
Expected Effects of Offshore Wind Farms on Mediterranean Marine Life

 , ,
, ,  ,
,
Abstract
:1. Introduction
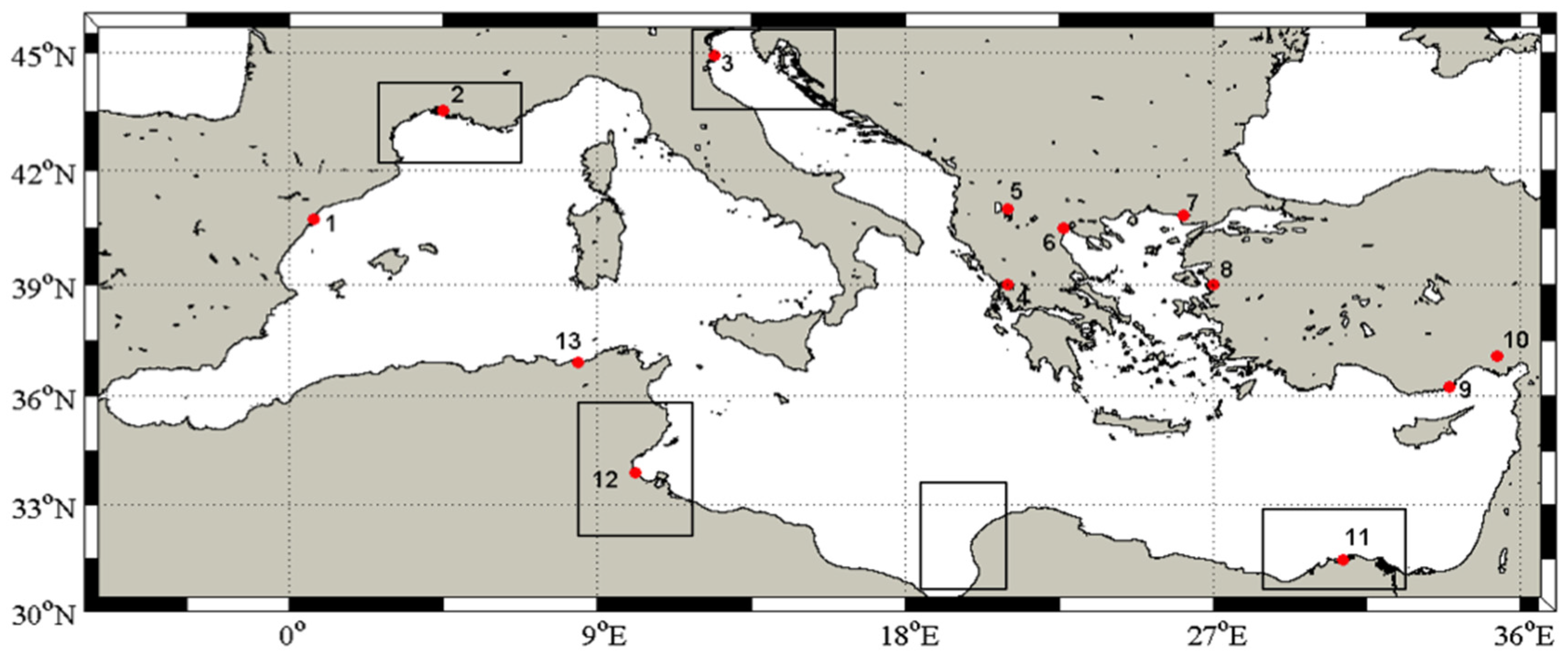
2. Methods
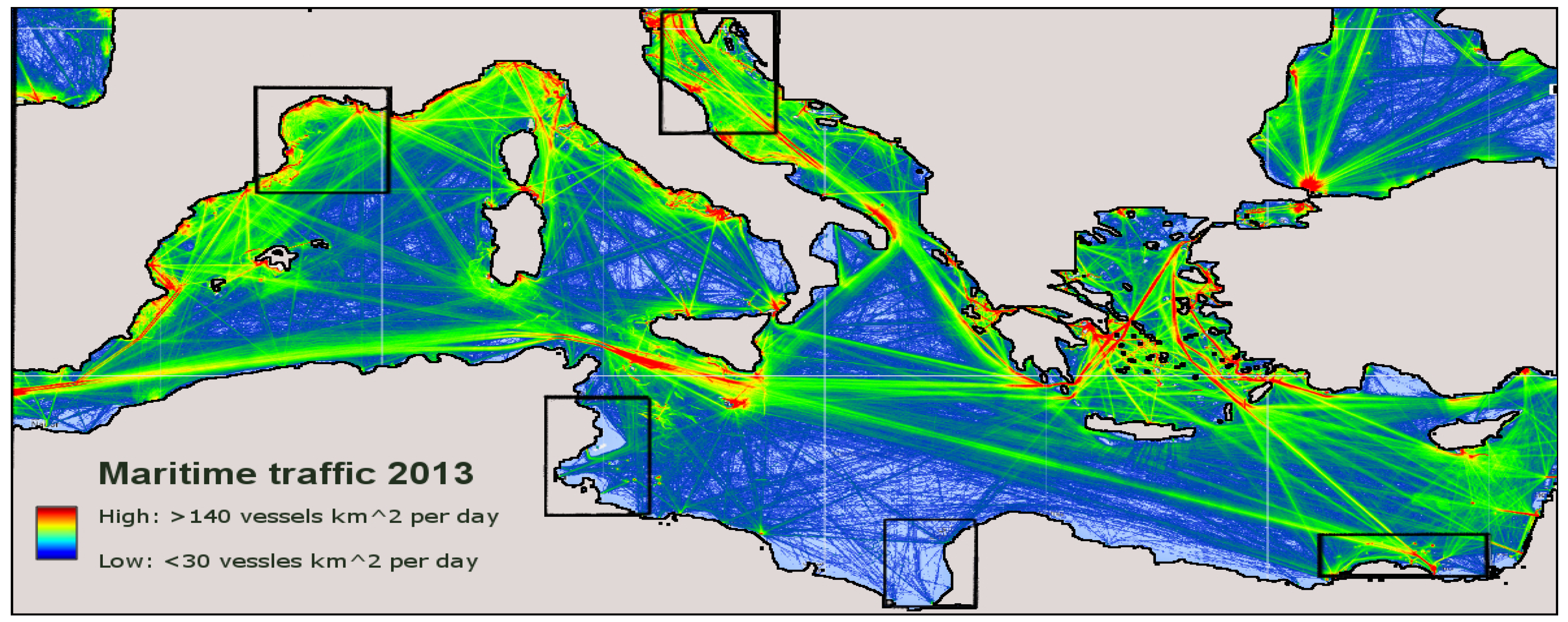
3. Results and Discussion
3.1. Potential Effects of Mediterranean Offshore Wind Farms on Birds
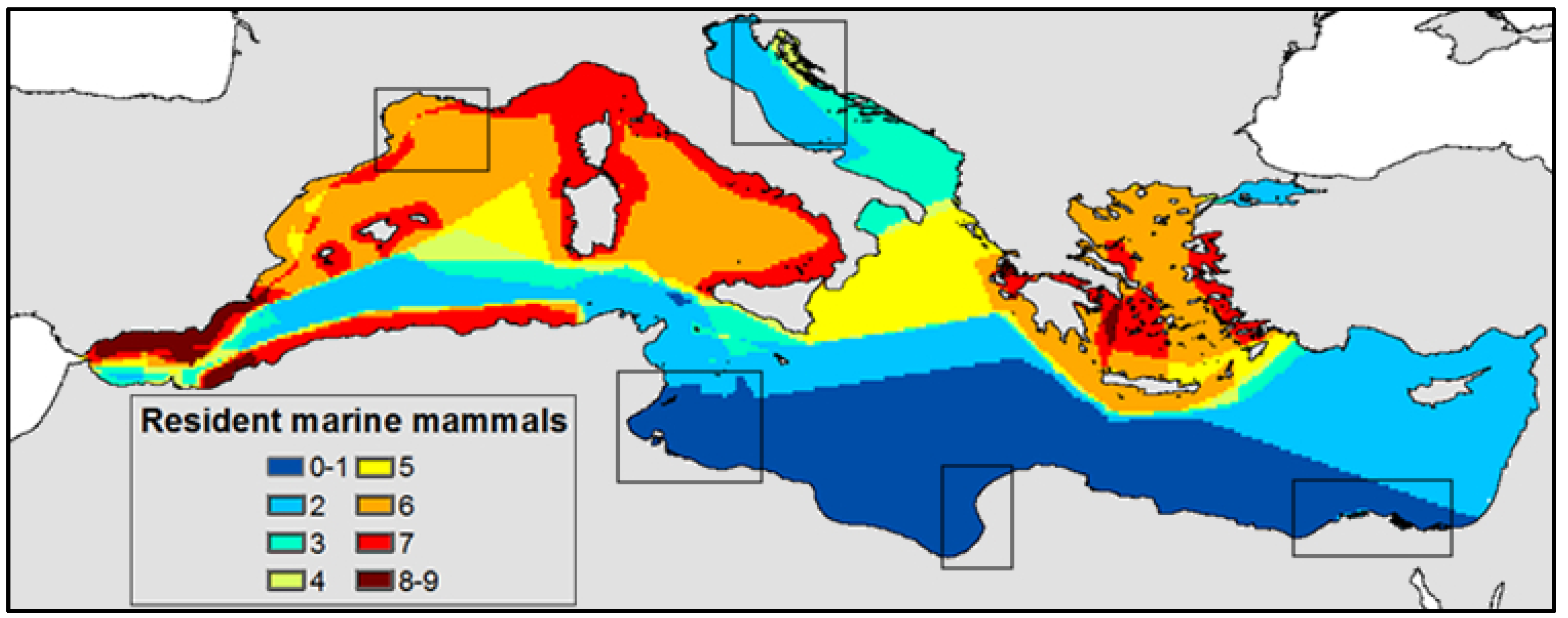
3.2. Potential Effects of Mediterranean Marine Mammals
3.3. Potential Effects on Mediterranean Fish Communities
3.4. Potential Effects of Mediterranean Benthic Communities
3.5. Potential Effects of Mediterranean Planktonic Communities
4. Conclusions
Supplementary Materials
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kermeli, K.; Graus, W.H.J.; Worrell, E. Energy efficiency improvement potentials and a low energy demand scenario for the global industrial sector. Energy Effic. 2014, 7, 987–1011. [Google Scholar] [CrossRef]
- Esseghir, A.; Haouaoui Khouni, L. Economic growth, energy consumption and sustainable development: The case of the Union for the Mediterranean countries. Energy 2014, 71, 218–225. [Google Scholar] [CrossRef]
- Cartalis, C.; Synodinou, A.; Proedrou, M.; Tsangrassoulis, A.; Santamouris, M. Modifications in energy demand in urban areas as a result of climate changes: An assessment for the southeast Mediterranean region. Energy Convers. Manag. 2001, 42, 1647–1656. [Google Scholar] [CrossRef]
- Tsikalakis, A.; Tomtsi, T.; Hatziargyriou, N.D.; Poullikkas, A.; Malamatenios, C.; Giakoumelos, E.; Jaouad, O.C.; Chenak, A.; Fayek, A.; Matar, T.; et al. Review of best practices of solar electricity resources applications in selected Middle East and North Africa (MENA) countries. Renew. Sustain. Energy Rev. 2011, 15, 2838–2849. [Google Scholar] [CrossRef]
- European Wind Energy Association (EWEA); Corbetta, G.; Pineda, I.; Wilkes, J. The European Offshore Wind Industry—Key Trends and Statistics 1st Half 2014; The European Wind Energy Association: Brussels, Belgium, 2014; p. 6. [Google Scholar]
- European Environment Agency (EEA). Europe’s Onshore and Offshore Wind Energy Potential: An Assessment of Environmental and Economic Constraints; EEA: Copenhagen, Denmark, 2009. [Google Scholar]
- Bilgili, M.; Yasar, A.; Simsek, E. Offshore wind power development in Europe and its comparison with onshore counterpart. Renew. Sustain. Energy Rev. 2011, 15, 905–915. [Google Scholar] [CrossRef]
- Jacques, S.; Kreutzkamp, P.; Joseph, P. Seanergy 2020: Analysis of Planned/Suggested Offshore Electricity Infrastructure Relatively to Existing International MSP Instruments 2011; European Wind Energy Association: Brussels, Belgium, 2011. [Google Scholar]
- European Wind Energy Association (EWEA); Moccia, J.; Arapogianni, A.; Wilkes, J.; Kjaer, C.; Gruet, R. Pure Power Wind Energy Targets for 2020 and 2030; European Wind Energy Association: Brussels, Belgium, 2011; p. 96. [Google Scholar]
- De Decker, J.; Kreutzkamp, P.; Joesph, P.; Woyte, A.; Cowdroy, S.; Warland, L.; Svendsen, H.; Völker, J.; Funk, C.; Peinil, H.; et al. Offshore Electricity Grid Infrastructure in Europe; OffshoreGrid Final Report; OffshoreGrid: Brussels, Belgium, 2011. [Google Scholar]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The biodiversity of the Mediterranean Sea: Estimates, patterns, and threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed]
- Zenetos, A.; Gofas, S.; Morri, C.; Rosso, A.; Violanti, D.; Garcia Raso, J.E.; Cinar, M.E.; Almogi-Labin, A.; Ates, A.S.; Azzurro, E.; et al. Alien species in the Mediterranean Sea by 2012. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part 2. Introduction trends and pathways. Mediterr. Mar. Sci. 2012, 13. [Google Scholar] [CrossRef]
- Micheli, F.; Halpern, B.S.; Walbridge, S.; Ciriaco, S.; Ferretti, F.; Fraschetti, S.; Lewison, R.; Nykjaer, L.; Rosenberg, A.A. Cumulative human impacts on mediterranean and black sea marine ecosystems: Assessing current pressures and opportunities. PLoS ONE 2013, 8, e79889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadopoulos, A.; Katsafados, P. Verification of operational weather forecasts from the POSEIDON system across the Eastern Mediterranean. Nat. Hazards Earth Syst. Sci. 2009, 9, 1299–1306. [Google Scholar] [CrossRef]
- Papadopoulos, A.; Kallos, G.; Katsafados, P.; Nickovic, S. The Poseidon weather forecasting system: An overview. Glob. Atmos. Ocean Syst. 2002, 8, 219–237. [Google Scholar] [CrossRef]
- Papadopoulos, A.; Korres, G.; Katsafados, P.; Ballas, D.; Perivoliotis, L.; Nittis, K. Dynamic downscaling of the ERA-40 data using a mesoscale meteorological model. Mediterr. Mar. Sci. 2011, 12. [Google Scholar] [CrossRef]
- Uppala, S.M.; KÅllberg, P.W.; Simmons, A.J.; Andrae, U.; Bechtold, V.D.C.; Fiorino, M.; Gibson, J.K.; Haseler, J.; Hernandez, A.; Kelly, G.A.; et al. The ERA-40 re-analysis. Q. J. R. Meteorol. Soc. 2005, 131, 2961–3012. [Google Scholar] [CrossRef]
- Soukissian, T.H.; Papadopoulos, A. Effects of different wind data sources in offshore wind power assessment. Renew. Energy 2015, 77, 101–114. [Google Scholar] [CrossRef]
- Desholm, M.; Kahlert, J. Avian collision risk at an offshore wind farm. Biol. Lett. 2005, 1, 296–298. [Google Scholar] [CrossRef] [PubMed]
- Scott, B.E.; Langton, R.; Philpott, E.; Waggitt, J.J. Seabirds and Marine Renewables: Are We Asking the Right Questions? In Marine Renewable Energy Technology and Environmental Interactions; Shields, M.A., Payne, A.I.L., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2014; pp. 81–92. [Google Scholar]
- The IUCN Red List of Threatened Species. Version 2014.2. Available online: http://www. iucnredlist.org (accessed on 28 August 2014).
- United Nations Environment Programme-Mediterranean Action Plan (UNEP-MAP). Protocol Concerning Specially Protected Areas and Biological Diversity in the Mediterranean; UNEP/Map Publications: Athens, Greece, 1999; p. 3. [Google Scholar]
- Cooper, J.; Baccetti, N.; Belda, E.J.; Borg, J.J.; Oro, D.; Papaconstantinou, C.; Sánchez, A. Seabird mortality from longline fishing in the Mediterranean Sea and Macaronesian waters: A review and a way forward. Sci. Mar. 2003, 67, 57–64. [Google Scholar] [CrossRef]
- Bourgeois, K.; Vidal, E. The endemic Mediterranean yelkouan shearwater Puffinus yelkouan: Distribution, threats and a plea for more data. Oryx 2008, 42, 187–194. [Google Scholar] [CrossRef]
- Thibault, J.C.; Zotier, R.; Guyot, I.; Bretagnolle, V. Recent trends in breeding marine birds of the Mediterranean region with special reference to Corsica. Colon Waterbirds 1996, 19, 31–40. [Google Scholar] [CrossRef]
- Donald, P.F.; Sanderson, F.J.; Burfield, I.J.; Bierman, S.M.; Gregory, R.D.; Waliczky, Z. International conservation policy delivers benefits for birds in Europe. Science 2007, 317, 810–813. [Google Scholar] [CrossRef] [PubMed]
- Oro, D. Effects of trawler discard availability on egg laying and breeding success in the lesser black-backed gull Larus fuscus in the western Mediterranean. Mar. Ecol. Prog. Ser. 1996, 132, 43–46. [Google Scholar] [CrossRef]
- Oro, D.; Ruiz, X. Exploitation of trawler discards by breeding seabirds in the north-western Mediterranean: Differences between the Ebro Delta and the Balearic Islands areas. ICES J. Mar. Sci. 1997, 54, 695–707. [Google Scholar] [CrossRef]
- Bicknell, A.W.J.; Oro, D.; Camphuysen, K.; Votier, S.C. Potential consequences of discard reform for seabird communities. J. Appl. Ecol. 2013, 50, 649–658. [Google Scholar] [CrossRef]
- Bradbury, G.; Trinder, M.; Furness, B.; Banks, A.N.; Caldow, R.W.G.; Hume, D. Mapping seabird sensitivity to offshore wind farms. PLoS ONE 2014, 9, e106366. [Google Scholar] [CrossRef] [PubMed]
- Furness, R.W.; Wade, H.M.; Masden, E.A. Assessing vulnerability of marine bird populations to offshore wind farms. J. Environ. Manag. 2013, 119, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Garthe, S.; Hüppop, O. Scaling possible adverse effects of marine wind farms on seabirds: Developing and applying a vulnerability index. J. Appl. Ecol. 2004, 41, 724–734. [Google Scholar] [CrossRef]
- Gallo-Orsi, U. Species Action Plans for the conservation of seabirds in the Mediterranean Sea: Audouin’s gull, Balearic shearwater and Mediterranean shag. Sci. Mar. 2003, 67 (Suppl. 2), 47–55. [Google Scholar] [CrossRef]
- Wilcove, D.S.; Wikelski, M. Going, going, gone: Is animal migration disappearing. PLoS Biol. 2008, 6, e188. [Google Scholar] [CrossRef] [PubMed]
- Boere, G.C.; Stroud, D.A. The Flyway Concept: What It Is and What It Isn’t; The Stationery Office: Edinburgh, UK, 2006; pp. 40–47. [Google Scholar]
- Denac, D.; Schneider-Jacoby, M.; Stumberger, B. Adriatic Flyway: Closing the Gap in Bird Conservation; Euronatur: Liboa, Portugal, 2010. [Google Scholar]
- Alerstam, T.; Pettersson, S.G. Why do migrating birds fly along coastlines? J. Theor. Biol. 1977, 65, 699–712. [Google Scholar] [CrossRef]
- Pennycuick, C.J. Soaring behavior and performance of some east African birds, observed from a motor-glider. IBIS 1972, 114, 178–218. [Google Scholar] [CrossRef]
- Bijlsma, R.G. Bottleneck Areas for Migratory Birds in the Mediterranean Region: An Assessment of the Problems and Recommendations for Action; ICBP: Cambridge, UK, 1987; Volume 18. [Google Scholar]
- Kirby, J.S.; Stattersfield, A.J.; Butchart, S.H.M.; Evans, M.I.; Grimmett, R.F.A.; Jones, V.R.; O’Sullivan, J.; Tucker, G.M.; Newton, I. Key conservation issues for migratory land- and waterbird species on the world’s major flyways. Bird Conserv. Int. 2008, 18, S49–S73. [Google Scholar] [CrossRef]
- Jourdain, E.; Gauthier-Clerc, M.; Bicout, D.; Sabatier, P. Bird Migration Routes and Risk for Pathogen Dispersion into Western Mediterranean Wetlands. Emerg Infect Dis 2007, 13, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Hüppop, O.; Dierschke, J.; Exo, K.-M.; Fredrich, E.; Hill, R. Bird migration studies and potential collision risk with offshore wind turbines. IBIS 2006, 148, 90–109. [Google Scholar] [CrossRef]
- Biebach, H.; Biebach, I.; Friedrich, W.; Heine, G.; Partecke, J.; Schmidl, D. Strategies of passerine migration across the Mediterranean Sea and the Sahara Desert: A radar study. IBIS 2000, 142, 623–634. [Google Scholar] [CrossRef]
- Alerstam, T.; Lindström, Å. Optimal bird migration: The relative importance of time, energy, and safety. In Bird Migration; Gwinner, P.D.E., Ed.; Springer: Berlin, Germany, 1990; pp. 331–351. [Google Scholar]
- Gschweng, M.; Kalko, E.K.V.; Querner, U.; Fiedler, W.; Berthold, P. All across Africa: Highly individual migration routes of Eleonora’s falcon. Proc. Biol. Sci. 2008, 275, 2887–2896. [Google Scholar] [CrossRef] [PubMed]
- Wildfowl and Wetlands Trust (WWT). Strategic assessment of collision risk of Scottish offshore wind farms to migrating birds. In Scottish Marine and Freshwater Science; Scottish Government: Edinburgh, UK, 2014; Volume 5. [Google Scholar]
- Masden, E.A.; Reeve, R.; Desholm, M.; Fox, A.D.; Furness, R.W.; Haydon, D.T. Assessing the impact of marine wind farms on birds through movement modelling. J. R. Soc. Interface 2012, 9, 2120–2130. [Google Scholar] [CrossRef] [PubMed]
- Poot, H.; Ens, B.J.; de Vries, H.; Donners, M.A.; Wernand, M.R.; Marquenie, J.M. Green light for nocturnally migrating birds. Ecol. Soc. 2008, 13, 47. [Google Scholar]
- Saidur, R.; Rahim, N.A.; Islam, M.R.; Solangi, K.H. Environmental impact of wind energy. Renew. Sustain. Energy Rev. 2011, 15, 2423–2430. [Google Scholar] [CrossRef]
- Notarbartolo-di-Sciara, G.; Agardy, T.; Hyrenbach, D.; Scovazzi, T.; van Klaveren, P. The Pelagos Sanctuary for Mediterranean marine mammals. Aquat. Conserv. Mar. Freshw. Ecosyst. 2008, 18, 367–391. [Google Scholar] [CrossRef]
- Frantzis, A.; Nikolaou, O.; Bompar, J.M.; Cammedda, A. Humpback whale (Megaptera novaeangliae) occurrence in the Mediterranean Sea. J. Cetacean Res. Manag. 2004, 6, 25–28. [Google Scholar]
- Notarbartolo-Di-Sciara, G.; Zanardelli, M.; Jahoda, M.; Panigada, S.; Airoldi, S. The fin whale Balaenoptera physalus (L. 1758) in the Mediterranean Sea. Mamm. Rev. 2003, 33, 105–150. [Google Scholar] [CrossRef]
- Cañadas, A.; Sagarminaga, R. The Northeastern Alboran Sea, an important breeding and feeding ground for the long-finned pilot whale (Globicephala Melas) in the Mediterranean Sea. Mar. Mammal Sci. 2000, 16, 513–529. [Google Scholar] [CrossRef]
- Frantzis, A.; Airoldi, S.; Notarbartolo-di-Sciara, G.; Johnson, C.; Mazzariol, S. Inter-basin movements of Mediterranean sperm whales provide insight into their population structure and conservation. Deep Sea Res. I 2011, 58, 454–459. [Google Scholar] [CrossRef]
- Bearzi, G.; Fortuna, C.M.; Reeves, R.R. Ecology and conservation of common bottlenose dolphins Tursiops truncatus in the Mediterranean Sea. Mammal Rev. 2009, 39, 92–123. [Google Scholar] [CrossRef]
- Dendrinos, P.; Karamanlidis, A.A.; Kotomatas, S.; Paravas, V.; Adamantopoulou, S. Report of a New Mediterranean monk seal (Monachus monachus) breeding colony in the Aegean Sea, Greece. Aquat. Mamm. 2008, 34, 355–361. [Google Scholar] [CrossRef]
- Gomerčić, T.; Huber, D.; Gomerčić, M.; Gomerčić, H. Presence of the Mediterranean monk seal (Monachus monachus) in the Croatian part of the Adriatic Sea. Aquat. Mamm. 2011, 37, 243–247. [Google Scholar] [CrossRef]
- Thomsen, F.; Lüdemann, K.; Kafemann, R.; Piper, W. Effects of Offshore Wind Farm Noise on Marine Mammals and Fish; Germany on Behalf of COWRIE Ltd.: Hamburg, Germany, 2006. [Google Scholar]
- Madsen, P.; Wahlberg, M.; Tougaard, J.; Lucke, K.; Tyack, P. Wind turbine underwater noise and marine mammals: Implications of current knowledge and data needs. Mar. Ecol. Prog. Ser. 2006, 309, 279–295. [Google Scholar] [CrossRef]
- Bergström, L.; Kautsky, L.; Malm, T.; Rosenberg, R.; Wahlberg, M.; Capetillo, N.Å.; Wilhelmsson, D. Effects of offshore wind farms on marine wildlife—A generalized impact assessment. Environ. Res. Lett. 2014, 9, 034012. [Google Scholar] [CrossRef]
- Bailey, H.; Clay, G.; Coates, E.A.; Lusseau, D.; Senior, B.; Thompson, P.M. Using T-PODs to assess variations in the occurrence of coastal bottlenose dolphins and harbor porpoises. Aquat. Conserv. Mar. Freshw. Ecosyst. 2010, 20, 150–158. [Google Scholar] [CrossRef]
- Carstensen, J.; Henriksen, O.D.; Teilmann, J. Impacts of offshore wind farm construction on harbor porpoises: Acoustic monitoring of echo-location activity using porpoise detectors (T-PODs). Mar. Ecol. Prog. Ser. 2006, 321, 295–308. [Google Scholar] [CrossRef]
- Teilmann, J.; Carstensen, J. Negative long term effects on harbor porpoises from a large scale offshore wind farm in the Baltic—Evidence of slow recovery. Environ. Res. Lett. 2012, 7, 045101. [Google Scholar] [CrossRef]
- Scheidat, M.; Tougaard, J.; Brasseur, S.; Carstensen, J.; van Polanen Petel, T.; Teilmann, J.; Reijnders, P. Harbour porpoises (Phocoena phocoena) and wind farms: A case study in the Dutch North Sea. Environ. Res. Lett. 2011, 6, 025102. [Google Scholar] [CrossRef]
- Russell, D.J.F.; Brasseur, S.M.; Thompson, D.; Hastie, G.D.; Janik, V.M.; Aarts, G.; McClintock, B.T.; Matthiopoulos, J.; Moss, S.E.W.; McConnell, B. Marine mammals trace anthropogenic structures at sea. Curr. Biol. 2014, 24, R638–R639. [Google Scholar] [CrossRef] [PubMed]
- Abdulla, A. Maritime Traffic Effects on Biodiversity in the Mediterranean Sea: Review of Impacts, Priority Areas and Mitigation Measures; IUCN: Gland, Switzerland, 2008; Volume 1. [Google Scholar]
- Bailey, H.; Senior, B.; Simmons, D.; Rusin, J.; Picken, G.; Thompson, P.M. Assessing underwater noise levels during pile-driving at an offshore windfarm and its potential effects on marine mammals. Mar. Pollut. Bull. 2010, 60, 888–897. [Google Scholar] [CrossRef] [PubMed]
- David, J.A. Likely sensitivity of bottlenose dolphins to pile-driving noise. Water Environ. J. 2006, 20, 48–54. [Google Scholar] [CrossRef]
- Tzanatos, E.; Dimitriou, E.; Papaharisis, L.; Roussi, A.; Somarakis, S.; Koutsikopoulos, C. Principal socio-economic characteristics of the Greek small-scale coastal fishermen. Ocean Coast. Manag. 2006, 49, 511–527. [Google Scholar] [CrossRef]
- Malak, D.A. Overview of the Conservation Status of the Marine Fishes of the Mediterranean Sea; IUCN: Gland, Switzerland, 2011. [Google Scholar]
- Vasilakopoulos, P.; Maravelias, C.D.; Tserpes, G. The alarming decline of Mediterranean fish stocks. Curr. Biol. 2014, 24, 1643–1648. [Google Scholar] [CrossRef] [PubMed]
- Debusschere, E.; Coensel, B.D.; Vandendriessche, S.; Botteldooren, D.; Hostens, K.; Vincx, M.; Degraer, S. Effects of offshore wind farms on the early life stages of Dicentrarchus Labrax. In The Effects of Noise on Aquatic Life II; Popper, A.N., Hawkins, A., Eds.; Springer: New York, NY, USA, 2016; pp. 197–204. [Google Scholar]
- Wahlberg, M.; Westerberg, H. Hearing in fish and their reactions to sounds from offshore wind farms. Mar. Ecol. Prog. Ser. 2005, 288, 295–309. [Google Scholar] [CrossRef]
- Sigray, P.; Andersson, M.H. Particle motion measured at an operational wind turbine in relation to hearing sensitivity in fish. J. Acoust. Soc. Am. 2011, 130, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Radford, A.N.; Kerridge, E.; Simpson, S.D. Acoustic communication in a noisy world: Can fish compete with anthropogenic noise? Behav. Ecol. 2014, 25, 1022–1030. [Google Scholar] [CrossRef]
- Tricas, T.; Gill, A.B. Effects of EMFs from Undersea Power Cables on Elasmobranchs and Other Marine Species; U.S. Dept. of the Interior: Camirillo, CA, USA, 2011; p. 426.
- Gill, A.B.; Taylor, H. The Potential Effects of Electromagnetic Fields Generated by Cabling between Offshore Wind Turbines upon Elasmobranch Fishes; Countryside Council for Wales and University of Liverpool: Bangor, UK, 2001; p. 60. [Google Scholar]
- Öhman, M.C.; Sigray, P.; Westerberg, H. Offshore windmills and the effects of electromagnetic fields on fish. AMBIO 2007, 36, 630–633. [Google Scholar] [CrossRef]
- Gill, A.B.; Bartlett, M.; Thomsen, F. Potential interactions between diadromous fishes of U.K. conservation importance and the electromagnetic fields and subsea noise from marine renewable energy developments. J. Fish Biol. 2012, 81, 664–695. [Google Scholar] [CrossRef] [PubMed]
- Westerberg, H.; Lagenfelt, I. Sub-sea power cables and the migration behaviour of the European eel. Fish. Manag. Ecol. 2008, 15, 369–375. [Google Scholar] [CrossRef]
- Krone, R.; Gutow, L.; Joschko, T.J.; Schröder, A. Epifauna dynamics at an offshore foundation—Implications of future wind power farming in the North Sea. Mar. Environ. Res. 2013, 85, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Reubens, J.T.; Braeckman, U.; Vanaverbeke, J.; van Colen, C.; Degraer, S.; Vincx, M. Aggregation at windmill artificial reefs: CPUE of Atlantic cod (Gadus morhua) and pouting (Trisopterus luscus) at different habitats in the Belgian part of the North Sea. Fish. Res. 2013, 139, 28–34. [Google Scholar] [CrossRef]
- Wilhelmsson, D.; Malm, T.; Öhman, M.C. The influence of offshore windpower on demersal fish. ICES J. Mar. Sci. 2006, 63, 775–784. [Google Scholar] [CrossRef]
- Wilson, J.C.; Elliott, M. The habitat-creation potential of offshore wind farms. Wind Energy 2009, 12, 203–212. [Google Scholar] [CrossRef]
- Reubens, J.T.; Degraer, S.; Vincx, M. The ecology of benthopelagic fishes at offshore wind farms: A synthesis of 4 years of research. Hydrobiologia 2014, 727, 121–136. [Google Scholar] [CrossRef]
- De Troch, M.; Reubens, J.T.; Heirman, E.; Degraer, S.; Vincx, M. Energy profiling of demersal fish: A case-study in wind farm artificial reefs. Mar. Environ. Res. 2013, 92, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Reubens, J.T.; de Rijcke, M.; Degraer, S.; Vincx, M. Diel variation in feeding and movement patterns of juvenile Atlantic cod at offshore wind farms. J. Sea Res. 2014, 85, 214–221. [Google Scholar] [CrossRef]
- Bergström, L.; Sundqvist, F.; Bergstrm, U. Effects of an offshore wind farm on temporal and spatial patterns in the demersal fish community. Mar. Ecol. Prog. Ser. 2013, 485, 199–210. [Google Scholar] [CrossRef]
- Wilhelmsson, D.; Langhamer, O. The influence of fisheries exclusion and addition of hard substrata on fish and crustaceans. In Marine Renewable Energy Technology and Environmental Interactions; Shields, M.A., Payne, A.I.L., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2014; pp. 49–60. [Google Scholar]
- Guidetti, P.; Baiata, P.; Ballesteros, E.; di Franco, A.; Hereu, B.; Macpherson, E.; Micheli, F.; Pais, A.; Panzalis, P.; Rosenberg, A.A.; et al. Large-scale assessment of Mediterranean marine protected areas effects on fish assemblages. PLoS ONE 2014, 9, e91841. [Google Scholar] [CrossRef] [PubMed]
- Montefalcone, M.; Albertelli, G.; Morri, C.; Parravicini, V.; Bianchi, C.N. Legal protection is not enough: Posidonia oceanica meadows in marine protected areas are not healthier than those in unprotected areas of the northwest Mediterranean Sea. Mar. Pollut. Bull. 2009, 58, 515–519. [Google Scholar] [CrossRef] [PubMed]
- Halpern, B.S.; Gaines, S.D.; Warner, R.R. Confounding effects of the export of production and the displacement of fishing effort from marine reserves. Ecol. Appl. 2004, 14, 1248–1256. [Google Scholar] [CrossRef]
- Slabbekoorn, H.; Bouton, N.; van Opzeeland, I.; Coers, A.; ten Cate, C.; Popper, A.N. A noisy spring: The impact of globally rising underwater sound levels on fish. Trends Ecol. Evol. 2010, 25, 419–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danovaro, R.; Company, J.B.; Corinaldesi, C.; D’Onghia, G.; Galil, B.; Gambi, C.; Gooday, A.J.; Lampadariou, N.; Luna, G.M.; Morigi, C.; et al. Deep-sea biodiversity in the Mediterranean Sea: The known, the unknown, and the unknowable. PLoS ONE 2010, 5, e11832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, G.; Agnesi, S.; Evans, D.; Populus, J.; Tunesi, L. Mediterranean benthic EUNIS habitats: Structural considerations and lessons learned from mapping. Rev. Investig. Mar. 2012, 19, 48–50. [Google Scholar]
- Vassallo, P.; Paoli, C.; Rovere, A.; Montefalcone, M.; Morri, C.; Bianchi, C.N. The value of the seagrass Posidonia oceanica: A natural capital assessment. Mar. Pollut. Bull. 2013, 75, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Jackson, E.L.; Rees, S.E.; Wilding, C.; Attrill, M.J. Use of a seagrass residency index to apportion commercial fishery landing values and recreation fisheries expenditure to seagrass habitat service. Conserv. Biol. 2015, 29, 899–909. [Google Scholar] [CrossRef] [PubMed]
- Sanchezjerez, P. Effect of an artificial reef in Posidonia meadows on fish assemblage and diet of Diplodus annularis. ICES J. Mar. Sci. 2002, 59, S59–S68. [Google Scholar] [CrossRef]
- Coates, D.A.; Deschutter, Y.; Vincx, M.; Vanaverbeke, J. Enrichment and shifts in macrobenthic assemblages in an offshore wind farm area in the Belgian part of the North Sea. Mar. Environ. Res. 2014, 95, 1–12. [Google Scholar] [CrossRef] [PubMed]
- De Backer, A.; van Hoey, G.; Coates, D.; Vanaverbeke, J.; Hostens, K. Similar diversity-disturbance responses to different physical impacts: Three cases of small-scale biodiversity increase in the Belgian part of the North Sea. Mar. Pollut. Bull. 2014, 84, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Birklund, J.; Petersen, A.H. Development of the Fouling Community on Turbine Foundations and Scour Protections in Nysted Offshore Wind Farm, 2003; Report Energi E2 A/S; DHI Water and Environment: Hørsholm, Denmark, 2004. [Google Scholar]
- Kerckhof, F.; Norro, A.; Jacques, T.; Degraer, S. Early colonization of a concrete offshore windmill foundation by marine biofouling on the Thornton Bank (southern North Sea). In Offshore Wind Farms in the Belgian Part of the North Sea: State of the Art after Two Years of Environmental Monitoring; Royal Belgian Institute of Natural Sciences: Brussel, Belgium, 2009. [Google Scholar]
- Kerckhof, F.; Rumes, B.; Norro, A.; Jacques, T.G.; Degraer, S. Seasonal variation and vertical zonation of the marine biofouling on a concrete offshore windmill foundation on the Thornton Bank (southern North Sea). In Offshore Wind Farms in the Belgian Part of the North Sea: State of the Art after Two Years of Environmental Monitoring; Royal Belgian Institute of Natural Sciences: Brussel, Belgium, 2010. [Google Scholar]
- Kerckhof, F.; Rumes, B.; Jacques, T.; Degraer, S.; Norro, A. Early development of the subtidal marine biofouling on a concrete offshore windmill foundation on the Thornton Bank (southern North Sea): First monitoring results. Underw. Technol. 2010, 29, 137–149. [Google Scholar] [CrossRef]
- Langhamer, O. Wave Energy Conversion and the Marine Environment: Colonization Patterns and Habitat Dynamics. Ph.D. Thesis, Uppsala University, Uppsala, Sweden, 2009. [Google Scholar]
- Lindeboom, H.J.; Kouwenhoven, H.J.; Bergman, M.J.N.; Bouma, S.; Brasseur, S.; Daan, R.; Fijn, R.C.; de Haan, D.; Dirksen, S.; van Hal, R. Short-term ecological effects of an offshore wind farm in the Dutch coastal zone; a compilation. Environ. Res. Lett. 2011, 6, 035101. [Google Scholar] [CrossRef]
- Maar, M.; Bolding, K.; Petersen, J.K.; Hansen, J.L.S.; Timmermann, K. Local effects of blue mussels around turbine foundations in an ecosystem model of Nysted off-shore wind farm, Denmark. J. Sea Res. 2009, 62, 159–174. [Google Scholar] [CrossRef]
- Shi, W.; Park, H.C.; Baek, J.H.; Kim, C.W.; Kim, Y.C.; Shin, H.K. Study on the marine growth effect on the dynamic response of offshore wind turbines. Int. J. Precis. Eng. Manuf. 2012, 13, 1167–1176. [Google Scholar] [CrossRef]
- Van Hoey, G.; Degraer, S.; Vincx, M. Macrobenthic community structure of soft-bottom sediments at the Belgian Continental Shelf. Estuar. Coast. Shelf Sci. 2004, 59, 599–613. [Google Scholar] [CrossRef]
- Wilhelmsson, D.; Malm, T. Fouling assemblages on offshore wind power plants and adjacent substrata. Estuar. Coast. Shelf Sci. 2008, 79, 459–466. [Google Scholar] [CrossRef]
- Langhamer, O. Artificial reef effect in relation to offshore renewable energy conversion: State of the art. Sci. World J. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Fabi, G.; Luccarini, F.; Panfili, M.; Solustri, C.; Spagnolo, A. Effects of an artificial reef on the surrounding soft-bottom community (central Adriatic Sea). ICES J. Mar. Sci. 2002, 59, S343–S349. [Google Scholar] [CrossRef]
- Fabi, G.; Fiorentini, L. Comparison between an artificial reef and a control site in the Adriatic Sea: Analysis of four years of monitoring. Bull. Mar. Sci. 1994, 55, 538–558. [Google Scholar]
- Sinis, A.I.; Chintiroglou, C.C.; Stergiou, K.I. Preliminary results from the establishment of experimental artificial reefs in the N. Aegean Sea (Chalkidiki, Greece). Belg. J. Zool. 2000, 130, 143–147. [Google Scholar]
- Airoldi, L.; Bulleri, F. Anthropogenic disturbance can determine the magnitude of opportunistic species responses on marine urban infrastructures. PLoS ONE 2011, 6, e22985. [Google Scholar] [CrossRef] [PubMed]
- Bulleri, F.; Airoldi, L. Artificial marine structures facilitate the spread of a non-indigenous green alga, Codium fragile ssp. tomentosoides, in the north Adriatic Sea. J. Appl. Ecol. 2005, 42, 1063–1072. [Google Scholar] [CrossRef]
- Goren, M. Development of benthic community on artificial substratum at Ashdod (Eastern Mediterranean). Oceanol. Acta 1980, 3, 275–283. [Google Scholar]
- Kocak, F.; Ergen, Z.; Çinar, M.E. Fouling organisms and their developments in a polluted and an unpolluted marina in the Aegean Sea (Turkey). Ophelia 1999, 50, 1–20. [Google Scholar] [CrossRef]
- Relini, G.; Relini, M.; Montanari, M. An offshore buoy as a small artificial island and a fish-aggregating device (FAD) in the Mediterranean. In Island, Ocean and Deep-Sea Biology; Jones, M.B., Azevedo, J.M.N., Neto, A.I., Costa, A.C., Martins, A.M.F., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2000; pp. 65–80. [Google Scholar]
- Moreno, I.; Roca, I.; Reñones, O.; Coll, J.; Salamanca, M. Artificial reef program in Balearic waters (western Mediterranean). Bull. Mar. Sci. 1994, 55, 667–671. [Google Scholar]
- Relini, G.; Tixi, F.; Relini, M.; Torchia, G. The macrofouling on offshore platforms at Ravenna. Int. Biodeterior. Biodegrad. 1998, 41, 41–55. [Google Scholar] [CrossRef]
- Badalamenti, F. Are artificial reefs comparable to neighbouring natural rocky areas? A mollusc case study in the Gulf of Castellammare (NW Sicily). ICES J. Mar. Sci. 2002, 59, S127–S131. [Google Scholar] [CrossRef]
- Relini, G.; Zamboni, N.; Tixi, F.; Torchia, G. Patterns of sessile Macrobenthos community development on an artificial reef in the gulf of genoa (Northwestern Mediterranean). Bull. Mar. Sci. 1994, 55, 745–771. [Google Scholar]
- Ponti, M.; Fava, F.; Perlini, R.A.; Giovanardi, O.; Abbiati, M. Benthic assemblages on artificial reefs in the northwestern Adriatic Sea: Does structure type and age matter? Mar. Environ. Res. 2015, 104, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti, L.; Marzialetti, S.; Paganelli, D.; Ardizzone, G.D. Long-term changes in a benthic assemblage associated with artificial reefs. In Biodiversity in Enclosed Seas and Artificial Marine Habitats; Relini, G., Ryland, J., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2007; pp. 233–240. [Google Scholar]
- Galil, B.S.; Boero, F.; Campbell, M.L.; Carlton, J.T.; Cook, E.; Fraschetti, S.; Gollasch, S.; Hewitt, C.L.; Jelmert, A.; Macpherson, E.; et al. “Double trouble”: The expansion of the Suez Canal and marine bioinvasions in the Mediterranean Sea. Biol. Invasions 2014, 17, 973–976. [Google Scholar] [CrossRef] [Green Version]
- Çinar, M.E. Serpulid species (Polychaeta: Serpulidae) from the Levantine coast of Turkey (eastern Mediterranean), with special emphasis on alien species. Aquat. Invasions 2006, 1, 223–240. [Google Scholar] [CrossRef]
- Adams, T.P.; Miller, R.G.; Aleynik, D.; Burrows, M.T. Offshore marine renewable energy devices as stepping stones across biogeographical boundaries. J. Appl. Ecol. 2014, 51, 330–338. [Google Scholar] [CrossRef]
- Siokou-Frangou, I.; Christaki, U.; Mazzocchi, M.G.; Montresor, M.; Ribera d’Alcalá, M.; Vaqué, D.; Zingone, A. Plankton in the open Mediterranean Sea: A review. Biogeosciences 2010, 7, 1543–1586. [Google Scholar] [CrossRef] [Green Version]
- Cerino, F.; Bernardi Aubry, F.; Coppola, J.; La Ferla, R.; Maimone, G.; Socal, G.; Totti, C. Spatial and temporal variability of pico-, nano- and microphytoplankton in the offshore waters of the southern Adriatic Sea (Mediterranean Sea). Cont. Shelf Res. 2012, 44, 94–105. [Google Scholar] [CrossRef]
- Shields, M.A.; Woolf, D.K.; Grist, E.P.M.; Kerr, S.A.; Jackson, A.C.; Harris, R.E.; Bell, M.C.; Beharie, R.; Want, A.; Osalusi, E.; et al. Marine renewable energy: The ecological implications of altering the hydrodynamics of the marine environment. Ocean Coast. Manag. 2011, 54, 2–9. [Google Scholar] [CrossRef]
- Broström, G. On the influence of large wind farms on the upper ocean circulation. J. Mar. Syst. 2008, 74, 585–591. [Google Scholar] [CrossRef]
- Paskyabi, M.B.; Fer, I. Upper ocean response to large wind farm effect in the presence of surface gravity waves. Energy Procedia 2012, 24, 245–254. [Google Scholar] [CrossRef]
- Adams, T.P.; Aleynik, D.; Burrows, M. Larval dispersal of intertidal organisms and the influence of coastline geography. Ecography 2014, 37, 698–710. [Google Scholar] [CrossRef]
- Inger, R.; Attrill, M.J.; Bearhop, S.; Broderick, A.C.; James Grecian, W.; Hodgson, D.J.; Mills, C.; Sheehan, E.; Votier, S.C.; Witt, M.J. Marine renewable energy: Potential benefits to biodiversity? An urgent call for research. J. Appl. Ecol. 2009, 46, 1145–1153. [Google Scholar] [CrossRef]
- Miller, R.G.; Hutchison, Z.L.; Macleod, A.K.; Burrows, M.T.; Cook, E.J.; Last, K.S.; Wilson, B. Marine renewable energy development: Assessing the Benthic Footprint at multiple scales. Front. Ecol. Environ. 2013, 11, 433–440. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name | Species | Endemic | Listed under Barcelona Convention | Wind Farm Sensitivity Index [13] | Vulnerability Index for Collision Impacts [30] | Vulnerability Index for Disturbance Impacts [30] |
|---|---|---|---|---|---|---|
| Cory’s Shearwater (Mediterranean) | Calonectris diomedea diomedea | Y | Y | - | - | - |
| Yelkouan Shearwater (Mediterranean) | Puffinus yelkouan | Y | N | - | - | - |
| Balearic Shearwater | Puffinus mauretanicus | Y | N | - | - | - |
| European Shag (Mediterranean) | Phalacrocorax aristotelis desmarestii | Y | Y | - | 150 | 14 |
| Great Comorant | Phalacrocorax carbo | N | N | 23.3 | - | - |
| Pygmy Comorant | Phalacrocorax pygmeus | N | N | - | - | - |
| Audouin’s gull | Larus audouinii | N | Y | - | - | - |
| Little Gull | Hydrcoleus minutus | N | N | 12.8 | - | - |
| Lesser black-backed gull | Larus fuscus | N | N | 13.8 | 960 | 3 |
| Slender billed gull | Larus genei | N | Y | - | - | - |
| Mediterranean gull | Larus melanocephalus | N | Y | - | - | - |
| Black-headed gull | Larus ridibundus | N | N | 7.5 | - | - |
| Caspian gull | Larus cachinnans | N | N | - | - | - |
| Black legged kittiwake | Rissa tridactyla | N | N | 7.5 | - | |
| Yellow legged gull | Larus michahellis | N | N | - | - | - |
| Great skua | Catharacta skua | N | N | - | 320 | 3 |
| Caspian tern | Hydroprogne caspia | N | N | - | - | - |
| Common tern | Sterna hirundo | N | N | 15.0 | 229 | 8 |
| Little tern | Sterna albifrons | N | N | - | 212 | 10 |
| Sandwich tern | Sterna sandvicensis | N | N | 25.0 | 245 | 9 |
| Lesser-crested tern | Thalasseus bengalensis | N | N | - | - | |
| Razorbill | Alca torda | N | N | 15.8 | 32 | 14 |
| Atlantic puffin | Fratercula arctica | N | N | 15.0 | 27 | 10 |
| European Storm petrel | Hydrobates pelagic melitensis | Y | Y | - | 91 | 2 |
| Northern gannet | Morus bassanus | N | N | - | - | - |
| Osprey | Pandion haliaetus | N | Y | - | - | - |
| Eleanore’s falcon | Falco eleonorae | N | Y | - | - | - |
| Red throated diver | Gavia stellata | N | N | 43.3 | 213 | 32 |
| Black throated diver | Gavia arctica | N | N | 40.3 | 240 | 32 |
| Great crested grebe | Podicep scristatus | N | N | 19.3 | 84 | 8 |
| Red-necked grebe | Podiceps grisegena | N | N | 18.7 | - | - |
| Eared grebe | Podiceps nigricollis | N | N | - | - | - |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bray, L.; Reizopoulou, S.; Voukouvalas, E.; Soukissian, T.; Alomar, C.; Vázquez-Luis, M.; Deudero, S.; Attrill, M.J.; Hall-Spencer, J.M. Expected Effects of Offshore Wind Farms on Mediterranean Marine Life. J. Mar. Sci. Eng. 2016, 4, 18. https://doi.org/10.3390/jmse4010018
Bray L, Reizopoulou S, Voukouvalas E, Soukissian T, Alomar C, Vázquez-Luis M, Deudero S, Attrill MJ, Hall-Spencer JM. Expected Effects of Offshore Wind Farms on Mediterranean Marine Life. Journal of Marine Science and Engineering. 2016; 4(1):18. https://doi.org/10.3390/jmse4010018
Chicago/Turabian StyleBray, Laura, Sofia Reizopoulou, Evangelos Voukouvalas, Takvor Soukissian, Carme Alomar, Maite Vázquez-Luis, Salud Deudero, Martin J. Attrill, and Jason M. Hall-Spencer. 2016. "Expected Effects of Offshore Wind Farms on Mediterranean Marine Life" Journal of Marine Science and Engineering 4, no. 1: 18. https://doi.org/10.3390/jmse4010018
APA StyleBray, L., Reizopoulou, S., Voukouvalas, E., Soukissian, T., Alomar, C., Vázquez-Luis, M., Deudero, S., Attrill, M. J., & Hall-Spencer, J. M. (2016). Expected Effects of Offshore Wind Farms on Mediterranean Marine Life. Journal of Marine Science and Engineering, 4(1), 18. https://doi.org/10.3390/jmse4010018