
Feeding Strategy of the Wild Korean Seahorse (Hippocampus haema)

 ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
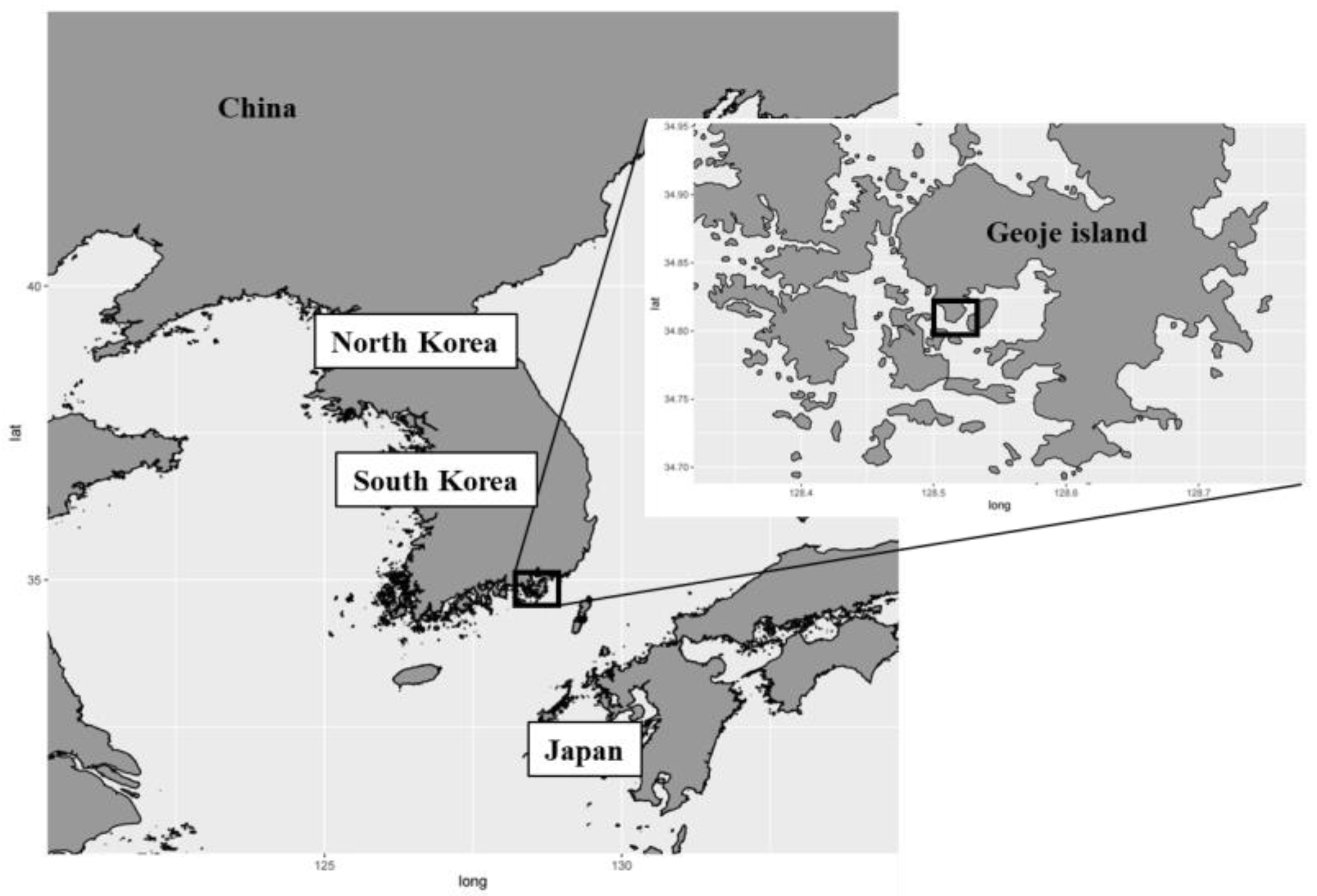
2.1. Study Area and Samplings
2.2. Genomic DNA Extraction and NGS Library Construction
2.3. Bioinformatic Analysis
3. Results
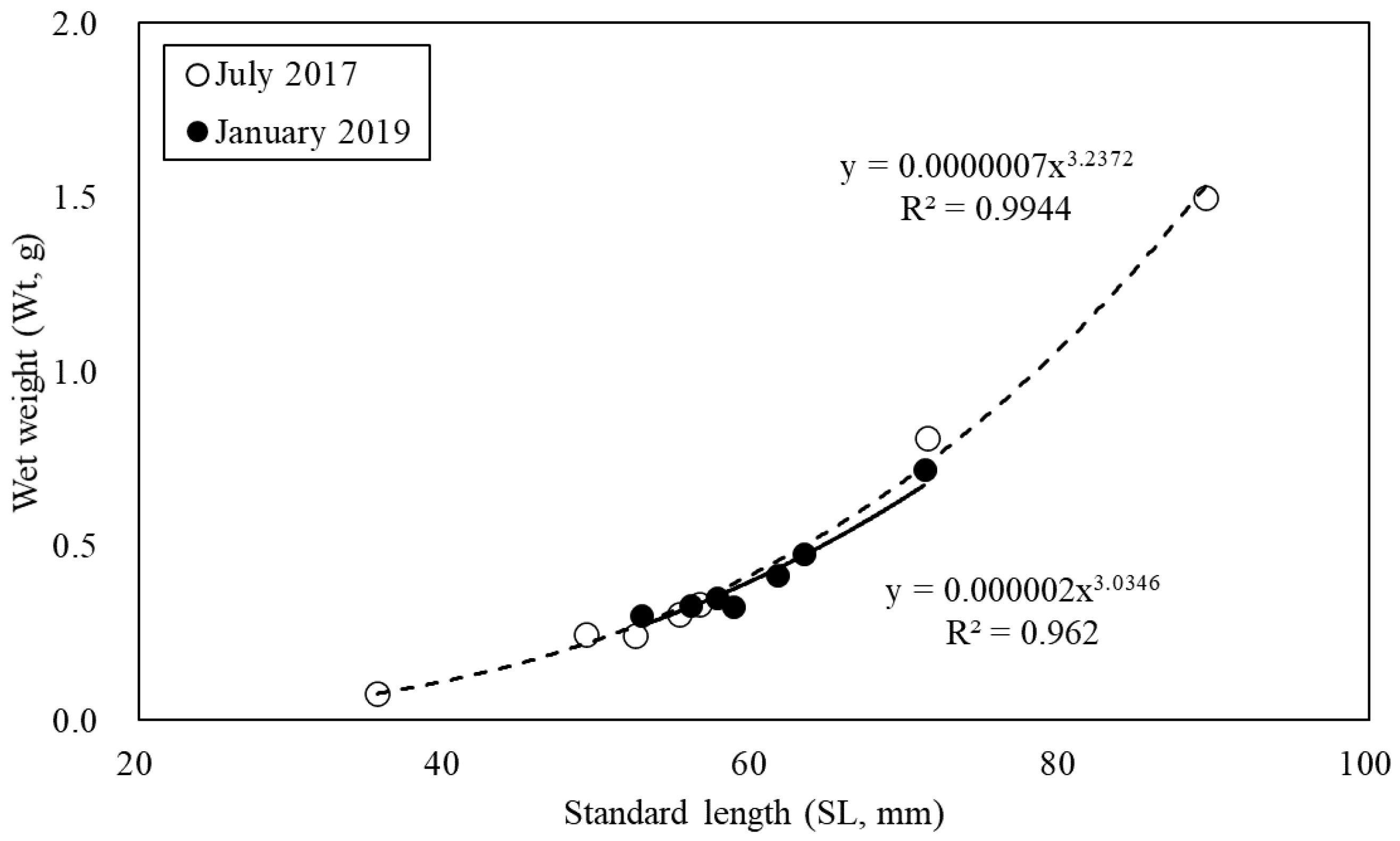
3.1. Morphometric Measurements
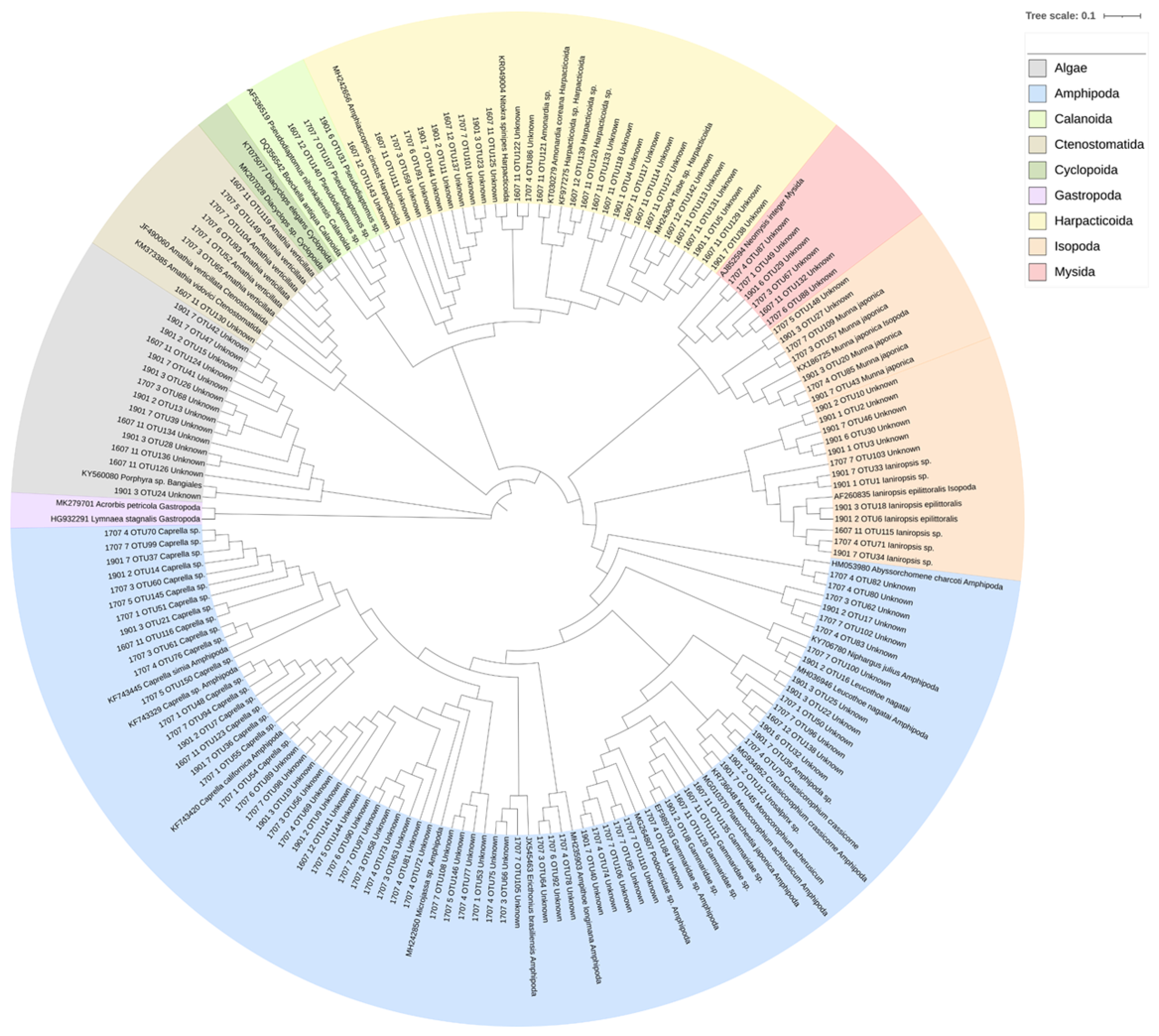
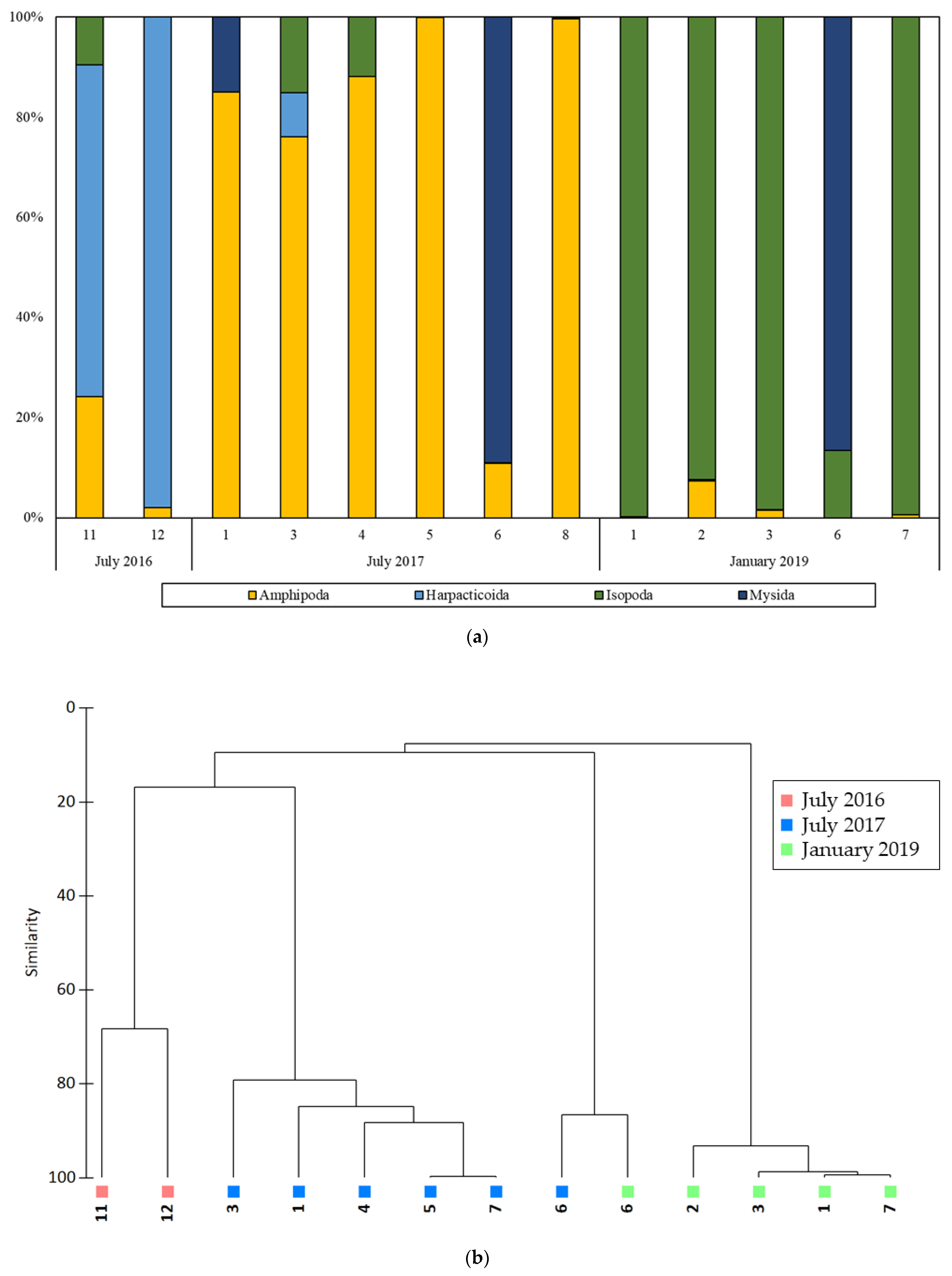
3.2. NGS Analysis
4. Discussion
NGS Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gasparini, J.L.; Floeter, S.R.; Ferreira, C.E.L.; Sazima, I. Marine ornamental trade in Brazil. Biodivers. Conserv. 2005, 14, 2883–2899. [Google Scholar] [CrossRef]
- Vincent, A.C.J.; Foster, S.J.; Koldewey, H.J. Conservation and management of seahorses and other Syngnathidae. J. Fish Biol. 2011, 78, 1681–1724. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.J.; Vincent, A.C.J. Life history and ecology of seahorses: Implications for conservation and management. J. Fish Biol. 2004, 65, 1–61. [Google Scholar] [CrossRef]
- Martin-Smith, K.M.; Samoilys, M.A.; Meeuwig, J.J.; Vincent, A.C.J. Collaborative development of management options for an artisanal fishery for seahorses in the central Philippines. Ocean Coast. Manag. 2004, 47, 165–193. [Google Scholar] [CrossRef]
- Curtis, J.M.R.; Ribeiro, J.; Erzini, K.; Vincent, A.C.J. A conservation trade-off? Interspecific differences in seahorse responses to experimental changes in fishing effort. Aquat. Conserv. Mar. Freshw. Ecosyst. 2007, 17, 468–484. [Google Scholar] [CrossRef]
- Lourie, S.A.; Pritchard, J.C.; Casey, S.P.; Truong, S.K.; Hall, H.J.; Vincent, A.C.J. The taxonomy of Vietnam’s exploited seahorses (family Syngnathidae). Biol. J. Linn. Soc. 1999, 66, 231–256. [Google Scholar] [CrossRef]
- Caldwell, I.R.; Vincent, A.C.J. A sedentary fish on the move: Effects of displacement on long-snouted seahorse (Hippocampus guttulatus Cuvier) movement and habitat use. Environ. Biol. Fishes 2012, 96, 67–75. [Google Scholar] [CrossRef]
- Kim, M.J.; Kim, H.C.; Lee, W.C.; Park, J.M.; Kwak, S.N.; Oh, Y.; Kang, M.G.; Lee, S.H. Ecological Characteristics of the New Recorded Seahorse (Hippocampus haema) in Geoje-Hansan Bay, Korea. J. Coast. Res. 2018, 85, 351–355. [Google Scholar] [CrossRef]
- Park, J.M.; Kwak, S.N. Length–weight relationships and reproductive characteristics of the crowned seahorse (Hippocampus coronatus) in eelgrass beds (Zostera marina) of Dongdae Bay, Korea. In Marine Biology Research; Taylor and Francis Ltd.: Abingdon, UK, 2014; Volume 11. [Google Scholar] [CrossRef]
- Bell, E.M.; Lockyear, J.F.; Mcpherson, J.M.; Marsden, A.D.; Vincent, A.C.J. First field studies of an Endangered South African seahorse, Hippocampus capensis. Environ. Biol. Fishes 2003, 67, 35–46. [Google Scholar] [CrossRef]
- Caldwell, I.R.; Vincent, A.C.J. Revisiting two sympatric European seahorse species: Apparent decline in the absence of exploitation. Aquat. Conserv. Mar. Freshw. Ecosyst. 2012, 22, 427–435. [Google Scholar] [CrossRef]
- Vincent, A.C.J.; Evans, K.L.; Marsden, A.D. Home range behaviour of the monogamous Australian seahorse, Hippocampus whitei. Environ. Biol. Fishes 2005, 72, 1–12. [Google Scholar] [CrossRef]
- Vincent, A.C.J.; Sadler, L.M. Faithful pair bonds in wild seahorses, Hippocampus whitei. Anim. Behav. 1995, 50, 1557–1569. [Google Scholar] [CrossRef]
- Choi, Y.-U.; Rho, S.; Jung, M.-M.; Lee, Y.-D.; Noh, G.-A. Parturition and Early Growth of Crowned Seahorse, Hippocampus coronatus in Korea. J. Aquac. 2006, 19, 109–118. [Google Scholar]
- Jung, M.-M.; Choi, Y.-U.; Lee, J.-E.; Kim, J.-W.; Kim, S.-C.; Lee, Y.-H.; Rho, S. Coexisting Fish Fauna in the Seahorse Habitats. J. Aquac. 2007, 20, 41–46. [Google Scholar]
- Kim, I.-S.; Lee, W.-O. First Record of the Seahorse Fish, Hippocampus trimaculatus (Pisces: Syngnathidae) from Korea. Korean J. Zool. 1995, 38, 74–77. [Google Scholar]
- Kim, S.-Y.; Kweon, S.-M.; Choi, S.-H. First Record of Hippocampus sindonis (Syngnathiformes: Syngnathidae) from Korea. Korean J. Ichthyol. 2013, 25, 42–45. [Google Scholar]
- Han, S.-Y.; Kim, J.-K.; Kai, Y.; Senou, H. Seahorses of the Hippocampus coronatus complex: Taxonomic revision, and description of Hippocampus haema, a new species from Korea and Japan (Teleostei, Syngnathidae). ZooKeys 2017, 712, 113–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braga, R.R.; Bornatowski, H.; Vitule, J.R.S. Feeding ecology of fishes: An overview of worldwide publications. Rev. Fish Biol. Fish. 2012, 22, 915–929. [Google Scholar] [CrossRef]
- Huh, S.-H.; Kwak, S.N. Feeding habits of Favonigobius gymnauchen in the eelgrass (Zostera marina) bed in Kwangyang Bay. J. Korean Fish. Soc. 1998, 31, 372–379. [Google Scholar]
- Rosas-Luis, R.; Navarro, J.; Loor-Andrade, P.; Forero, M.G. Feeding ecology and trophic relationships of pelagic sharks and billfishes coexisting in the central eastern Pacific Ocean. Mar. Ecol. Prog. Ser. 2017, 573, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Shreeve, R.S.; Collins, M.A.; Tarling, G.A.; Main, C.E.; Ward, P.; Johnston, N.M. Feeding ecology of myctophid fishes in the northern Scotia Sea. Mar. Ecol. Prog. Ser. 2009, 386, 221–236. [Google Scholar] [CrossRef] [Green Version]
- Tipton, K.; Bell, S.S. Foraging patterns of two syngnathid fishes: Importance of harpacticoid copepods. Mar. Ecol. Prog. Ser. 1988, 47, 31–43. [Google Scholar] [CrossRef]
- Woods, C.M.C. Natural diet of the seahorse Hippocampus abdominalis. N. Z. J. Mar. Freshw. Res. 2002, 36, 655–660. [Google Scholar] [CrossRef]
- Woods, C.M.C. Growth and survival of juvenile seahorse Hippocampus abdominalis reared on live, frozen and artificial foods. Aquaculture 2003, 220, 287–298. [Google Scholar] [CrossRef]
- Garcia, L.M.B.; Hilomen-Garcia, G.V.; Celino, F.T.; Gonzales, T.T.; Maliao, R.J. Diet composition and feeding periodicity of the seahorse Hippocampus barbouri reared in illuminated sea cages. Aquaculture 2012, 358–359, 1–5. [Google Scholar] [CrossRef]
- Vargas-Abúndez, J.A.; Simões, N.; Mascaró, M. Feeding the lined seahorse Hippocampus erectus with frozen amphipods. Aquaculture 2018, 491, 82–85. [Google Scholar] [CrossRef]
- Palma, J.; Stockdale, J.; Correia, M.; Andrade, J.P. Growth and survival of adult long snout seahorse (Hippocampus guttulatus) using frozen diets. Aquaculture 2008, 278, 55–59. [Google Scholar] [CrossRef]
- Segade, A.; Robaina, L.; Otero-Ferrer, F.; Romero, J.G.; Dominguez, L.M. Effects of the diet on seahorse (Hippocampus hippocampus) growth, body colour and biochemical composition. Aquac. Nutr. 2015, 21, 807–813. [Google Scholar] [CrossRef]
- Celino, F.T.; Hilomen-Garcia, G.V.; del Norte-Campos, A.G.C. Feeding selectivity of the seahorse, Hippocampus kuda (Bleeker), juveniles under laboratory conditions. Aquac. Res. 2012, 43, 1804–1815. [Google Scholar] [CrossRef]
- Felício, A.K.C.; Rosa, I.L.; Souto, A.; Freitas, R.H.A. Feeding behavior of the longsnout seahorse Hippocampus reidi Ginsburg, 1933. J. Ethol. 2006, 24, 219–225. [Google Scholar] [CrossRef]
- Olivotto, I.; Avella, M.A.; Sampaolesi, G.; Piccinetti, C.C.; Navarro Ruiz, P.; Carnevali, O. Breeding and rearing the longsnout seahorse Hippocampus reidi: Rearing and feeding studies. Aquaculture 2008, 283, 92–96. [Google Scholar] [CrossRef]
- Balasubramanian, R. Food and feeding habits of seahorse, Hippocampus kelloggi (Jordan and Snyder, 1902) in Cuddalore coastal water, Southeast coast of India. Indian J. Educ. Inf. Manag. 2017, 6, 1–8. [Google Scholar]
- da Costa Castro, A.L.; de Farias Diniz, A.; Martins, I.Z.; Vendel, A.L.; de Oliveira, T.P.R.; de Lucena Rosa, I.M. Assessing diet composition of seahorses in the wild using a non- destructive method: Hippocampus reidi (Teleostei: Syngnathidae) as a study-case. Neotrop. Ichthyol. 2008, 6, 637–644. [Google Scholar] [CrossRef]
- Gurkan, S.; Taskavak, E.; Murat Sever, T.; Akalin, S. Gut Contents of Two European Seahorses Hippocampus hippocampus and Hippocampus guttulatus in the Aegean Sea, Coasts of Turkey. Pak. J. Zool. 2011, 43, 1197–1201. [Google Scholar]
- Huh, S.-H.; Park, J.M.; Kwak, S.N.; Seong, B.J. Abundances and feeding habits of Hippocampus coronatus in an eelgrass (Zostera marina) bed of Dongdae Bay, Korea. J. Korean Soc. Fish. Technol. 2014, 50, 115–123. [Google Scholar] [CrossRef]
- Storero, L.P.; Gonzélez, R.A. Feeding habits of the seahorse Hippocampus patagonicus in San Antonio Bay (Patagonia, Argentina). J. Mar. Biol. Assoc. UK 2008, 88, 1503–1508. [Google Scholar] [CrossRef]
- Yip, M.Y.; Lim, A.C.O.; Chong, V.C.; Lawson, J.M.; Foster, S.J. Food and feeding habits of the seahorses Hippocampus spinosissimus and Hippocampus trimaculatus (Malaysia). J. Mar. Biol. Assoc. UK 2015, 95, 1033–1040. [Google Scholar] [CrossRef] [Green Version]
- Valladares, S.; Soto, D.X.; Planas, M. Dietary composition of endangered seahorses determined by stable isotope analysis. Mar. Freshw. Res. 2017, 68, 831–839. [Google Scholar] [CrossRef] [Green Version]
- Ape, F.; Corriero, G.; Mirto, S.; Pierri, C.; Lazic, T.; Gristina, M. Trophic flexibility and prey selection of the wild long-snouted seahorse Hippocampus guttulatus Cuvier, 1829 in three coastal habitats. Estuar. Coast. Shelf Sci. 2019, 224, 1–10. [Google Scholar] [CrossRef]
- Manning, C.G.; Foster, S.J.; Vincent, A.C.J. A review of the diets and feeding behaviours of a family of biologically diverse marine fishes (Family Syngnathidae). In Reviews in Fish Biology and Fisheries; Springer International Publishing: Cham, Switzerland, 2019; pp. 1–25. [Google Scholar] [CrossRef]
- Lazic, T.; Corriero, G.; Balech, B.; Cardone, F.; Deflorio, M.; Fosso, B.; Gissi, C.; Marzano, M.; Pesole, G.; Santamaria, M.; et al. Evaluating DNA metabarcoding to analyze diet composition of wild long-snouted seahorse Hippocampus guttulatus. In 2021 International Workshop on Metrology for the Sea; Learning to Measure Sea Health Parameters (MetroSea); IEEE: Reggio Calabria, Italy, 2021; pp. 257–261. [Google Scholar] [CrossRef]
- Young, T.; Pincin, J.; Neubauer, P.; Ortega-García, S.; Jensen, O.P. Investigating diet patterns of highly mobile marine predators using stomach contents, stable isotope, and fatty acid analyses. ICES J. Mar. Sci. 2018, 75, 1583–1590. [Google Scholar] [CrossRef]
- Zorica, B.; Ezgeta-Balić, D.; Vidjak, O.; Vuletin, V.; Šestanović, M.; Isajlović, I.; Čikeš Keč, V.; Vrgoč, N.; Harrod, C. Diet Composition and Isotopic Analysis of Nine Important Fisheries Resources in the Eastern Adriatic Sea (Mediterranean). Front. Mar. Sci. 2021, 8, 183. [Google Scholar] [CrossRef]
- Piñol, J.; San Andrés, V.; Clare, E.L.; Mir, G.; Symondson, W.O.C. A pragmatic approach to the analysis of diets of generalist predators: The use of next-generation sequencing with no blocking probes. Mol. Ecol. Resour. 2014, 14, 18–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lourie, S.A.; Foster, S.J.; Cooper, E.W.T.; Vincent, A.C.J. A guide to the identification of seahorses. In Project Seahorse and TRAFFIC North America; Project Seahorse and TRAFFIC North America: Vancouver, Canada, 2004. [Google Scholar]
- Yoon, T.H.; Kang, H.E.; Lee, S.R.; Lee, J.B.; Baeck, G.W.; Park, H.; Kim, H.W. Metabarcoding analysis of the stomach contents of the Antarctic Toothfish (Dissostichus mawsoni) collected in the Antarctic Ocean. PeerJ 2017, 5, e3977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vestheim, H.; Jarman, S.N. Blocking primers to enhance PCR amplification of rare sequences in mixed samples–a case study on prey DNA in Antarctic krill stomachs. Front. Zool. 2008, 5, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–1000. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Planas, M.; Blanco, A.; Chamorro, A.; Valladares, S.; Pintado, J. Temperature-induced changes of growth and survival in the early development of the seahorse Hippocampus guttulatus. J. Exp. Mar. Biol. Ecol. 2012, 438, 154–162. [Google Scholar] [CrossRef]
- Strawn, K. Life History of the Pigmy Seahorse, Hippocampus zosterae Jordan and Gilbert, at Cedar Key, Florida. Copeia 1958, 1958, 16–22. Available online: https://about.jstor.org/terms (accessed on 7 February 2022). [CrossRef]
- Kendrick, A.J.; Hyndes, G.A. Variations in the dietary compositions of morphologically diverse syngnathid fishes. Environ. Biol. Fishes 2005, 72, 415–427. [Google Scholar] [CrossRef]
- Reid, G.K. An ecological study of the Gulf of the Mexico fishes, in the vicinity of Cedar Key, Florida. Bull. Mar. Sci. Gulf Caribb. 1954, 4, 1–12. [Google Scholar]
- Teixeira, R.L.; Musick, J.A. Reproduction and food habits of the lined seahorse, Hippocampus erectus (Teleostei: Syngnathidae) of Chesapeake Bay, Virginia. Rev. Brasil. Biol. 2001, 61, 79–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caine, E.A. Ecology of Two Littoral Species of Caprellid Amphipods (Crustacea) from Washington, USA. Mar. Biol. 1980, 56, 327–335. [Google Scholar] [CrossRef]
- Lolas, A.; Vafidis, D. Population dynamics of two caprellid species (Crustaceae: Amphipoda: Caprellidae) from shallow hard bottom assemblages. Mar. Biodivers. 2013, 43, 227–236. [Google Scholar] [CrossRef]
- Yun, S.G.; Byun, S.H.; Kwak, S.N.; Huh, S.-H. Seasonal Variation of Caprellids (Crustacea: Amphipoda) on Blades of Zostera marina in Kwangyang Bay, Korea. J. Korean Fish. 2002, 35, 105–109. [Google Scholar]
- Edgar, G.J. Population regulation, population dynamics and competition amongst mobile epifauna associated with seagrass. Mar. Biol. Ecol. 1990, 144, 205–234. [Google Scholar] [CrossRef]
- Bae, J.-Y.; Noh, G.E.; Park, W.-G. Population structure and life history of Neomysis nigra Nakazawa, 1910 (Mysida) on Jeju Island, South Korea. Crustaceana 2016, 89, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Ohtsuka, S.; Inagaki, H.; Onbe, T.; Gushima, K.; Yoon, Y.H. Direct observations of groups of mysids in shallow coastal waters of western Japan and southern Korea. Mar. Ecol. Prog. Ser. 1995, 123, 33–44. [Google Scholar] [CrossRef] [Green Version]
- Ryer, C.H.; Orth, R.J. Feeding Ecology of the Northern Pipefish, Syngnathus fuscus, in a Seagrass Community of the Lower Chesapeake Bay. Estuaries 1987, 10, 330–336. [Google Scholar] [CrossRef]
- Shin, A.; Kim, D.; Kang, T.; Oh, J.H. Seasonal fluctuation of the meiobenthic fauna community in the intertidal zone sediments of coastal areas in Jeju Island, Korea. Environ. Biol. Res. 2019, 37, 406–425. [Google Scholar] [CrossRef]
- Song, S.J.; Ryu, J.; Khim, J.S.; Kim, W.; Yun, S.G. Seasonal variability of community structure and breeding activity in marine phytal harpacticoid copepods on Ulva pertusa from Pohang, east coast of Korea. J. Sea Res. 2010, 63, 1–10. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Date | July 2017 | January 2019 | July 2016 | |
|---|---|---|---|---|
| n | 7 | 7 | 2 | |
| Range | SL | 35.5–89.4 | 52.7–71.1 | 71.4–83.6 |
| Wt | 0.078–1.500 | 0.300–0.721 | 0.681–0.814 | |
| Average | SL | 58.43 | 60.08 | 77.50 |
| Wt | 0.50 | 0.42 | 0.75 | |
| SD | SL | 17.27 | 5.96 | 8.57 |
| Wt | 0.49 | 0.15 | 0.09 | |
| t | SL | 0.815 | ||
| Wt | 0.679 | |||
| Name | Direction | Sequence (5′ to 3′) | Reference |
|---|---|---|---|
| COIMISQ | Forward | ATNGGNGGNTTYGGNAA | [47] |
| Reverse | TANACYTCNGGRTGNCC | ||
| NEXCOIMISQ | Forward | TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGGGNGGNTTYGGNAAYTG | |
| Reverse | GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGGGRTGNCCRAARAAYCA | ||
| Blocking Primer | GCTTTGGTAATTGACTTG-C3 spacer | This study |
| Phylum | Class | Order | Species | Proportion (%) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| July 2016 | July 2017 | January 2019 | ||||||||||||||
| No. 11 | No. 12 | No. 1 | No. 3 | No. 4 | No. 5 | No. 6 | No. 7 | No. 1 | No. 2 | No. 3 | No. 6 | No. 7 | ||||
| Arthropoda | Hexanauplia | Calanoida | Pseudodiaptomus sp. | 0.02 | 0.00 | 0.00 | ||||||||||
| Harpacticoida | 43.67 | 97.76 | 6.67 | |||||||||||||
| Harpacticoida | 0.13 | 0.12 | ||||||||||||||
| Amonardia sp. | 0.07 | |||||||||||||||
| Malacostraca | Amphipoda | 57.47 | 58.53 | 96.20 | 10.80 | 0.14 | 0.40 | |||||||||
| Amphipoda | Caprella sp. | 2.12 | 84.35 | 9.33 | 21.58 | 3.37 | 67.37 | 3.88 | 0.15 | 0.13 | ||||||
| Crassicorophium crassicorne | 0.13 | |||||||||||||||
| Gammaridae sp. | 22.03 | 2.90 | ||||||||||||||
| Leucothoe nagatai | 0.01 | |||||||||||||||
| Monocorophium acherusicum | 0.00 | |||||||||||||||
| Isopoda | 7.89 | 13.41 | ||||||||||||||
| Isopoda | Ianiropsis epilittoralis | 92.03 | 97.74 | |||||||||||||
| Ianiropsis sp. | 9.44 | 11.87 | 88.67 | 99.39 | ||||||||||||
| Munna japonica | 15.08 | 0.00 | 0.00 | 0.63 | 0.01 | |||||||||||
| Mysida | 89.02 | 86.59 | ||||||||||||||
| Bryozoa | Gymnolaemata | Ctenostomatida | Amathia verticillate | 0.17 | 0.26 | 0.04 | 0.06 | 0.00 | 0.08 | |||||||
| Chordata | Actinopterygii | Perciformes | Pictichromis paccagnellae | 0.23 | ||||||||||||
| Unknown | 22.36 | 2.10 | 15.39 | 11.42 | 0.15 | 0.18 | 32.55 | 11.33 | 1.04 | 1.48 | 0.07 | |||||
| Total | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.-J.; Kim, H.-W.; Lee, S.-R.; Kim, N.-Y.; Lee, Y.-J.; Joo, H.-T.; Kwak, S.-N.; Lee, S.-H. Feeding Strategy of the Wild Korean Seahorse (Hippocampus haema). J. Mar. Sci. Eng. 2022, 10, 357. https://doi.org/10.3390/jmse10030357
Kim M-J, Kim H-W, Lee S-R, Kim N-Y, Lee Y-J, Joo H-T, Kwak S-N, Lee S-H. Feeding Strategy of the Wild Korean Seahorse (Hippocampus haema). Journal of Marine Science and Engineering. 2022; 10(3):357. https://doi.org/10.3390/jmse10030357
Chicago/Turabian StyleKim, Myung-Joon, Hyun-Woo Kim, Soo-Rin Lee, Na-Yeong Kim, Yoon-Ji Lee, Hui-Tae Joo, Seok-Nam Kwak, and Sang-Heon Lee. 2022. "Feeding Strategy of the Wild Korean Seahorse (Hippocampus haema)" Journal of Marine Science and Engineering 10, no. 3: 357. https://doi.org/10.3390/jmse10030357
APA StyleKim, M. -J., Kim, H. -W., Lee, S. -R., Kim, N. -Y., Lee, Y. -J., Joo, H. -T., Kwak, S. -N., & Lee, S. -H. (2022). Feeding Strategy of the Wild Korean Seahorse (Hippocampus haema). Journal of Marine Science and Engineering, 10(3), 357. https://doi.org/10.3390/jmse10030357