The Effects of Ocean Acidification and Warming on Growth of a Natural Community of Coastal Phytoplankton

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Measurement of Parameters
2.3. Statistical Analysis
3. Results
3.1. Carbonate Parameters and Temperature Dynamics
3.2. Bloom Development
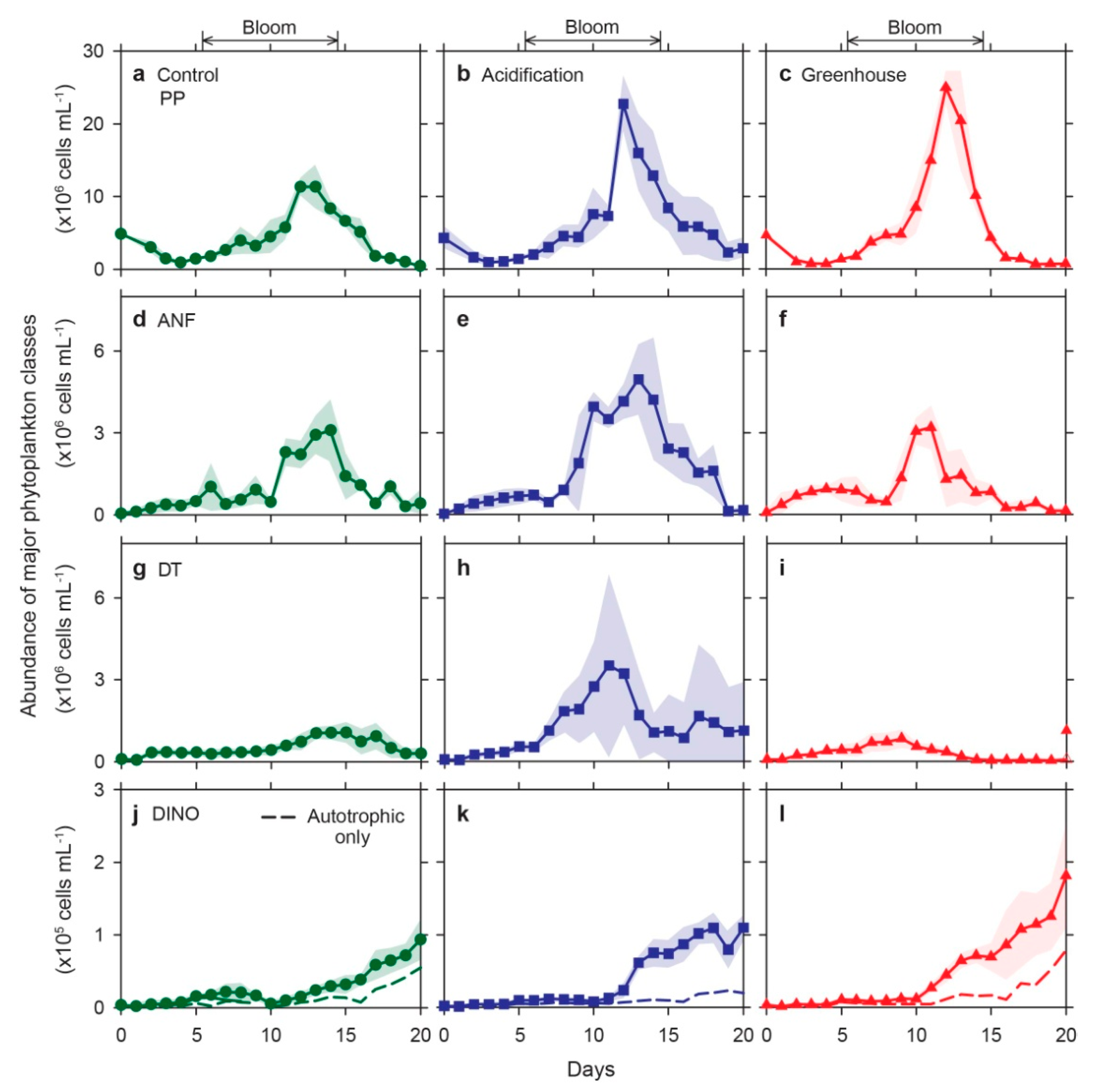
3.3. Population Dynamics of Major Phytoplankton Classes
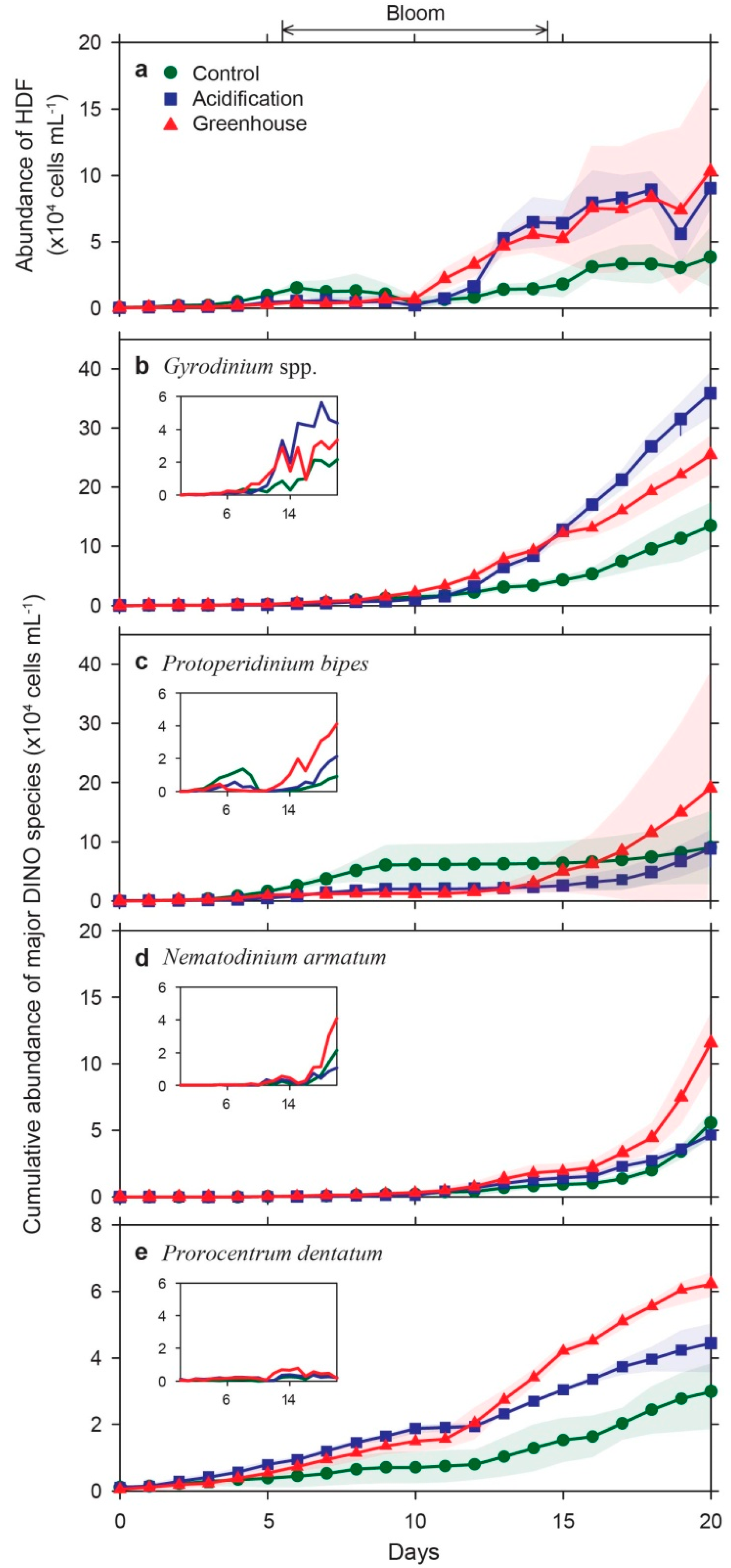
3.4. Species-Specific Growth Response of Diatoms and Dinoflagellates
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2014: Mitigation of Climate Change. Contribution of Working Group III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Edenhofer, O., Pichs-Madruga, R., Sokona, Y., Farahani, E., Kadner, S., Seyboth, K., Adler, A., Baum, I., Brunner, S., Eickemeier, P., et al., Eds.; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Caldeira, K.; Wickett, M.E. Anthropogenic carbon and ocean pH. Nature 2003, 425, 365. [Google Scholar] [CrossRef] [PubMed]
- Sommer, U.; Paul, C.; Moustaka-Gouni, M. Warming and ocean acidification effects on phytoplankton from species shifts to size shifts within species in a mesocosm experiment. PLoS ONE 2015, 10, e0125239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Field, C.B.; Barros, V.R.; Dokken, D.; Mach, K.; Mastrandrea, M.; Bilir, T.; Cbatterjee, M.; Ebi, K.; Estrada, Y.; Genova, R. IPCC 2014: Summary for policymakers in Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. 2014, pp. 1–32. Available online: https://www.ipcc.ch/report/ar5/wg2/ (accessed on 18 October 2020).
- Beardall, J.; Stojkovic, S. Microalgae under global environmental change: Implications for growth and productivity, populations and trophic flow. ScienceAsia 2006, 32, 1–10. [Google Scholar] [CrossRef]
- Boyd, P.W.; Doney, S.C. Modelling regional responses by marine pelagic ecosystems to global climate change. Geophys. Res. Lett. 2002, 29, 53-1–53-4. [Google Scholar] [CrossRef] [Green Version]
- Hays, G.C.; Richardson, A.J.; Robinson, C. Climate change and marine plankton. Trends Ecol. Evol. 2005, 20, 337–344. [Google Scholar] [CrossRef]
- Hein, M.; Sand-Jensen, K. CO2 increases oceanic primary production. Nature 1997, 388, 526–527. [Google Scholar] [CrossRef]
- Schippers, P.; Lürling, M.; Scheffer, M. Increase of atmospheric CO2 promotes phytoplankton productivity. Ecol. Lett. 2004, 7, 446–451. [Google Scholar] [CrossRef]
- Tortell, P.D.; Payne, C.D.; Li, Y.; Trimborn, S.; Rost, B.; Smith, W.O.; Riesselman, C.; Dunbar, R.B.; Sedwick, P.; DiTullio, G.R. CO2 sensitivity of Southern Ocean phytoplankton. Geophys. Res. Lett. 2008, 35. [Google Scholar] [CrossRef] [Green Version]
- Egge, J.; Thingstad, J.; Larsen, A.; Engel, A.; Wohlers, J.; Bellerby, R.; Riebesell, U. Primary production during nutrient-induced blooms at elevated CO2 concentrations. Biogeosci. Discuss. 2009, 6, 877–885. [Google Scholar] [CrossRef] [Green Version]
- Engel, A.; Borchard, C.; Piontek, J.; Schulz, K.G.; Riebesell, U.; Bellerby, R. CO2 increases 14C primary production in an Arctic plankton community. Biogeosci. Discuss. 2013, 10, 1291–1308. [Google Scholar] [CrossRef] [Green Version]
- Fu, F.-X.; Zhang, Y.; Warner, M.E.; Feng, Y.; Sun, J.; Hutchins, D.A. A comparison of future increased CO2 and temperature effects on sympatric Heterosigma akashiwo and Prorocentrum minimum. Harm. Algae 2008, 7, 76–90. [Google Scholar] [CrossRef]
- Sciandra, A.; Harlay, J.; Lefèvre, D.; Lemée, R.; Rimmelin, P.; Denis, M.; Gattuso, J.-P. Response of coccolithophorid Emiliania huxleyi to elevated partial pressure of CO2 under nitrogen limitation. Mar. Ecol. Prog. Ser. 2003, 261, 111–122. [Google Scholar] [CrossRef]
- Delille, B.; Harlay, J.; Zondervan, I.; Jacquet, S.; Chou, L.; Wollast, R.; Bellerby, R.G.; Frankignoulle, M.; Borges, A.V.; Riebesell, U. Response of primary production and calcification to changes of pCO2 during experimental blooms of the coccolithophorid Emiliania huxleyi. Glob. Biogeochem. Cycles 2005, 19. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-M.; Lee, K.; Shin, K.; Kang, J.-H.; Lee, H.-W.; Kim, M.; Jang, P.-G.; Jang, M.-C. The effect of seawater CO2 concentration on growth of a natural phytoplankton assemblage in a controlled mesocosm experiment. Limnol. Oceanogr. 2006, 51, 1629–1636. [Google Scholar] [CrossRef] [Green Version]
- Hare, C.E.; Leblanc, K.; DiTullio, G.R.; Kudela, R.M.; Zhang, Y.; Lee, P.A.; Riseman, S.; Hutchins, D.A. Consequences of increased temperature and CO2 for phytoplankton community structure in the Bering Sea. Mar. Ecol. Prog. Ser. 2007, 352, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-H.; Kim, K.; Kang, E.; Lee, K.; Kim, J.-M.; Park, K.; Shin, K.; Hyun, B.; Jeong, H.J. Enhancement of photosynthetic carbon assimilation efficiency of phytoplankton assemblage in the future coastal ocean. Biogeosci. Discuss. 2013, 10. [Google Scholar] [CrossRef]
- Sommer, U.; Lewandowska, A. Climate change and the phytoplankton spring bloom: Warming and overwintering zooplankton have similar effects on phytoplankton. Glob. Chang. Biol. 2011, 17, 154–162. [Google Scholar] [CrossRef]
- Brussaard, C.P.D.; Noordeloos, A.A.M.; Witte, H.; Collenteur, M.C.J.; Schulz, K.; Ludwig, A.; Riebesell, U. Arctic microbial community dynamics influenced by elevated CO2 levels. Biogeosci. Discuss. 2013, 10, 719–731. [Google Scholar] [CrossRef] [Green Version]
- Gao, K.; Helbling, E.W.; Häder, D.-P.; Hutchins, D.A. Responses of marine primary producers to interactions between ocean acidification, solar radiation, and warming. Mar. Ecol. Prog. Ser. 2012, 470, 167–189. [Google Scholar] [CrossRef] [Green Version]
- Riebesell, U.; Bellerby, R.; Grossart, H.-P.; Thingstad, F. Mesocosm CO2 perturbation studies: From organism to community level. Biogeosci. Discuss. 2008, 5, 1157–1164. [Google Scholar] [CrossRef] [Green Version]
- Riebesell, U.; Czerny, J.; Bröckel, K.; Boxhammer, T.; Büdenbender, J.; Deckelnick, M.; Fischer, M.; Hoffmann, D.; Krug, S.; Lentz, U. A mobile sea-going mesocosm system–new opportunities for ocean change research. Biogeosci. Discuss. 2013, 10, 1835–1847. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Climate Change 2007: Contribution of the Working Group III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Metz, B., Davidson, O.R., Bosch, P.R., Dave, R., Meyer, L.A., Eds.; IPCC: Cambridge, UK; New York, NY, USA, 2007. [Google Scholar]
- Solomon, S. The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Intergovernmental Panel on Climate Change (IPCC), Climate Change 2007. 2007. Available online: https://www.ipcc.ch/report/ar4/wg1/ (accessed on 18 October 2020).
- Kim, J.-M.; Lee, K.; Shin, K.; Yang, E.J.; Engel, A.; Karl, D.M.; Kim, H.-C. Shifts in biogenic carbon flow from particulate to dissolved forms under high carbon dioxide and warm ocean conditions. Geophys. Res. Lett. 2011, 38. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-M.; Shin, K.; Lee, K.; Park, B.K. In situ ecosystem-based carbon dioxide perturbation experiments: Design and performance evaluation of a mesocosm facility. Limnol. Oceanogr. Methods 2008, 6, 208–217. [Google Scholar] [CrossRef] [Green Version]
- Parsons, T.R. A Manual of Chemical & Biological Methods for Seawater Analysis; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Mehrbach, C.; Culberson, C.H.; Hawley, J.E.; Pytkowicx, R.M. Measurement of the apparent dissociation constants of carbonic acid in seawater at atmospheric pressure. Limnol. Oceanogr. 1973, 18, 897–907. [Google Scholar] [CrossRef]
- Fong, M.B.; Dickson, A.G. Insights from GO-SHIP hydrography data into the thermodynamic consistency of CO2 system measurements in seawater. Mar. Chem. 2019, 211, 52–63. [Google Scholar] [CrossRef]
- Lee, K.; Millero, F.J.; Byrne, R.H.; Feely, R.A.; Wanninkhof, R. The recommended dissociation constants for carbonic acid in seawater. Geophys. Res. Lett. 2000, 27, 229–232. [Google Scholar] [CrossRef]
- Lueker, T.J.; Dickson, A.G.; Keeling, C.D. Ocean pCO2 calculated from dissolved inorganic carbon, alkalinity, and equations for K1 and K2: Validation based on laboratory measurements of CO2 in gas and seawater at equilibrium. Mar. Chem. 2000, 70, 105–119. [Google Scholar] [CrossRef]
- McElligott, S.; Byrne, R.H.; Lee, K.; Wanninkhof, R.; Millero, F.J.; Freely, R.A. Discrete water column measurements of CO2 fugacity and pHT in seawater: A comparison of direct measurements and thermodynamic calculations. Mar. Chem. 1998, 60, 63–73. [Google Scholar] [CrossRef]
- Millero, F.J.; Graham, T.B.; Huang, F.; Bustos-Serrano, H.; Pierrot, D. Dissociation constants of carbonic acid in seawater as a function of salinity and temperature. Mar. Chem. 2006, 100, 80–94. [Google Scholar] [CrossRef]
- Rines, J.B. The Chaetoceros Ehrenberg (Baciullariophyceae) flora of Narragansett Bay, Rhode Island, USA. Bibl. Phycol. 1988, 79, 1–196. [Google Scholar]
- Tomas, C.R. Identifying Marine Phytoplankton; Elsevier: Amsterdam, The Netherlands, 1997. [Google Scholar]
- Lee, K.; Millero, F.J.; Campbell, D.M. The reliability of the thermodynamic constants for the dissociation of carbonic acid in seawater. Mar. Chem. 1996, 55, 233–245. [Google Scholar] [CrossRef]
- Takahashi, T.; Sutherland, S.C.; Sweeney, C.; Poisson, A.; Metzl, N.; Tilbrook, B.; Bates, N.; Wanninkhof, R.; Feely, R.A.; Sabine, C.; et al. Global sea–air CO2 flux based on climatological surface ocean pCO2, and seasonal biological and temperature effects. Deep Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 1601–1622. [Google Scholar] [CrossRef]
- Falkowski, P.; Raven, J. Aquatic Photosynthesis; Princeton University Press: Princeton, NJ, USA, 2007. [Google Scholar]
- Raven, J.A.; Giordano, M.; Beardall, J.; Maberly, S.C. Algal and aquatic plant carbon concentrating mechanisms in relation to environmental change. Photosynth. Res. 2011, 109, 281–296. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Li, Y.; Wu, Y.; Huang, B.; Dai, M.; Fu, F.; Hutchins, D.A.; Gao, K. Effects of elevated CO2 on phytoplankton during a mesocosm experiment in the southern eutrophicated coastal water of China. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Low-Décarie, E.; Bell, G.; Fussmann, G.F. CO2 alters community composition and response to nutrient enrichment of freshwater phytoplankton. Oecologia 2015, 177, 875–883. [Google Scholar] [CrossRef]
- Horn, H.G.; Sander, N.; Stuhr, A.; Alguero-Muniz, M.; Bach, L.T.; Loder, M.G.J.; Boersma, M.; Riebesell, U.; Aberle, N. Low CO2 sensitivity of microzooplankton communities in the Gullmar Fjord, Skagerrak: Evidence from a long-term mesocosm study. PLoS ONE 2016, 11, e0165800. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Hernández, N.; Bach, L.T.; Montero, M.F.; Taucher, J.; Baños, I.; Guan, W.; Esposito, M.; Ludwig, A.; Achterberg, E.P.; Riebesell, U. High CO2 under nutrient fertilization increases primary production and biomass in subtropical phytoplankton communities: A mesocosm approach. Front. Mar. Sci. 2018, 5, 213. [Google Scholar] [CrossRef]
- Rose, J.M.; Feng, Y.; Gobler, C.J.; Gutierrez, R.; Hare, C.E.; Leblanc, K.; Hutchins, D.A. Effects of increased pCO2 and temperature on the North Atlantic spring bloom. II. Microzooplankton abundance and grazing. Mar. Ecol. Prog. Ser. 2009, 388, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Calbet, A.; Landry, M.R. Phytoplankton growth, microzooplankton grazing, and carbon cycling in marine systems. Limnol. Oceanogr. 2004, 49, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Artamonova, E.; Svenning, J.; Vasskog, T.; Hansen, E.; Eilertsen, H. Analysis of phospholipids and neutral lipids in three common northern cold water diatoms: Coscinodiscus concinnus, Porosira glacialis, and Chaetoceros socialis, by ultra-high performance liquid chromatography-mass spectrometry. J. Appl. Phycol. 2017, 29, 1241–1249. [Google Scholar] [CrossRef]
- Yoder, J.A. Effect of temperature on light-limited growth and chemical composition of Skeletonema costatum (BACILLARIOPHYCEAE) 1. J. Phycol. 1979, 15, 362–370. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytoplankton Classes | Species | Specific Growth Rate (d−1) ± s.d. | Mean Cell Abundance (103 Cells L−1) ± s.d. | |||||
|---|---|---|---|---|---|---|---|---|
| Control | Acidification | Greenhouse | Control | Acidification | Greenhouse | F Value | ||
| Picophytoplankton (PP) | 0.26 ± 0.02 | 0.40 ± 0.02 | 0.42 ± 0.02 | 5864 ± 1209 A | 8894 ± 1332 A,B | 10,421 ± 1,595 B | 3.3 * | |
| Autotrophic nanoflagellate (ANF) | 0.22 ± 0.05 | 0.26 ± 0.03 | 0.26 ± 0.13 | 1541 ± 261 A | 2743 ± 392 B | 1435 ± 159 A | 6.8 ** | |
| Diatoms (DT) | 0.26 ± 0.01 | 0.32 ± 0.15 | 0.26 ± 0.13 | 576 ± 141 A | 1976 ± 1297 B | 478 ± 115 A | 19.3 *** | |
| Skeletonema spp. | 0.60 ± 0.25 | 0.73 ± 0.11 | 0.62 ± 0.04 | 127 ± 53 A | 1298 ± 673 B | 315 ± 111 A | 21.7 *** | |
| Chaetoceros socialis | 0.48 ± 0.12 | 0.65 ± 0.14 | 0.32 ± 0.23 | 59 ± 32 A | 211 ± 311 B | 5 ± 4 A | 6.3 ** | |
| Cerataulina spp. | 0.49 ± 0.12 | 0.41 ± 0.00 | 0.31 ± 0.06 | 66 ± 12 B | 17 ± 17 A | 5 ± 1 A | 14.3 *** | |
| Chaetoceros decipiens | 0.52 ± 0.04 | 0.39 ± 0.08 | 0.44 ± 0.04 | 74 ± 25 B | 27 ± 12 A | 10 ± 3 A | 17.6 *** | |
| Eucampia zodiacus | 0.70 ± 0.25 | 0.53 ± 0.03 | 0.46 ± 0.06 | 24 ± 7 B | 9 ± 3 A | 2 ± 0 A | 7.3 ** | |
| Dinoflagellates (DINO) | 0.20 ± 0.03 | 0.35 ± 0.02 | 0.27 ± 0.02 | 60 ± 23 A | 93 ± 16 B | 114 ± 39 B | 14.1 *** | |
| Gyrodinium spp. | 0.41 ± 0.06 | 0.50 ± 0.14 | 0.41 ± 0.15 | 16.8 ± 0.6 A | 45.8 ± 0.8 C | 26.9 ± 0.4 B | 40.7 *** | |
| Protoperidinium bipes | 0.50 ± 0.06 | 0.58 ± 0.10 | 0.62 ± 0.23 | 4.6 ± 0.4 A | 10.9 ± 0.4 A | 26.7 ± 0.4 B | 7.7 * | |
| Nematodinium armatum | 0.59 ± 0.15 | 0.56 ± 0.27 | 0.70 ± 0.06 | 7.9 ± 0.2 A,B | 5.6 ± 0.1 A | 16.3 ± 0.3 B | 5.1 * | |
| Prorocentrum dentatum | 0.51 ± 0.21 | 0.63 ± 0.15 | 0.71 ± 0.08 | 2.8 ± 0.0 | 2.9 ± 0.1 | 4.7 ± 0.1 | n.s. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hyun, B.; Kim, J.-M.; Jang, P.-G.; Jang, M.-C.; Choi, K.-H.; Lee, K.; Yang, E.J.; Noh, J.H.; Shin, K. The Effects of Ocean Acidification and Warming on Growth of a Natural Community of Coastal Phytoplankton. J. Mar. Sci. Eng. 2020, 8, 821. https://doi.org/10.3390/jmse8100821
Hyun B, Kim J-M, Jang P-G, Jang M-C, Choi K-H, Lee K, Yang EJ, Noh JH, Shin K. The Effects of Ocean Acidification and Warming on Growth of a Natural Community of Coastal Phytoplankton. Journal of Marine Science and Engineering. 2020; 8(10):821. https://doi.org/10.3390/jmse8100821
Chicago/Turabian StyleHyun, Bonggil, Ja-Myung Kim, Pung-Guk Jang, Min-Chul Jang, Keun-Hyung Choi, Kitack Lee, Eun Jin Yang, Jae Hoon Noh, and Kyoungsoon Shin. 2020. "The Effects of Ocean Acidification and Warming on Growth of a Natural Community of Coastal Phytoplankton" Journal of Marine Science and Engineering 8, no. 10: 821. https://doi.org/10.3390/jmse8100821
APA StyleHyun, B., Kim, J. -M., Jang, P. -G., Jang, M. -C., Choi, K. -H., Lee, K., Yang, E. J., Noh, J. H., & Shin, K. (2020). The Effects of Ocean Acidification and Warming on Growth of a Natural Community of Coastal Phytoplankton. Journal of Marine Science and Engineering, 8(10), 821. https://doi.org/10.3390/jmse8100821