, M.T. Relationships between heterosis, genetic distances and specific combining ability among CIMMYT and Zimbabwe developed maize inbred lines under stress and optimal conditions. Euphytica 2015, 204, 635–647. [Google Scholar] [CrossRef]
- Xie, F.; He, Z.; Esguerra, M.Q.; Qiu, F.; Ramanathan, V. Determination of heterotic groups for tropical Indica hybrid rice germplasm. Theor. Appl. Genet. 2014, 127, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Qiu, F.; Larazo, W.; dela Paz, M.A.; Xie, F. Heterotic groups of tropical indica rice germplasm. Theor. Appl. Genet. 2015, 128, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Gvozdenović, S.; Saftić-Panković, D.; Jocić, S.; Radić, V. Correlation between heterosis and genetic distance based on SSR markers in sunflower (Helianthus annus L.). J. Agric. Sci. 2009, 54, 1–10. [Google Scholar]
- Burton, G.W. Pearl millets Tift 23DA and Tift 23DB released. Georgia Agric. Res. 1967, 9, 6. [Google Scholar]
- Burton, G.W. Registration of Pearl Millet Inbreds Tift 23B1, Tift 23A1, Tift 23DB1, and Tift 23DA11 (Reg. Nos. PL 1, PL 2, PL 3, and PL 4). Crop Sci. 1969, 9, 397. [Google Scholar] [CrossRef]
- Allouis, S.; Qi, X.; Lindup, S.; Gale, M.; Devos, K. Construction of a BAC library of pearl millet, Pennisetum glaucum. Theor. Appl. Genet. 2001, 102, 1200–1205. [Google Scholar] [CrossRef]
- Budak, H.; Pedraza, F.; Cregan, P.; Baenziger, P.; Dweikat, I. Development and utilization of SSRs to estimate the degree of genetic relationships in a collection of pearl millet germplasm. Crop Sci. 2003, 43, 2284–2290. [Google Scholar] [CrossRef]
- Qi, X.; Pittaway, T.; Lindup, S.; Liu, H.; Waterman, E.; Padi, F.; Hash, C.T.; Zhu, J.; Gale, M.D.; Devos, K.M. An integrated genetic map and a new set of simple sequence repeat markers for pearl millet, Pennisetum glaucum. Theor. Appl. Genet. 2004, 109, 1485–1493. [Google Scholar] [CrossRef]
- Rajaram, V.; Nepolean, T.; Senthilvel, S.; Varshney, R.K.; Vadez, V.; Srivastava, R.K.; Shah, T.M.; Supriya, A.; Kumar, S.; Kumari, B.R.; et al. Pearl millet [Pennisetum glaucum (L.) R. Br.] consensus linkage map constructed using four RIL mapping populations and newly developed EST-SSRs. BMC Genom. 2013, 14, 159. [Google Scholar] [CrossRef]
- Senthilvel, S.; Jayashree, B.; Mahalakshmi, V.; Kumar, P.S.; Nakka, S.; Nepolean, T.; Hash, C.T. Development and mapping of simple sequence repeat markers for pearl millet from data mining of expressed sequence tags. BMC Plant Biol. 2008, 8, 119. [Google Scholar] [CrossRef]
- Prasanth, V.; Chandra, S.; Hoisington, D.; Jayashree, B. AlleloBin: A Program for Allele Binning in Microsatellite Markers based on the Algorithm of Idury and Cardon (1997); International Crops Research Institute for the Semi-Arid Tropics (ICRISAT): New Delhi, India, 2006. [Google Scholar]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Lipka, A.E.; Glaubitz, J.; Elshire, R.; Cherney, J.H.; Casler, M.D.; Buckler, E.S.; Costich, D.E. Switchgrass genomic diversity, ploidy, and evolution: Novel insights from a network-based SNP discovery protocol. PLoS Genet. 2013, 9, e1003215. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Bidinger, F.; Blümmel, M. Determinants of ruminant nutritional quality of pearl millet [Pennisetum glaucum (L.) R. Br.] stover: I. Effects of management alternatives on stover quality and productivity. Field Crops Res. 2007, 103, 119–128. [Google Scholar] [CrossRef]
- Blümmel, M.; Bidinger, F.; Hash, C. Management and cultivar effects on ruminant nutritional quality of pearl millet (Pennisetum glaucum (L.) R. Br.) stover: II. Effects of cultivar choice on stover quality and productivity. Field Crops Res. 2007, 103, 129–138. [Google Scholar] [CrossRef]
- Liu, K.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Guan, R.; Liu, Z.; Ma, Y.; Wang, L.; Li, L.; Lin, F.; Luan, W.; Chen, P.; Yan, Z.; et al. Genetic structure and diversity of cultivated soybean (Glycine max (L.) Merr.) landraces in China. Theor. Appl. Genet. 2008, 117, 857–871. [Google Scholar] [CrossRef]
- Upadhyaya, H.D.; Dwivedi, S.L.; Baum, M.; Varshney, R.K.; Udupa, S.M.; Gowda, C.L.; Hoisington, D.; Singh, S. Genetic structure, diversity, and allelic richness in composite collection and reference set in chickpea (Cicer arietinum L.). BMC Plant Biol. 2008, 8, 106. [Google Scholar] [CrossRef]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [PubMed]
- Perrier, X.; Jacquemoud-Collet, J.P. DARwin Software. Available online: https://darwin.cirad.fr/ (accessed on 11 March 2017).
- Mantel, N. The detection of disease clustering and a generalized regression approach. Cancer Res. 1967, 27, 209–220. [Google Scholar] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: https://www.R-project.org/ (accessed on 21 July 2017).
- SAS Institute Inc. SAS/STAT® 14.1 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2017. [Google Scholar]
- Kapila, R.; Yadav, R.; Plaha, P.; Rai, K.; Yadav, O.; Hash, C.; Howarth, C.J. Genetic diversity among pearl millet maintainers using microsatellite markers. Plant Breed. 2008, 127, 33–37. [Google Scholar] [CrossRef]
- Sumanth, M.; Sumathi, P.; Vinodhana, N.; Sathya, M. Assessment of Genetic Distance Among the Inbred Lines of Pearl Millet (Pennisetum glaucum (L.) R. Br) Using SSR Markers. Int. J. Biotechnol. Allied Fields 2013, 1, 153–162. [Google Scholar]
- He, Z.; Xie, F.; Chen, L.; Paz, M.A.D. Genetic diversity of tropical hybrid rice germplasm measured by molecular markers. Rice Sci. 2012, 19, 193–201. [Google Scholar] [CrossRef]
- Filippi, C.V.; Aguirre, N.; Rivas, J.G.; Zubrzycki, J.; Puebla, A.; Cordes, D.; Moreno, M.V.; Fusari, C.M.; Alvarez, D.; Heinz, R.A.; et al. Population structure and genetic diversity characterization of a sunflower association mapping population using SSR and SNP markers. BMC Plant Biol. 2015, 15, 52. [Google Scholar] [CrossRef]
- Hamblin, M.T.; Warburton, M.L.; Buckler, E.S. Empirical comparison of simple sequence repeats and single nucleotide polymorphisms in assessment of maize diversity and relatedness. PLoS ONE 2007, 2, e1367. [Google Scholar] [CrossRef]
- Jones, E.; Sullivan, H.; Bhattramakki, D.; Smith, J. A comparison of simple sequence repeat and single nucleotide polymorphism marker technologies for the genotypic analysis of maize (Zea mays L.). Theor. Appl. Genet. 2007, 115, 361–371. [Google Scholar] [CrossRef]
- Akmal, M.; Naeem, M.; Nasim, S.; Shakoor, A. Performance of different pearl millet genotypes under rainfed conditions. J. Agric. Res. 1992, 30, 53–58. [Google Scholar]
- Byregowda, M. Performance of fodder bajra genotypes under rainfed conditions. Curr. Res.-Univ. Agric. Sci. 1990, 19, 128–129. [Google Scholar]
- Mohammad, D.; Hussain, A.; Khan, S.; Bhatti, M.B. Performance of new pearl millet cultivars. Pakistan J. Sci. Ind. R. 1993, 36, 261–263. [Google Scholar]
- Naeem, M.; Nasim, S.; Shakoor, A. Performance of new pearl millet varieties under rainfed conditions. J. Agric. Res. 1993, 31, 295–298. [Google Scholar]
- Rai, K.N.; Blümmel, M.; Singh, A.K.; Rao, A.S. Variability and relationships among forage yield and quality traits in pearl millet. Eur. J. Plant Sci. Biotechnol. 2012, 6, 118–124. [Google Scholar]
- Van Soest, P. Nitrogen metabolism. In Nutritional Ecology of the Ruminant, 2nd ed.; Comstock Publishing Associates-Cornell University Press: Ithaca, NY, USA, 1994; pp. 290–311. [Google Scholar]
- Blümmel, M.; Rai, K. Stover quality and grain yield relationships and heterosis effects in pearl millet. Int. Sorghum Millets Newsl. 2003, 44, 141–145. [Google Scholar]
- Hash, C.; Blummel, M.; Bidinger, F. Genotype x environment interactions in food-feed traits in pearl millet cultivars. Int. Sorghum Millets Newsl. 2006, 47, 153–157. [Google Scholar]
- Vinayan, M.; Babu, R.; Jyothsna, T.; Zaidi, P.; Blümmel, M. A note on potential candidate genomic regions with implications for maize stover fodder quality. Field Crops Res. 2013, 153, 102–106. [Google Scholar] [CrossRef]
- Zaidi, P.; Vinayan, M.; Blümmel, M. Genetic variability of tropical maize stover quality and the potential for genetic improvement of food-feed value in India. Field Crops Res. 2013, 153, 94–101. [Google Scholar] [CrossRef]
- Blümmel, M.; Deshpande, S.; Kholova, J.; Vadez, V. Introgression of staygreen QLT’s for concomitant improvement of food and fodder traits in Sorghum bicolor. Field Crops Res. 2015, 180, 228–237. [Google Scholar] [CrossRef]
- Kristjanson, P.; Zerbini, E.; Rao, K. Genetic Enhancement of Sorghum and Millet Residues Fed to Ruminants: An Ex Ante Assessment of Returns to Research; Impact Assessment Series No. 3; ILRI (aka ILCA and ILRAD): Nairobi, Kenya, 1999. [Google Scholar]
- Blümmel, M.; Rao, P.P. Economic value of sorghum stover traded as fodder for urban and peri-urban dairy production in Hyderabad, India. Int. Sorghum Millets Newsl. 2006, 47, 97–100. [Google Scholar]
- Melchinger, A.E.; Lee, M.; Lamkey, K.R.; Woodman, W.L. Genetic diversity for restriction fragment length polymorphisms: Relation to estimated genetic effects in maize inbreds. Crop Sci. 1990, 30, 1033–1040. [Google Scholar] [CrossRef]
- Charcosset, A.; Lefort-Busen, M.M.; Gallais, A. Relationship between herosis and heterozgosity at marker loci: A theoretical computation. Theor. Appl. Genet. 1991, 81, 571–575. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Allelic Richness † | Gene Diversity † | Heterozygosity † | PIC † |
|---|---|---|---|---|
| SSRs | ||||
| Seed parents | 4.63 (1.00–13.00) | 0.55 (0.00–0.90) | 0.02 (0.00–0.14) | 0.52 (0.00–0.89) |
| Pollinator parents | 9.81 (2.00–29.00) | 0.66 (0.05–0.90) | 0.03 (0.00–0.17) | 0.63 (0.05–0.93) |
| Seed and Pollinator parents | 10.6 (2.00–31.00) | 0.68 (0.06–0.94) | 0.02 (0.00–0.12) | 0.65 (0.06–0.94) |
| Standard error | 0.90 (0.35–0.92) | 0.03 (0.03–0.03) | 0.00 (0.01–0.01) | 0.03 (0.03–0.03) |
| GBS-Identified SNPs | ||||
| Seed parents | 1.95 (1.00–2.00) | 0.37 (0.00–0.50) | 0.13 (0.00–1.00) | 0.29 (0.00–0.38) |
| Pollinator parents | 2.00 (1.00–2.00) | 0.48 (0.00–0.50) | 0.15 (0.00–0.96) | 0.37 0.00–0.38) |
| Seed and Pollinator parents | 2.00 (2.00–2.00) | 0.48 (0.02–0.50) | 0.15 (0.00–0.97) | 0.37 (0.02–0.38) |
| Standard error | 0.00 (0.00–0.00) | 0.00 (0.00–0.00) | 0.00 (0.00–0.00) | 0.00 (0.00–0.00) |
| Traits | Lines | First Cut | Second Cut | ||
|---|---|---|---|---|---|
| Mean | Range | Mean | Range | ||
| Green fodder yield (t ha−1) | Seed parents | 20.32 | 16.30–22.89 | 19.79 | 12.35–26.21 |
| Pollinator parents | 22.21 | 15.18–29.16 | 28.03 | 15.61–42.42 | |
| Dry fodder yield (t ha−1) | Seed parents | 4.25 | 3.63–5.19 | 6.21 | 5.23–8.99 |
| Pollinator parents | 4.42 | 3.34–5.91 | 6.31 | 5.06–8.09 | |
| Crude protein (%) | Seed parents | 12.92 | 11.94–14.61 | 11.86 | 11.40–12.44 |
| Pollinator parents | 12.55 | 11.49–13.52 | 11.71 | 11.04–12.51 | |
| In vitro organic matter digestibility (%) | Seed parents | 54.66 | 53.93–55.40 | 52.67 | 51.16–54.26 |
| Pollinator parents | 54.81 | 53.78–55.63 | 52.58 | 51.01–55.04 | |
| Lines | Cluster No. | First Cut | Second Cut | ||||||
|---|---|---|---|---|---|---|---|---|---|
| GFY (t ha−1) † | DFY (t ha−1) † | CP (%) † | IVOMD (%) † | GFY (t ha−1) † | DFY (t ha−1) † | CP (%) † | IVOMD (%) † | ||
| Seed parents | B-I | 20.14 (17.13, 22.39) | 4.21 (3.63, 4.46) | 12.75 (11.94, 13.47) | 54.69 (54.04, 55.40) | 18.72 (12.35, 22.02) | 6.31 (5.23, 8.99) | 11.82 (11.56, 12.13) | 52.90 (51.80, 54.26) |
| B-II | 19.67 (16.30, 22.89) | 4.10 (3.70, 4.54) | 13.07 (12.22, 14.61) | 54.79 (54.47, 55.03) | 20.53 (14.57, 26.21) | 6.16 (5.98, 6.43) | 11.82 (11.40, 12.44) | 52.68 (52.01, 53.70) | |
| B-III | 21.63 (20.60, 22.33) | 4.54 (3.87, 5.19) | 13.02 (12.57, 13.49) | 54.43 (53.93, 55.06) | 20.84 (17.30, 23.81) | 6.09 (6.02, 6.25) | 12.01 (11.81, 12.13) | 52.18 (51.16, 53.30) | |
| Pollinator parents | R-I | 22.81 (18.44, 27.40) | 4.46 (3.64, 5.61) | 12.40 (11.49, 13.18) | 54.79 (53.78, 55.56) | 29.73 (15.61, 37.30) | 6.44 (5.47, 7.69) | 11.67 (11.04, 12.36) | 52.41 (51.01, 53.83) |
| R-II | 21.22 (15.18, 25.49) | 4.29 (3.41, 4.80) | 12.75 (12.23, 13.52) | 54.85 (54.23, 55.47) | 26.36 (16.35, 34.75) | 6.21 (5.06, 6.94) | 11.69 (11.21, 12.04) | 52.60 (51.33, 53.69) | |
| R-III | 22.24 (17.48, 29.16) | 4.42 (3.69, 5.91) | 12.64 (11.77, 13.42) | 54.85 (54.03, 55.63) | 27.99 (19.56, 42.42) | 6.28 (5.31, 8.09) | 11.78 (11.13, 12.51) | 52.77 (51.60, 55.04) | |
| R-IV | 21.97 (19.89, 24.07) | 4.53 (3.34, 5.51) | 12.52 (11.55, 13.12) | 54.68 (54.21, 55.15) | 23.29 (18.83, 28.89) | 6.06 (5.17, 7.21) | 11.72 (11.35, 12.38) | 52.91 (51.85, 54.27) | |
| Source of Variation | DF | GFY | DFY | CP | IVOMD | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| † FC | ǂ SC | FC | SC | FC | SC | FC | SC | FC | SC | |
| Locations | 1 | 1 | 5.71 * | 309.26 *** | 277.36 *** | 97.57 *** | 116.88 *** | 25.07 *** | 562.89 *** | 987.69 *** |
| Replication (Locations) | 3 | 3 | 4.43 * | 2.63 | 6.85 *** | 1.27 | 3.47 * | 5.57 ** | 0.29 | 6.00 ** |
| Treatments | 97 | 58 | 6.18 *** | 11.08 *** | 2.58 *** | 3.44 *** | 4.73 *** | 2.56 *** | 1.55 * | 3.59 *** |
| Parents | 17 | 13 | 3.37 *** | 5.98 *** | 0.11 | 1.65 | 6.98 *** | 3.00 *** | 1.64 | 6.17 *** |
| (i) Females | 9 | 8 | 1.12 | 2.86 | 0.31 | 0.68 | 8.08 *** | 3.18 *** | 2.21 * | 3.71 *** |
| (ii) Males | 7 | 4 | 21.19 *** | 9.15 *** | 1.22 | 3.02 * | 4.42 *** | 2.35 | 1.12 | 5.15 *** |
| (iii) Females vs. Males | 1 | 1 | 264.05 *** | 21.50 *** | 141.81 *** | 4.37 * | 12.99 *** | 3.35 | 0.22 | 33.7 *** |
| Hybrids | 79 | 44 | 11.13 *** | 8.08 *** | 1.42 | 2.57 *** | 2.65 *** | 1.65 * | 1.44 * | 2.88 *** |
| (i) Lines | 9 | 8 | 6.96 | 2.16 * | 0.96 | 1.65 | 3.63 *** | 1.48 | 3.13 *** | 4.84 *** |
| (ii) Testers | 7 | 4 | 0.97 | 21.82 *** | 1.05 | 11.53 *** | 6.92 *** | 6.04 *** | 2.06 * | 4.94 *** |
| (iii) Lines × Testers | 63 | 32 | 3.49 *** | 7.04 *** | 0.23 | 1.64 * | 2.15 *** | 1.04 | 1.24 | 2.17 *** |
| Hyb vs. Par | 1 | 1 | 1.46 * | 190.82 *** | 1.83 *** | 61.11 *** | 108.76 *** | 29.97 *** | 7.70 ** | 1.77 |
| Location × Treatments | 97 | 58 | 2.76 *** | 6.12 *** | 1.09 | 1.84 *** | 2.48 *** | 1.68 * | 1.76 *** | 2.95 *** |
| Location × Parents | 17 | 13 | 0.87 | 4.08 *** | 0.21 | 0.96 | 2.47 *** | 1.29 | 0.75 | 4.02 *** |
| (i) Location × Females | 9 | 8 | 0.94 | 3.54 *** | 0.08 | 0.92 | 3.29 *** | 0.89 | 0.41 | 4.59 *** |
| (ii) Location × Males | 7 | 4 | 0.55 | 6.03 *** | 0.4 | 1.28 | 1.67 | 1.65 | 1.17 | 0.82 |
| (iii) Location × (Lines vs. Testers) | 1 | 1 | 2.37 | 1 | 0.03 | 0.01 | 1.07 | 3.51 | 0.94 | 13.35 *** |
| Location × Hybrids | 79 | 44 | 1.22 | 5.71 *** | 1.03 | 2.01 *** | 2.32 *** | 1.02 | 1.33 | 2.42 *** |
| (i) Location × Lines | 9 | 8 | 1.01 | 1.41 | 0.74 | 0.96 | 2.53 ** | 0.96 | 2.58 ** | 2.39 * |
| (ii) Location × Testers | 7 | 4 | 0.91 | 10.24 *** | 0.31 | 7.35 *** | 2.89 ** | 3.80 ** | 0.81 | 0.75 |
| (iii) Location × (Lines × Testers) | 63 | 32 | 1.29 | 5.82 *** | 1.14 | 1.53 * | 2.27 *** | 0.64 | 1.19 | 2.59 *** |
| Location × (Hybrids vs. Parents | 1 | 1 | 28.05 *** | 45.02 *** | 79.69 *** | 6.21 ** | 12.76 *** | 34.65 *** | 50.91 *** | 16.97 *** |
| Traits | Cutting Intervals | Mid-Parent Heterosis (%) | Better-Parent Heterosis (%) | ||||
|---|---|---|---|---|---|---|---|
| Minimum | Maximum | Average | Minimum | Maximum | Average | ||
| GFY | First cut | 20.6 | 115.7 | 58.2 | −8.2 | 74.4 | 32.6 |
| Second cut | −22.2 | 378.1 | 103.8 | −37.0 | 301.9 | 51.5 | |
| DFY | First cut | 33.1 | 406.5 | 189.1 | −13.1 | 344.3 | 154.2 |
| Second cut | −15.5 | 290.0 | 93.2 | −29.7 | 248.2 | 50.7 | |
| CP | First cut | −23.6 | 9.8 | −9.5 | −27.8 | 8.8 | −14.4 |
| Second cut | −24.1 | 7.2 | −7.8 | −30.7 | 4.5 | −12.8 | |
| IVOMD | First cut | −6.8 | 7.1 | 1.8 | −8.2 | 6.7 | 0.1 |
| Second cut | −7.5 | 5.9 | 0.0 | −10.8 | 3.4 | −2.9 | |
| Traits | Cutting Intervals | Correlation Coefficient between GD and MPH | Correlation Coefficient between GD and BPH | ||
|---|---|---|---|---|---|
| SSRs | SNPs | SSRs | SNPs | ||
| Green forage yield (GFY, t ha−1) | First cut | 0.11 | −0.02 | 0.12 | −0.15 |
| Second cut | 0.34 * | 0.15 | 0.41 * | 0.13 | |
| Dry forage yield (DFY, t ha−1) | First cut | −0.10 | 0.32 ** | −0.14 | 0.28 * |
| Second cut | 0.25 | 0.19 | 0.33 * | 0.19 | |
| Crude protein (CP, %) | First cut | 0.26 * | 0.04 | 0.20 | −0.12 |
| Second cut | −0.05 | −0.22 | −0.02 | −0.19 | |
| In vitro organic matter digestibility (IVOMD, %) | First cut | 0.11 | 0.00 | 0.18 | −0.01 |
| Second cut | −0.18 | 0.06 | −0.17 | 0.11 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ponnaiah, G.; Gupta, S.K.; Blümmel, M.; Marappa, M.; Pichaikannu, S.; Das, R.R.; Rathore, A. Utilization of Molecular Marker Based Genetic Diversity Patterns in Hybrid Parents to Develop Better Forage Quality Multi-Cut Hybrids in Pearl Millet. Agriculture 2019, 9, 97. https://doi.org/10.3390/agriculture9050097
Ponnaiah G, Gupta SK, Blümmel M, Marappa M, Pichaikannu S, Das RR, Rathore A. Utilization of Molecular Marker Based Genetic Diversity Patterns in Hybrid Parents to Develop Better Forage Quality Multi-Cut Hybrids in Pearl Millet. Agriculture. 2019; 9(5):97. https://doi.org/10.3390/agriculture9050097
Chicago/Turabian StylePonnaiah, Govintharaj, Shashi Kumar Gupta, Michael Blümmel, Maheswaran Marappa, Sumathi Pichaikannu, Roma Rani Das, and Abhishek Rathore. 2019. "Utilization of Molecular Marker Based Genetic Diversity Patterns in Hybrid Parents to Develop Better Forage Quality Multi-Cut Hybrids in Pearl Millet" Agriculture 9, no. 5: 97. https://doi.org/10.3390/agriculture9050097
APA StylePonnaiah, G., Gupta, S. K., Blümmel, M., Marappa, M., Pichaikannu, S., Das, R. R., & Rathore, A. (2019). Utilization of Molecular Marker Based Genetic Diversity Patterns in Hybrid Parents to Develop Better Forage Quality Multi-Cut Hybrids in Pearl Millet. Agriculture, 9(5), 97. https://doi.org/10.3390/agriculture9050097