Multi-Trait Diverse Germplasm Sources from Mini Core Collection for Sorghum Improvement


 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of Materials
2.2. Experimental Details
2.3. Statistical Analysis
2.4. Trait-Specific Sources and Their Agronomic Performance
3. Results
3.1. Analysis of Variance
3.2. Trait-Specific Sources and Their Agronomic Performance
3.2.1. Agronomic Traits
3.2.2. Grain Nutritional Contents
3.2.3. Bioenergy Traits
3.2.4. Biotic Stress Tolerance
3.2.5. Abiotic Stress Tolerance
3.3. Multi-Trait-Specific Sources
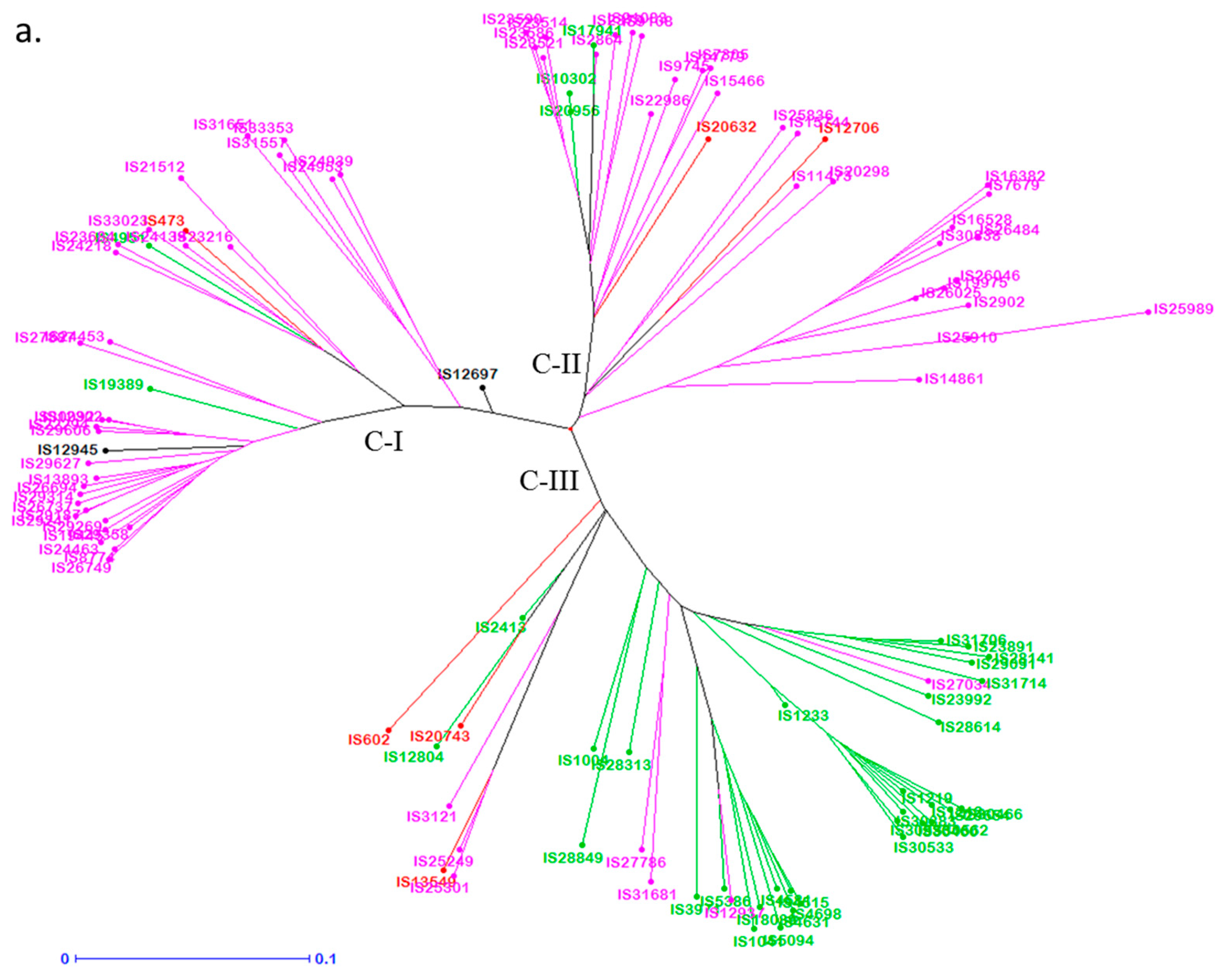
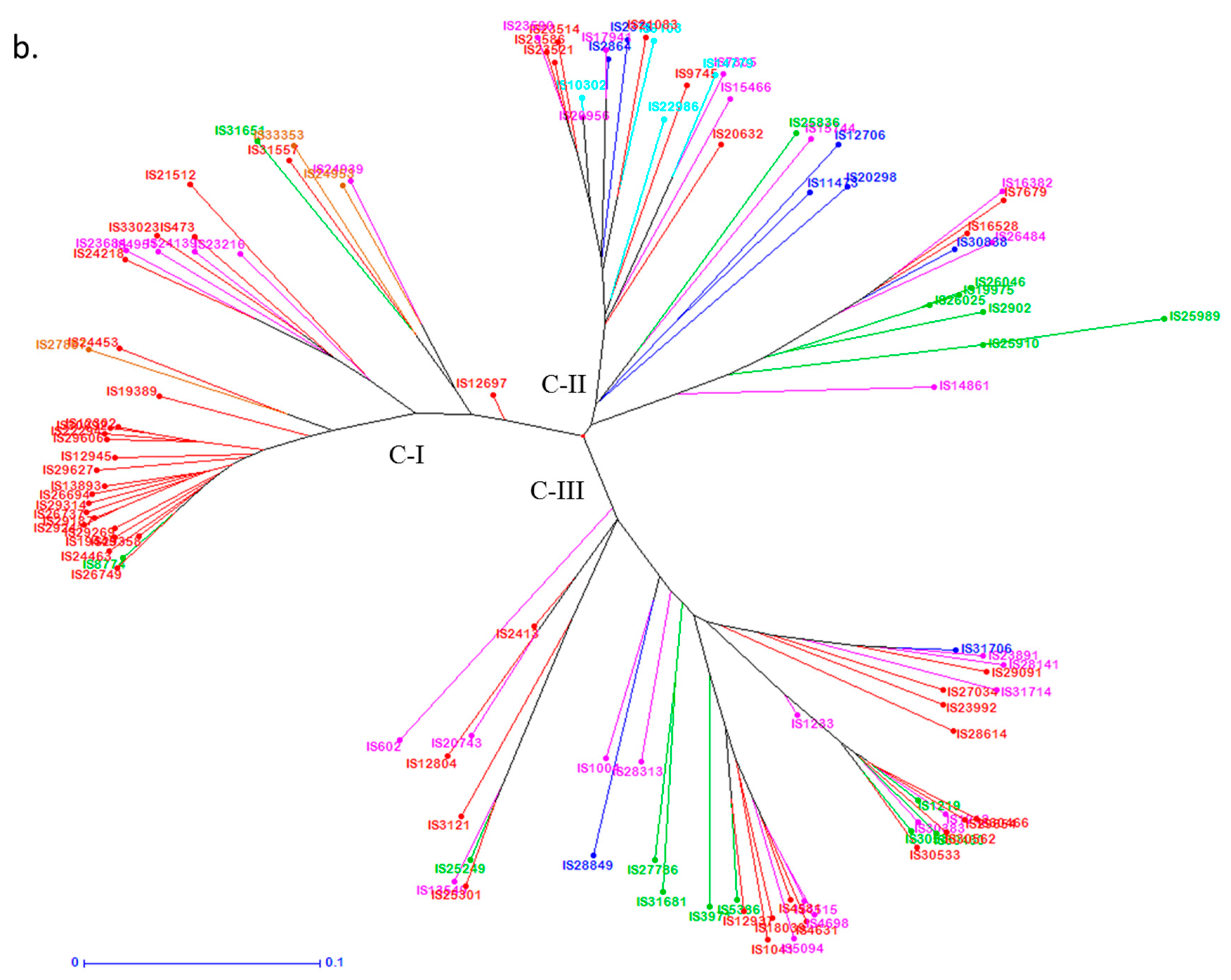
3.4. Genetic Distance Among Trait-Specific Sources
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Muraya, M.M.; de Villiers, S.; Parzies, H.K.; Mutegi, E.; Sagnard, F.; Kanyenji, B.M.; Kiambi, D.; Geiger, H.H. Genetic Structure and Diversity of Wild Sorghum Populations (Sorghum Spp.) from Different Eco-Geographical Regions of Kenya. Theor. Appl. Genet. 2011, 123, 571–583. [Google Scholar] [CrossRef]
- Mace, E.S.; Tai, S.; Gilding, E.K.; Li, Y.; Prentis, P.J.; Bian, L.; Campbell, B.C.; Hu, W.; Innes, D.J.; Han, X.; et al. Whole-Genome Sequencing Reveals Untapped Genetic Potential in Africa’s Indigenous Cereal Crop Sorghum. Nat. Commun. 2013, 4, 2320. [Google Scholar] [CrossRef]
- Murray, S.C.; Rooney, W.L.; Hamblin, M.T.; Mitchell, S.E.; Kresovich, S. Sweet Sorghum Genetic Diversity and Association Mapping for Brix and Height. Plant Genome J. 2009, 2, 48–62. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.; Primomo, V.; Monk, R.; Nelson, B.; Jons, E.; Porter, K. Genetic Diversity of Widely Used U.S. Sorghum Hybrids 1980–2008. Crop Sci. 2010, 50, 1664–1673. [Google Scholar] [CrossRef]
- Jordan, D.R.; Tao, Y.Z.; Godwin, I.D.; Henzell, R.G.; Cooper, M.; McIntyre, C.L. Loss of Genetic Diversity Associated with Selection for Resistance to Sorghum Midge in Australian Sorghum. Euphytica 1998, 102, 1–7. [Google Scholar] [CrossRef]
- Jordan, D.R.; Tao, Y.; Godwin, I.D.; Henzell, R.G.; Cooper, M.; McIntyre, C.L. Prediction of Hybrid Performance in Grain Sorghum Using RFLP Markers. Theor. Appl. Genet. 2003, 106, 559–567. [Google Scholar] [CrossRef]
- Keneni, G.; Bekele, E.; Imtiaz, M.; Dagne, K. Genetic Vulnerability of Modern Crop Cultivars: Causes, Mechanism and Remedies. Int. J. Plant Res. 2012, 2, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Flath, K.; Miedaner, T.; Jin, Y.; Olivera, P.D.; Rouse, M.N. Genes for wheat stem rust resistance postulated in German cultivars and their efficacy in seedling and adult-plant field tests. Plant Breeding 2018, 3, 301–312. [Google Scholar] [CrossRef]
- Lewis, C.M.; Persoons, A.; Bebber, D.P.; Kigathi, R.N.; Maintz, J.; Findlay, K.; Bueno-sancho, V.; Corredor-moreno, P.; Harrington, S.A.; Kangara, N.; et al. Potential for re-emergence of wheat stem rust in the United Kingdom. Commun. Biol. 2018, 13. [Google Scholar] [CrossRef]
- Upadhyaya, H.D.; Dwivedi, S.L.; Vadez, V.; Hamidou, F.; Singh, S.; Varshney, R.K.; Liao, B. Multiple Resistant and Nutritionally Dense Germplasm Identified from Mini Core Collection in Peanut. Crop Sci. 2014, 54, 679–693. [Google Scholar] [CrossRef] [Green Version]
- Upadhyaya, H.D.; Pundir, R.P.S.; Dwivedi, S.L.; Gowda, C.L.L.; Reddy, V.G.; Singh, S. Developing a Mini Core Collection of Sorghum for Diversified Uilization of Germplasm. Crop Sci. 2009, 49, 1769–1780. [Google Scholar] [CrossRef]
- Sharma, R.; Rao, V.P.; Upadhyaya, H.D.; Reddy, V.G.; Thakur, R.P. Resistance to Grain Mold and Downy Mildew in a Mini-Core Collection of Sorghum Sermplasm. Plant Dis. 2010, 94, 439–444. [Google Scholar] [CrossRef]
- Sharma, R.; Upadhyaya, H.D.; Manjunatha, S.V.; Rao, V.P.; Thakur, R.P. Resistance to Foliar Diseases in a Mini-Core Collection of Sorghum Germplasm. Plant Dis. 2012, 96, 1629–1633. [Google Scholar] [CrossRef] [Green Version]
- Radwan, G.L.; Perumal, R.; Isakeit, T.; Magill, C.W.; Prom, L.K.; Little, C.R. Screening Exotic Sorghum Germplasm, Hybrids, and Elite Lines for Resistance to a New Virulent Pathotype (P6) of Peronosclerospora Sorghi Causing Downy Mildew. Plant Heal. Prog. 2011. [Google Scholar] [CrossRef]
- Borphukan, B. Evaluation of Minicore Germplasm of Rabi Sorghum for Charcoal Rot Resistance and Yield Component Traits, Expression Analysis of Selected R-Genes during Charcoal Rot Disease Incidence. Master’s Thesis, University of Agricultural Sciences, Dharwad, India, 2014. [Google Scholar]
- Seifers, D.L.; Perumal, R.; Little, C.R. New Sources of Resistance in Sorghum (Sorghum Bicolor) Germplasm Are Effective against a Diverse Array of Potyvirus Spp. Plant Dis. 2012, 96, 1775–1779. [Google Scholar] [CrossRef]
- Kapanigowda, M.H.; Perumal, R.; Djanaguiraman, M.; Aiken, R.M.; Tesso, T.; Prasad, P.V.; Little, C.R. Genotypic Variation in Sorghum [Sorghum Bicolor (L.) Moench] Exotic Germplasm Collections for Drought and Disease Tolerance. SpringerPlus 2013, 2, 650. [Google Scholar] [CrossRef]
- Upadhyaya, H.D.; Dwivedi, S.L.; Vetriventhan, M.; Krishnamurthy, L.; Singh, S.K. Post-Flowering Drought Tolerance Using Managed Stress Trials, Adjustment to Flowering, and Mini Core Collection in Sorghum. Crop Sci. 2017, 57, 310–321. [Google Scholar] [CrossRef]
- Upadhyaya, H.D.; Wang, Y.-H.; Dintyala, S.V.; Dwivedi, S.L.; Prasad, P.V.V.; Burrell, A.M.; Klein, R.; Morris, G.P.; Klein, P.E. Association Mapping of Germinability and Seedling Vigor in Sorghum under Controlled Low Temperature Conditions. Genome 2016, 59, 137–145. [Google Scholar] [CrossRef]
- Upadhyaya, H.D.; Dwivedi, S.L.; Singh, S.; Sahrawat, K.L.; Singh, S.K. Genetic Variation and Postflowering Drought Effects on Seed Iron and Zinc in ICRISAT Sorghum Mini Core Collection. Crop Sci. 2016, 56, 374–383. [Google Scholar] [CrossRef]
- Upadhyaya, H.D.; Dwivedi, S.L.; Ramu, P.; Singh, S.K.; Singh, S. Genetic Variability and Effect of Postflowering Drought on Stalk Sugar Content in Sorghum Mini Core Collection. Crop Sci. 2014, 54, 2120–2130. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Poudel, D.D.; Hasenstein, K.H. Identification of SSR Markers Associated with Saccharification Yield Using Pool-Based Genome-Wide Association Mapping in Sorghum. Genome 2011, 54, 883–889. [Google Scholar] [CrossRef]
- Upadhyaya, H.D.; Vetriventhan, M. Ensuring the Genetic Diversity of Sorghum. In Achieving Sustainable Cultivation of Sorghum; Roony, W., Ed.; Burleigh Dodds Science Publishing Limited: Cambridge, UK, 2018; Volume 1, p. 546. [Google Scholar]
- Sharma, H.C.; Taneja, S.L.; Rao, N.K.; Rao, K.E.P. Evaluation of Sorghum Germplasm for Resistance to Insect Pests; Information Bulletin No. 63; International Crops Research Institute for the Semi-Arid Tropics: Patancheru, India, 2003. [Google Scholar]
- El-Swaify, S.A.; Pathak, P.; Rego, T.J.; Singh, S. Soil Management for Optimized Productivity Under Rainfed Conditions in the Semi-Arid Tropics. In Advances in Soil Science; Steward, B.A., Ed.; Springer: New York, NY, USA, 1985; Volume 1, pp. 1–64. [Google Scholar]
- Bidinger, F.R.; Mahalakshimi, V.; Rao, G.D.P. Assessment of Drought Resistance in Pearl Millet [ Pennisetum americanum (L.) Leeke]. I Factors Affecting Yields under Stress. Aust. J. Agric. Res. 1987, 38, 37–48. [Google Scholar] [CrossRef]
- Wang, Y.; Upadhyaya, H.D.; Burrell, A.M.; Sahraeian, S.M.E.; Klein, R.R.; Klein, P.E. Genetic Structure and Linkage Disequilibrium in a Diverse, Representative Collection of the C4 Model Plant, Sorghum Bicolor. G3 Genes Genomes Genet. 2013, 3, 783–793. [Google Scholar] [CrossRef]
- Tajima, F.; Nei, M. Estimation Nucleotide of Evolutionary Distance between Nucleotide Sequences. Mol. Biol. Evol. 1984, 1, 269–285. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The Neighbour-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Perrier, X.; Jacquemoud-Collet, J.P. DARwin Software. Available online: http://darwin.cirad.fr/darwin (accessed on 5 August 2018).
- Morris, G.P.; Ramu, P.; Deshpande, S.P.; Hash, C.T.; Shah, T.; Upadhyaya, H.D.; Riera-Lizarazu, O.; Brown, P.J.; Acharya, C.B.; Mitchell, S.E.; et al. Population Genomic and Genome-Wide Association Studies of Agroclimatic Traits in Sorghum. Proc. Natl. Acad. Sci. USA 2013, 110, 453–458. [Google Scholar] [CrossRef]
- Vadez, V.; Krishnamurthy, L.; Hash, C.T.; Upadhyaya, H.D.; Borrell, A.K. Yield, Transpiration Efficiency, and Water-Use Variations and Their Interrelationships in the Sorghum Reference Collection. Crop Pasture Sci. 2011, 62, 645–655. [Google Scholar] [CrossRef]
- Jordan, D.R.; Mace, E.S.; Cruickshank, A.W.; Hunt, C.H.; Henzell, R.G. Exploring and Exploiting Genetic Variation from Unadapted Sorghum Germplasm in a Breeding Program. Crop Sci. 2011, 51, 1444–1457. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Trait | Mini Core Accession | Reference |
|---|---|---|
| Grain nutritional traits | ||
| Fe, 40.3 to 48.6 mg kg−1 seed | IS# 16382, 23992, 28313, 28389, 28849, 20743, 21645, 21863, 28747, 30508, 31681 | Upadhyaya et al. [20] |
| Zn, 32.2 to 36.4 mg kg−1 seed | IS# 30460, 602, 17980, 19859, 28451, 30466, 30536, 5301, 8774, 4951, 25249, 24139, 24175, 24218 | Upadhyaya et al. [20] |
| Fe, 40.8 to 48.9 mg kg−1 seed and Zn, 32.8 to 42.6 mg kg−1 seed | IS# 1219, 1233, 30450, 30507, 1212, 27786, 30383, 31651, 24503 | Upadhyaya et al. [20] |
| Protein (12.2 to 13.8%) | IS# 2902, 4951, 19975, 23684, 25249, 25910, 25989, 26025, 26046 | http://genebank.icrisat.org/ |
| Lysine (3.1 to 4.3%) | IS# 3971, 25836, 5386 | http://genebank.icrisat.org/ |
| Bioenergy traits | ||
| Stalk sugar content (Brix: 14.0 to 15.2%) | IS# 13294, 13549, 23216, 23684, 24139, 24939, 24953 | Upadhyaya et al. [21] |
| Dual purpose (grain and sweet stalk) | IS# 1004, 4698, 23891, 28141 | Upadhyaya et al. [21] |
| High saccharification yield | IS# 2872, 27887, 19262, 3158, 7305, 33353, 4951 | Wang et al. [22] |
| Biotic stresses | ||
| Downy mildew | IS# 28747, 31714, 23992, 27697, 28449, 30400; 1212, 2413, 3121, 4060, 4360, 4372, 4613, 4631, 5094, 7305, 9745, 12302, 12804, 12883, 12965, 13549, 15170, 15478, 15945, 16528, 20625, 20632, 21083, 22294, 22720, 23216, 24453, 24462, 24463, 26222, 26484, 26617, 26749, 27557, 29239, 29314, 29358, 29392, 29606, 29627, 29654, 30092, 30383, 30443, 30466, 30562, 31557 | Sharma et al. [12]; Radwan et al. [14] |
| Grain mold | IS# 602, 603, 608, 1233, 2413, 3121, 12697, 12804, 20727, 20740, 20743, 20816, 30562, 31681, 2379, 2864, 12302, 13971, 17941, 19389, 23992, 26694, 29335, 21512, 21645, 12945, 22294, 995, 2426, 12706, 16151, 24453, 26701, 29326, 30383, 30533, 30536, 20956, 29314, 30092, 10969, 23590, 29187, 29269, 473, 29304, 1212, 13893, 29241, 29568 | Sharma et al. [12] |
| Anthracnose | IS# 473, 5301, 6354, 7679, 10302, 16382, 19153, 20632, 20956, 23521, 23684, 24218, 24939 | Sharma et al. [13] |
| Leaf blight | IS# 473, 2382, 7131, 9108, 9177, 9745, 12937, 12945, 14861, 19445, 20743, 21083, 23521, 23644, 23684, 24175, 24503, 24939, 24953, 26694, 26749, 28614, 29187, 29233, 29714, 31557, 33353, | Sharma et al. [13] |
| Charcoal rot | IS# 24463; 4515, 13549, 29582, 25301, 12735, 30533, 23514, 29950, 14010, 14090, 29358, 19859, 16528, 22986, 5094, 26046, 23590, 24503, 21512, 29269, 27697, 19676, 19389, 22294, 7250, 17941, 602, 30092, 29733, 31557, 23216, 10757, 12945, 29606, 12697, 31651, 7679, 23891, 32787, 29091, 29335, 30466, 4631, 29233, 28451, 24218, 1041, 30507, 29627, 2379 | Borphukan [15]; Kapanigowda et al. [17] |
| Rust | IS# 473, 23521, 23684, 24503, 26737, 33023 | Sharma et al. [13] |
| Potyvirus spp. | IS# 7679 and 20740 | Seifers et al. [16] |
| Shoot fly | IS# 2205, 4515, 4698, 5094 | ICRISAT Unpublished |
| Spotted stem borer | IS# 4698, 5094, 1041, 18039, 19445, 23992 | ICRISAT Unpublished |
| Sugarcane Aphid | IS# 2205, 4515, 4698, 18039, 1004, 3121, 4581, 5386, 12937, 15744, 16528, 20625, 20632, 23514, 23521, 23586, 23684, 24492, 24939, 25089, 25249, 25301, 25548, 27034, 27887, 28614, 29314, 29654, 29772, 31446, 31557, 33023 | ICRISAT Unpublished |
| Abiotic stress | ||
| Drought | IS# 14779, 23891, 31714, 4515, 5094, 9108, 15466, 1212 | Upadhyaya et al. [18]; Kapanigowda et al. [17] |
| Seedling vigor under low temperature stress | IS# 1212, 14779, 15170, 22986, 7305, 7310 | Upadhyaya et al. [19] |
| Germinability under low temperature stress | IS# 602, 1233, 7305, 10302, 20956 | Upadhyaya et al. [19] |
| IS# | Origin | Race | Combined Data over Two Seasons and Two Conditions (Optimally Irrigated and Post-Flowering Drought Stress) | ||
|---|---|---|---|---|---|
| Days to 50% Flowering | 100-Seed Weight (g) | Grain Yield (g Plant−1) | |||
| Group 1 | |||||
| 1233 | China | Bicolor | 54 * | 1.64 | 11.74 |
| 2379 | South Africa | Caudatum | 58 * | 2.11 | 13.92 |
| 2864 | South Africa | Caudatum | 59 * | 2.03 | 17.88 |
| 4698 | India | Durra | 77 | 3.18 | 43.32 * |
| 12706 | USA | Caudatum-bicolor | 53 * | 3.33 | 12.08 |
| 14861 | Cameroon | Caudatum | 58 * | 3.94 * | 18.63 |
| 16382 | Cameroon | Guinea | 58 * | 3.24 | 11.12 |
| 17941 | India | Caudatum | 56 * | 2.31 | 15.30 |
| 20298 | Niger | Caudatum-bicolor | 54 * | 2.85 | 12.88 |
| 26484 | Benin | Guinea | 72 | 4.25 * | 24.85 |
| 28313 | Yemen | Durra-caudatum | 52 * | 2.88 | 11.88 |
| 28849 | Yemen | Durra-caudatum | 52 * | 3.17 | 15.41 |
| Control | |||||
| 2205 | India | Durra-bicolor | 80 | 2.03 | 26.16 |
| 18758 | Ethiopia | Guinea-caudatum | 71 | 2.51 | 29.74 |
| 33844 | India | Durra | 78 | 3.44 | 35.25 |
| Trial Mean | 62 | 2.60 | 18.46 | ||
| Trial Range | 52–77 | 1.64–4.25 | 9.08–43.32 | ||
| LSD (P 0.05) | 1.08 | 0.17 | 2.29 | ||
| CV (%) | 2.50 | 9.30 | 19.30 | ||
| Group 2 | |||||
| 23590 | Ethiopia | Guinea-caudatum | 74 | 2.99 | 38.39 * |
| 23891 | Yemen | Durra | 76 | 5.14 * | 40.91 * |
| 31714 | Yemen | Durra-caudatum | 67 * | 4.45 * | 30.80 |
| Control | |||||
| 2205 | India | Durra-bicolor | 80 | 2.11 | 24.39 |
| 18758 | Ethiopia | Guinea-caudatum | 69 | 2.42 | 27.26 |
| 33844 | India | Durra | 77 | 3.61 | 33.49 |
| Trial mean | 68 | 2.41 | 21.02 | ||
| Trial range | 53–86 | 1.27–5.14 | 10.29–40.91 | ||
| LSD (P 0.05) | 1.21 | 0.17 | 2.78 | ||
| CV (%) | 2.30 | 8.80 | 17.10 | ||
| Group 3 | |||||
| 11473 | Ethiopia | Caudatum | 67 * | 4.49 * | 16.17 |
| 15466 | Cameroon | Caudatum | 80 | 3.78 * | 37.72 |
| 15744 | Cameroon | Durra-caudatum | 74 | 3.94 * | 36.96 |
| 28141 | Yemen | Durra-caudatum | 76 | 5.15 * | 40.20 * |
| 30838 | Cameroon | Guinea | 71 | 3.72 * | 23.01 |
| 31706 | Yemen | Durra | 76 | 4.69 * | 35.93 |
| Control | |||||
| 2205 | India | Durra-bicolor | 80 | 2.11 | 26.32 |
| 18758 | Ethiopia | Guinea-caudatum | 70 | 2.32 | 26.74 |
| 33844 | India | Durra | 77 | 3.53 | 35.99 |
| Trial mean | 76 | 2.40 | 23.20 | ||
| Trial range | 67–95 | 0.42–5.15 | 7.23–40.20 | ||
| LSD (P 0.05) | 1.25 | 0.17 | 3.51 | ||
| CV (%) | 2.10 | 8.20 | 18.00 | ||
| IS# | Origin | Race | Traits | Combined Data over Two Seasons and Two Conditions (Optimally Irrigated and Post-Flowering Drought) | |||||
|---|---|---|---|---|---|---|---|---|---|
| Days to 50% Flowering | 100-Seed Weight (g) | Grain Yield (g Plant−1) | Fe (mg kg−1) | Zn (mg kg−1) | Brix (%) | ||||
| 602 | USA | Bicolor | High Fe and Zn, resistant to grain mold and charcoal rot | 67 | 1.27 | 13.74 | 42.79 | 35.98 | 9.19 |
| 1004 | India | Durra | High Brix % and resistant to aphids | 86 | 3.78 | 38.71 | 28.51 | 27.38 | 12.28 |
| 1212 | China | Kafir-bicolor | High Fe and Zn, tolerance to drought and low temperature stress (high seedling vigor), and resistant to downy mildew and grain mold | 63 | 2.26 | 16.14 | 43.57 | 38.21 | 10.06 |
| 1233 | China | Bicolor | Early flowering and high Fe and Zn contents | 54 | 1.64 | 11.74 | 48.46 | 39.05 | 8.98 |
| 4515 | India | Durra | Tolerant to drought, and resistant to charcoal rot, shoot fly and aphids | 75 | 2.99 | 34.28 | 28.36 | 20.47 | 12.43 |
| 4698 | India | Durra | High Brix %, high grain yielding and resistant to stem borer, shoot fly and aphids | 77 | 3.18 | 43.32 | 23.91 | 20.35 | 12.75 |
| 4951 | India | Guinea | High Fe, Zn, protein contents and high saccharification yield | 83 | 1.64 | 14.19 | 41.90 | 33.11 | 12.08 |
| 5094 | India | Durra | Tolerant to drought, and resistant to downy mildew, charcoal rot, stem borer and shoot fly | 77 | 2.46 | 31.27 | 32.57 | 22.39 | 14.23 |
| 7305 | Nigeria | Caudatum | High saccharification yield and tolerant to low temperature stress (high germinability and vigor) | 74 | 3.04 | 24.37 | 37.25 | 25.63 | 10.73 |
| 13549 | Mexico | Caudatum-bicolor | High Brix %, and resistant to downy mildew and charcoal rot | 87 | 1.75 | 15.91 | 30.04 | 29.15 | 13.99 |
| 14861 | Cameroon | Caudatum | Early flowering, greater seed weight, and resistant to leaf blight | 58 | 3.94 | 18.63 | 33.13 | 20.28 | 9.22 |
| 15466 | Cameroon | Caudatum | Greater seed weight, high grain yielding and tolerant to drought | 80 | 3.78 | 37.72 | 28.07 | 13.53 | 13.23 |
| 15744 | Cameroon | Durra-caudatum | Greater seed weight and resistant to aphids | 74 | 3.94 | 36.96 | 27.23 | 15.40 | 11.61 |
| 16382 | Cameroon | Guinea | Early flowering and high Fe content | 58 | 3.24 | 11.12 | 46.55 | 23.66 | 9.19 |
| 17941 | India | Caudatum | Early flowering and resistant to grain mold and charcoal rot | 56 | 2.31 | 15.30 | 29.83 | 19.73 | 8.38 |
| 20743 | USA | Bicolor | High Fe and Zn, and resistant to grain mold and leaf blight | 63 | 1.80 | 15.18 | 46.18 | 29.74 | 8.19 |
| 20956 | Indonesia | Durra-caudatum | Tolerant to low temperature stress (high germinability) and resistant to grain mold and anthracnose | 72 | 2.97 | 24.38 | 35.06 | 25.72 | 11.71 |
| 23216 | Zambia | Caudatum-bicolor | High Brix % and resistant to downy mildew and charcoal rot | 86 | 1.89 | 12.28 | 33.36 | 24.49 | 14.69 |
| 23590 | Ethiopia | Guinea-caudatum | High grain yielding and resistant to grain mold and charcoal rot | 74 | 2.99 | 38.39 | 30.38 | 20.06 | 12.01 |
| 23684 | Mozambique | Guinea | High protein and Brix % and resistant to anthracnose, leaf blight, rust and aphids | 97 | 1.60 | 6.82 | 39.14 | 29.55 | 14.53 |
| 23891 | Yemen | Durra | Greater seed weight, high yielding, high Brix %, tolerant to drought and resistant to charcoal rot | 76 | 5.14 | 40.91 | 31.59 | 27.23 | 11.72 |
| 24139 | Tanzania | Guinea | High Zn and Brix % | 95 | 1.65 | 13.75 | 41.84 | 35.32 | 15.16 |
| 24939 | Zambia | Bicolor | High Brix %, and resistant to anthracnose, leaf blight and aphids | 85 | 1.76 | 12.18 | 42.13 | 29.65 | 14.61 |
| 26484 | Benin | Guinea | Greater seed weight and resistant to downy mildew | 72 | 4.25 | 24.85 | 41.00 | 27.98 | 10.73 |
| 28141 | Yemen | Durra-caudatum | High grain yielding, greater seed weight and high Brix % | 76 | 5.15 | 40.20 | 34.41 | 26.98 | 11.80 |
| 28313 | Yemen | Durra-caudatum | Early flowering and high Fe content | 52 | 2.88 | 11.88 | 47.93 | 29.16 | 10.15 |
| 30383 | China | Caudatum-bicolor | High Fe and Zn contents, and resistant to downy mildew and grain mold | 65 | 1.99 | 15.88 | 45.05 | 38.56 | 9.77 |
| 31714 | Yemen | Durra-caudatum | Greater seed weight, tolerant to drought and resistant to downy mildew | 67 | 4.45 | 30.80 | 32.99 | 21.36 | 10.93 |
| Pair of Accessions | Genetic Distance | Pair of Accessions | Genetic Distance |
|---|---|---|---|
| Agronomically desirable sources | |||
| IS 28141 and IS 16382 | 0.455 | IS 30838 and IS 28141 | 0.413 |
| IS 16382 and IS 4698 | 0.452 | IS 31714 and IS 2379 | 0.411 |
| IS 23891 and IS 16382 | 0.443 | IS 28849 and IS 16382 | 0.410 |
| IS 26484 and IS 4698 | 0.438 | IS 31714 and IS 30838 | 0.410 |
| IS 31714 and IS 16382 | 0.432 | IS 26484 and IS 23891 | 0.407 |
| IS 31706 and IS 16382 | 0.428 | IS 23891 and IS 2379 | 0.406 |
| IS 28141 and IS 26484 | 0.424 | IS 28141 and IS 23590 | 0.406 |
| IS 30838 and IS 4698 | 0.419 | IS 31714 and IS 26484 | 0.406 |
| IS 28141 and IS 2379 | 0.415 | IS 4698 and IS 2864 | 0.403 |
| IS 23590 and IS 4698 | 0.413 | IS 4698 and IS 2379 | 0.402 |
| Mean distance of 0.311 among 276 pairs of accessions; Minimum 0.038 between IS 17941 and IS 2864; Maximum 0.455 between IS 28141 and IS 16362 | |||
| Grain nutrient rich sources | |||
| IS 30460 and IS 25989 | 0.526 | IS 25989 and IS 25836 | 0.461 |
| IS 25989 and IS 5386 | 0.521 | IS 23684 and IS 1212 | 0.460 |
| IS 25989 and IS 3971 | 0.516 | IS 25989 and IS 8774 | 0.458 |
| IS 25989 and IS 1212 | 0.515 | IS 24139 and IS 1212 | 0.456 |
| IS 30383 and IS 25989 | 0.508 | IS 30460 and IS 24139 | 0.454 |
| IS 25989 and IS 1219 | 0.505 | IS 25989 and IS 602 | 0.453 |
| IS 30536 and IS 25989 | 0.502 | IS 23684 and IS 1219 | 0.452 |
| IS 27786 and IS 25989 | 0.491 | IS 30536 and IS 23684 | 0.451 |
| IS 31681 and IS 25989 | 0.479 | IS 25989 and IS 23684 | 0.448 |
| IS 30460 and IS 23684 | 0.465 | IS 30536 and IS 24139 | 0.448 |
| Mean distance of 0.329 among 351 pairs of accessions; Minimum 0.020 between IS 26025 and IS 19975; Maximum 0.526 between IS 30460 and IS 25989 | |||
| Bioenergy traits | |||
| IS 27887 and IS 4698 | 0.457 | IS 28141 and IS 23216 | 0.427 |
| IS 28141 and IS 24139 | 0.456 | IS 28141 and IS 24939 | 0.427 |
| IS 24139 and IS 4698 | 0.448 | IS 27887 and IS 13549 | 0.421 |
| IS 24139 and IS 23891 | 0.443 | IS 24939 and IS 4698 | 0.420 |
| IS 27887 and IS 28141 | 0.435 | IS 24939 and IS 23891 | 0.419 |
| IS 33353 and IS 4698 | 0.434 | IS 24953 and IS 23891 | 0.418 |
| IS 23216 and IS 4698 | 0.433 | IS 33353 and IS 23891 | 0.418 |
| IS 28141 and IS 24953 | 0.431 | IS 28141 and IS 7305 | 0.416 |
| IS 27887 and IS 23891 | 0.431 | IS 24953 and IS 4698 | 0.407 |
| IS 33353 and IS 28141 | 0.430 | IS 13549 and IS 4951 | 0.407 |
| Mean distance of 0.338 among 78 pairs of accessions; Minimum 0.104 between IS 28141 and IS 23891; Maximum 0.457 between IS 27887 and IS 4698 | |||
| Biotic stress resistance sources | |||
| IS 30466 and IS 21512 | 0.499 | IS 30562 and IS 23684 | 0.479 |
| IS 29654 and IS 21512 | 0.498 | IS 23684 and IS 5094 | 0.478 |
| IS 30562 and IS 21512 | 0.497 | IS 30466 and IS 23684 | 0.475 |
| IS 30466 and IS 24218 | 0.488 | IS 29654 and IS 24218 | 0.475 |
| IS 21512 and IS 1212 | 0.486 | IS 30533 and IS 24218 | 0.475 |
| IS 21512 and IS 5094 | 0.486 | IS 31714 and IS 21512 | 0.474 |
| IS 30533 and IS 21512 | 0.485 | IS 33023 and IS 29654 | 0.474 |
| IS 30562 and IS 24218 | 0.485 | IS 29091 and IS 24218 | 0.473 |
| IS 33023 and IS 30466 | 0.482 | IS 24218 and IS 1212 | 0.472 |
| IS 24218 and IS 5094 | 0.479 | IS 21512 and IS 4631 | 0.472 |
| IS 30383 and IS 21512 | 0.479 | ||
| Mean distance of 0.332 among 2415 pairs of accessions; Minimum 0.031 between IS 29241 and IS 29187; Maximum 0.499 between IS 30466 and IS 21512 | |||
| Abiotic stress tolerance sources | |||
| IS 9108 and IS 5094 | 0.427 | IS 14779 and IS 31714 | 0.404 |
| IS 9108 and IS 1212 | 0.419 | IS 7305 and IS 4515 | 0.401 |
| IS 9108 and IS 31714 | 0.417 | IS 14779 and IS 5094 | 0.398 |
| IS 7305 and IS 5094 | 0.416 | IS 9108 and IS 23891 | 0.397 |
| IS 7305 and IS 31714 | 0.415 | IS 14779 and IS 23891 | 0.397 |
| IS 10302 and IS 5094 | 0.407 | IS 10302 and IS 23891 | 0.392 |
| IS 7305 and IS 1212 | 0.405 | IS 10302 and IS 4515 | 0.391 |
| IS 14779 and IS 1212 | 0.405 | IS 7305 and IS 23891 | 0.39 |
| IS 9108 and IS 4515 | 0.405 | IS 10302 and IS 31714 | 0.389 |
| IS 10302 and IS 1212 | 0.404 | IS 15466 and IS 1212 | 0.387 |
| Mean distance of 0.301 among 66 pairs of accessions; Minimum 0.008 between IS 20956 and IS 10302; Maximum 0.427 between IS 9108 and IS 5094 | |||
| Multi-trait-specific sources | |||
| IS 23684 and IS 5094 | 0.478 | IS 16382 and IS 4698 | 0.452 |
| IS 28141 and IS 23684 | 0.465 | IS 31714 and IS 23684 | 0.452 |
| IS 24139 and IS 5094 | 0.462 | IS 23891 and IS 23684 | 0.452 |
| IS 23684 and IS 1212 | 0.460 | IS 23684 and IS 4515 | 0.451 |
| IS 23684 and IS 4698 | 0.458 | IS 24139 and IS 4698 | 0.448 |
| IS 31714 and IS 24139 | 0.457 | IS 30383 and IS 23684 | 0.446 |
| IS 24139 and IS 1212 | 0.456 | IS 16382 and IS 4515 | 0.444 |
| IS 16382 and IS 5094 | 0.456 | IS 24139 and IS 4515 | 0.444 |
| IS 28141 and IS 24139 | 0.456 | IS 23891 and IS 16382 | 0.443 |
| IS 28141 and IS 16382 | 0.455 | IS 24139 and IS 23891 | 0.443 |
| Mean distance of 0.328 among 378 pairs of accessions; Minimum 0.043 between IS 4698 and IS 4515; Maximum 0.478 between IS 23684 and IS 5094 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Upadhyaya, H.D.; Vetriventhan, M.; Asiri, A.M.; C.R. Azevedo, V.; Sharma, H.C.; Sharma, R.; Sharma, S.P.; Wang, Y.-H. Multi-Trait Diverse Germplasm Sources from Mini Core Collection for Sorghum Improvement. Agriculture 2019, 9, 121. https://doi.org/10.3390/agriculture9060121
Upadhyaya HD, Vetriventhan M, Asiri AM, C.R. Azevedo V, Sharma HC, Sharma R, Sharma SP, Wang Y-H. Multi-Trait Diverse Germplasm Sources from Mini Core Collection for Sorghum Improvement. Agriculture. 2019; 9(6):121. https://doi.org/10.3390/agriculture9060121
Chicago/Turabian StyleUpadhyaya, Hari D, Mani Vetriventhan, Abdullah M. Asiri, Vania C.R. Azevedo, Hari C. Sharma, Rajan Sharma, Suraj Prasad Sharma, and Yi-Hong Wang. 2019. "Multi-Trait Diverse Germplasm Sources from Mini Core Collection for Sorghum Improvement" Agriculture 9, no. 6: 121. https://doi.org/10.3390/agriculture9060121
APA StyleUpadhyaya, H. D., Vetriventhan, M., Asiri, A. M., C.R. Azevedo, V., Sharma, H. C., Sharma, R., Sharma, S. P., & Wang, Y. -H. (2019). Multi-Trait Diverse Germplasm Sources from Mini Core Collection for Sorghum Improvement. Agriculture, 9(6), 121. https://doi.org/10.3390/agriculture9060121