1. Introduction
Knowledge about the functional role of species, communities and ecosystems is crucial for understanding biodiversity patterns and adopting conservation strategies [
1]. Feeding traits have been studied to elucidate fundamental questions in ecology, such as the relationship between many taxonomic groups that share similar ecological functions [
2,
3,
4]. Marine scavengers are those organisms that detect carrion by chemoreception and deliberately move to consume it [
3].
Marine scavengers play an integral role in ecosystem functioning, nutrient cycling and circulating organic carbon [
3,
4,
5]. In this way, scavenger ecology is relevant to understanding trophic relationships and anthropogenic impacts (i.e., how fisheries discards affect benthic carnivores populations) on marine environments [
4]. Most studies on marine carrion consumption have reported that macrofaunal scavengers, especially amphipods, isopods and gastropods, play an important role in organic matter recycling [
1,
6,
7,
8,
9]. Scavenging amphipods can rapidly reach carrion because of their high mobility and well-developed chemoreceptors [
7]. Furthermore, these scavenging amphipods and isopods could play a crucial role in maintaining commercial fish and decapod stocks since they are part of their food sources. Therefore, an increase in the available carrion (e.g., commercial fishing discards) could also raise the biomass of small scavengers and, consequently, the potential food availability for higher levels of the food chain, including species with commercial interest [
10].
Marine scavengers are commonly used as a food resource for people, and fishers have taken advantage of the active scavenging behavior to reach carrion to capture some commercial species in baited traps for decades [
2,
4,
11,
12]. The importance of these fisheries continues to the present; for example, the annual landings of
Octopus vulgaris Cuvier, 1797 in the NW Iberian Peninsula by the artisanal fleet, reaches 1500 t in 2021 [
13,
14]. Therefore, baseline data of scavenging megafauna on the NW coast of the Iberian Peninsula that provides information on the abundance of species in different habitats is key to understanding the distribution of this crucial but understudied group. Moreover, fish damage by scavengers (amphipods and isopods) has been reported as a concern to commercial fisheries and aquaculture production [
15].
Scavenging fauna distribution depends on depth, hydrological conditions, sedimentation regime, food availability and bottom type [
16,
17,
18]. Some approaches have been used: (1) baited cameras that record or take images at pre-set intervals [
2,
5,
19,
20]; (2) small baited traps (1–5 cm entrance) used to sample scavenging macrofauna (0.5 mm to 2 cm) [
1,
9,
16,
17,
18,
21,
22]; and (3) big baited traps (>10 cm entrance) designed for fish, decapods and mollusc harvesting that also sample megafauna (>5 cm) [
10,
12]. The main advantage of baited cameras is that they are non-destructive sampling methods, but it may be difficult or impossible to identify small organisms, such as amphipods or isopods, which are relevant scavenger taxa [
4]. This problem can be dealt with by using baited traps, namely by using traps with a prefixed entrance that allows scavengers to be selected by size, and after the retrieval of traps, smaller macrofauna can be fixed for later identification in the laboratory, whereas the megafauna can be returned to the environment after identification and counting in situ [
9]. Baited traps are usually deployed for a few hours (i.e., 6 h) [
16], 18 h [
9], 24 h [
10,
12,
17,
18], or even for longer periods [
10].
Scavenging fauna is already well-studied in shallow and deep areas of the Artic Sea [
5,
16,
18,
21], shallow waters of the North Atlantic [
2,
12], tropical reef ecosystems of the Coral Sea [
9], the North Deep Pacific [
22], and Antarctica [
1,
19]. Most of these studies focused on commercial fishing discards [
11]. However, studies in the Iberian Peninsula are still scarce and are related to scavenging fauna associated with commercial fishing discards on the southern Portuguese coast [
10]. Nevertheless, these studies were not designed with robust and systematized sampling procedures to compare megafauna and benthopelagic macrofaunal assemblages separately in different habitats under the same depth conditions.
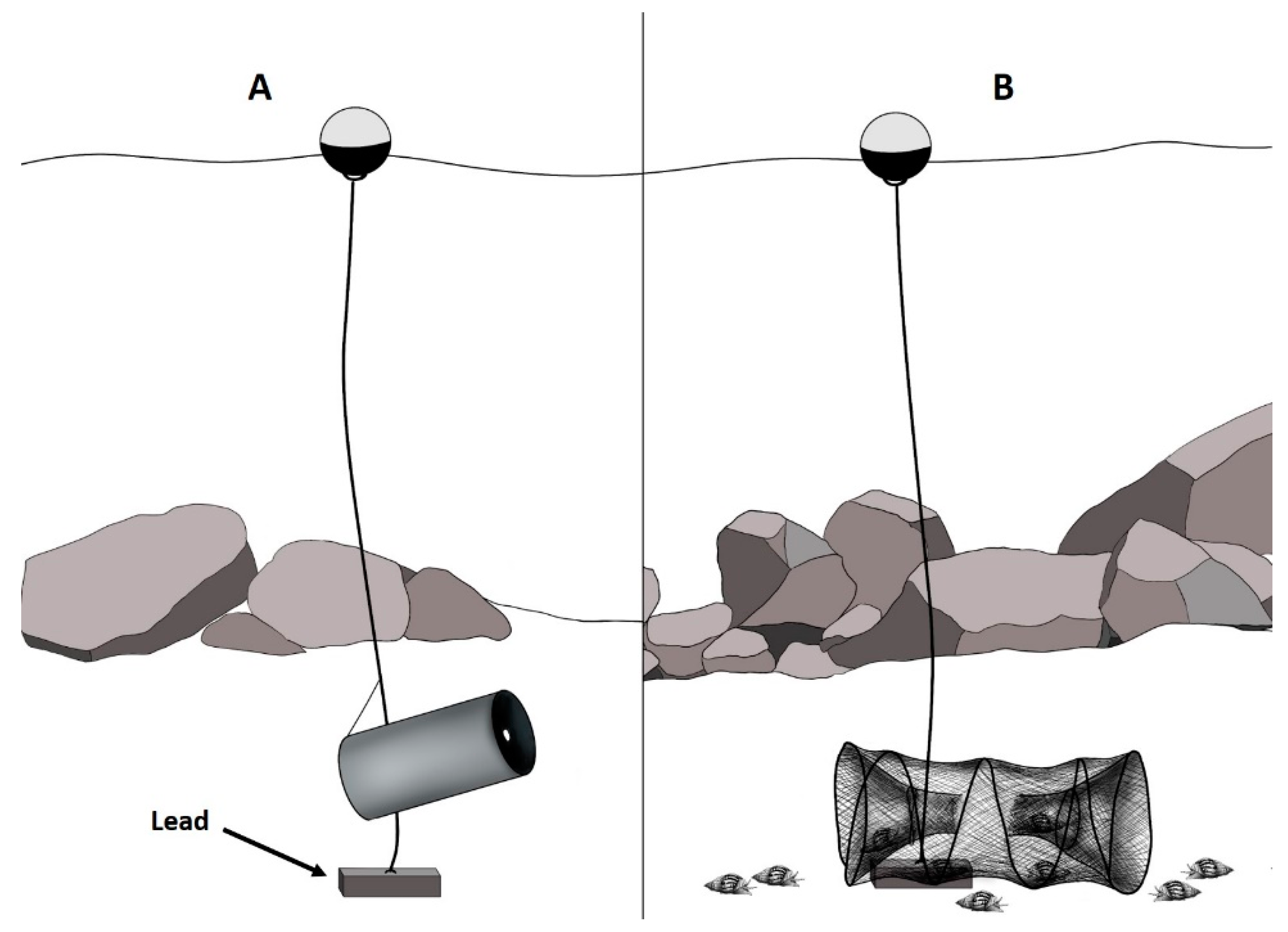
This study aims to set baseline data about scavenger assemblages in three different habitats in shallow waters of the NW Iberian Peninsula. For this, two different types of baited traps, intended to capture scavenging megafaunal and benthopelagic macrofaunal scavenger assemblages, were deployed at 20 m depth on the coast of Viana do Castelo (North Portugal) and to compare them among rocky, pebbled and sandy bottoms. The hypothesis tested was that megafaunal and benthopelagic macrofaunal scavenging assemblages will differ between habitats. These baseline data could improve the knowledge of this understudied group of animals in the scope of the implementation of the European Marine Strategy Framework Directive (MSFD; 2008/56/EC), where the species composition is one basic descriptor on the evaluation of the ‘Good Environmental Status’ (GES) [
23,
24]. Given the economic relevance of some scavenger species, this study could also contribute to fishery management by providing information to fishermen about the habitat preferences of commercial species.
4. Discussion
Results showed that the initial hypothesis was partially supported. On one hand, scavenging megafaunal assemblages differed between sand vs. rock and sand vs. pebbles. On the other, scavenging benthopelagic macrofaunal assemblages differed between pebbles vs. rock. Therefore, differences in the assemblage structure of scavengers among habitats showed different patterns for macrofauna and megafauna, with sand habitats different than the others in megafaunal assemblages and with non-differences between sand vs. pebbles and sand vs. rock in macrofaunal assemblages. This suggests that different drivers seem to shape the structure of species assemblages that we categorized as scavenging benthopelagic macrofaunal and megafaunal assemblages. Regarding megafauna, the dominance of Tritia gastropods in sandy habitats in comparison with its absence in rock and its very low abundance in pebbles was the main factor responsible for differences between habitats. However, in benthopelagic macrofauna, differences in assemblages between pebbles and rocky habitats were more related to changes in the relative abundance of crustacean species (C. cranchii and S. erythrophthalmus) that were less abundant in pebbles.
As it has been verified with the bathymetric maps obtained in this study, the nearby of Viana do Castelo is a very heterogeneous landscape, where sandy, rocky and pebbled habitats are mixed in a relatively small area. Despite this, with this multi-habitat approach, it was possible to observe differences between the habitats based on scavenging megafaunal and benthopelagic macrofaunal assemblages.
Nassariidae gastropods accounted for 97.23% of megafaunal scavengers, almost exclusively in sand habitats. According to Keable, 1995, common obligate scavenger gastropods of the family Nassaridae occurred in large numbers in the marine benthos [
9]. Our results showed that gastropods of the genus
Tritia (
T. reticulata and
T. varicosa) reached the maximum abundance in sandy habitats. This high dominance of
T. reticulata and
T. varicosa on sandy bottoms can be explained because both species’ distribution is limited to sandy habitats [
37]. Other Nassarids such as
Tritia incrassata (Strøm, 1768) are typical scavengers of rocky intertidal habitats, but in this study,
T. incrassata was not found on rocky shores, as this species is limited to low intertidal levels and very shallow subtidal ones [
37]. Therefore, based on our results, the scavenging activity of
Tritia in the shallow sublittoral waters of the NW Iberian Peninsula could be mostly restricted to sandy habitats. This result is also in accordance with previous studies where Nassarids typically present a lie-in-buried-response behavior in soft bottoms [
38,
39].
Sea stars have been reported as successful scavengers in previous studies [
40,
41].
M. glacialis is a chemoreception-based opportunistic feeder and a dominant benthic predator that plays a key role in structuring benthic assemblages [
42]. In our study,
M. glaciaris was the third most abundant megafaunal taxon, appearing in the three studied habitats. Our results agree with Onghia et al., 2014, that reported the preference of
M. glacialis for rocky and boulder bottoms because the abundance of
M. glacialis in rock was more than double compared to the reported abundance in sandy habitats [
42].
For British shallow waters, Nickell and Moore, 1991 reported that a 24 h period of deployment was not enough to collect megafaunal scavengers such as
Octopus or predator fishes in baited traps [
12]. In contrast, our results showed that within 24 h, scavenging macrofaunal species, some with commercial interest in the Iberian Peninsula, were captured, such as
C. conger,
O. vulgaris and
T. luscus [
13,
43,
44,
45].
Conger conger is considered a large opportunistic scavenger and predator typical of shallow and deep rocky areas where it finds refugee and foraging areas [
43,
45]. Our results agree with Castro et al., 2005, who reported
C. conger as the most common fish sampled with bait traps in southern Portugal between 1997 and 2001 [
10]. Our results pointed out a preference of
C. conger for rock and pebbles, in accordance with Xavier et al., 2010, who reported that
C. conger was mostly associated with rocky bottoms on the south coast of the Iberian Peninsula [
45].
C. conger is also a common scavenger of deep and shallow waters and one of the main target species of commercial fishing [
43], with more than 1500 t of annual landings and close to EUR 3 million in profits northwest of the Iberian Peninsula (Galicia) in 2021 [
14].
Gadiformes, and especially those belonging to the family Gadidae, have been reported as typical scavengers captured by baited traps [
40]. In the shallow waters of the NW Iberian Peninsula, one of the most common was
T. luscus. This species is distributed along rock and sand sea bottoms on the continental shelf at depths of 20–100 m [
46]. Despite the fact that the knowledge of its ecology is scarce [
47],
T. luscus has also been reported as one of the most common fish fauna in NW Iberia [
48]. Our results showed that the abundance of
T. luscus was similar to that of
C. conger, also widely distributed along the three studied habitats, but showing a preference for sand as hunting grounds. As
T. luscus is also a commercial species, knowledge about its ecology is also of special relevance; its annual landings reached around 700 t in 2021, with an economic value of direct auction sale of more than EUR 1.7 million [
14].
The very mobile opportunistic carnivore
O. vulgaris is well adapted to live in different habitats (sandy and muddy bottoms, rocks, and seagrass meadows) [
49]. Our results also showed that
O. vulgaris was present in the three studied habitats. Even so, Guerra et al., 2014 reported the sandy inlet of Rodas in the National Park of the Atlantic Galician Islands as a preferential habitat for
O. vulgaris [
49]. In our study,
O. vulgaris was more abundant in sandy bottoms than in rocky and pebbled habitats. Nevertheless, it was observed that the apparent fishing effort in the study area dedicated to
Octopus pots was substantially greater in the rocky habitats than in the sandy ones. This fact, together with the enormous fishing effort in the area, could be influencing our results. Moreover, taking into account that the annual landings of
O. vugaris in the NW Iberian Peninsula in 2021 were more than 1700 t with an economic value of the direct sale of more than EUR 15 million [
14], additional data with regard to
O. vulgaris ecology and distribution could be very helpful for fishers and the scientific community to achieve a more sustainable exploitation of this resource.
Since scavenging megafaunal assemblages included many species with commercial interest, overfishing could also contribute to the decline of these species. In this way, the effect of commercial fishing on scavengers could cause an imbalance in ecosystem functioning and nutrient cycling.
Nickell and Moore, 1991, [
12] pointed out the importance of assuring the non-interference in the baited traps of large scavengers with small ones to not bias the results. Furthermore, according to Legezyńska et al., 2000, some studies based on baited traps were not able to capture both scavenging megafaunal and benthopelagic macrofaunal assemblages [
21]. Thus, the implementation of two different bait traps in this study also guarantees no interference from the megafaunal scavengers with the capture of the macrofaunal ones. Moreover, our results showed different patterns among habitats for scavenging megafaunal and benthopelagic macrofaunal assemblages, reinforcing the implementation of the simultaneous deployment of the two types of baited traps.
The diversity of scavenging benthopelagic macrofaunal assemblages of the NW Iberian Peninsula is difficult to compare to other studies due to differences in trap design, deployment scheme, and target habitats [
9]. Even so, there are common patterns that can be discussed. Within scavenging benthopelagic macrofauna, lysianassoid amphipods and cirolanid isopods are reported as the most important scavengers in playing a key role to transfer the energy of dead organic matter to higher trophic levels [
7,
10]. Lysianasids and Cirolanids had been identified as the most important component for the recycling of organic matter in the deep waters of south Portugal [
10]. Our results showed that
C. cranchii and
S. erythrophthalmus are the most widespread and abundant benthopelagic macrofaunal scavengers in the three habitats. Our results are also in accordance with Keable, 1995 and Ellis et al., 2013, which reported that cirolanids constituted the major portion of the macrofaunal assemblages and that
C. cranchii was highly abundant and also a voracious scavenger [
9,
20].
Keable, 1995, [
9] reported that the composition of scavenger assemblages differs between temperate and cold waters. The NW Iberian Peninsula is an especially important area because it constitutes the transition between two biogeographic regions, the Mediterranean-Atlantic (warm-temperate) and the Boreal-Atlantic (warm-cold). Our results are more similar to those reported in temperate zones where Cirolanids dominate the abundance (i.e., Keable, 1995 [
9]), as compared to those for cold waters, where Lysianassids are usually numerically dominant in baited traps (i.e., [
17,
18,
21]).
In contrast with scavenging megafaunal assemblages, differences between benthopelagic macrofaunal assemblages from pebbles and rocky bottoms were not the result of the presence of exclusive species at any of the two habitats. The two species that explain differences between pebbles and rocky bottoms (i.e., C. cranchii and S. erythrophthalmus) were present in the three studied habitats. Therefore, differences between habitats for benthopelagic macrofauna seem to be explained by changes in the relative abundance of C. cranchii and S. erythrophthalmus between habitats. This result suggests a differential preference of these two species for the studied habitats, rocky shores being the more preferred habitat and pebbles the less preferred one.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}