Plasticity and Multiplicity of Trophic Modes in the Dinoflagellate Karlodinium and Their Pertinence to Population Maintenance and Bloom Dynamics



Abstract
:1. Introduction
2. Trophic Modes of Dinoflagellates
3. Autotrophy of Karlodinium
4. Osmotrophy of Karlodinium
5. Phagotrophy of Karlodinium
5.1. Karlodinium veneficum
5.2. Karlodinium australe
5.3. Karlodinium armiger
6. Evolution of the Feeding Mechanisms in Karlodinium
7. Mixotrophy in Regulating Population Dynamics and HABs Formation of Karlodinium
8. Perspectives for Future Investigations on the Mixotrophy in Karlodinium
8.1. The Ecophysiology of Karlodinium Under Global Changes
8.2. Molecular Basis of Phagotrophy-Relevant Genes in Karlodinium and Other Species
8.3. Energetics and Pathways Relevant to the Energy Metabolisms of Phagocytosis of Karlodinium
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Charles, B.M.; Patricia, A.W. Biological Oceanography, 2nd ed.; Wiley-Blackwell: Chichester, UK, 2012; pp. 1–464. [Google Scholar]
- Long, S.P.; Humphries, S.; Falkowski, P.G. Photoinhibition of photosynthesis in nature. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1994, 45, 633–662. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Barber, R.T.; Smetacek, V. Biogeochemical controls and feedbacks on ocean primary production. Science 1998, 281, 200–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burkholder, J.A.M.; Glibert, P.M.; Skelton, H.M. Mixotrophy, a major mode of nutrition for harmful algal species in eutrophic waters. Harmful Algae 2008, 8, 77–93. [Google Scholar] [CrossRef]
- Gomez, F. A quantitative review of the lifestyle, habitat and trophic diversity of dinoflagellates (Dinoflagellata, Alveolata). Syst. Biodivers. 2012, 10, 267–275. [Google Scholar] [CrossRef]
- Lin, S.; Cheng, S.; Song, B.; Zhong, X.; Lin, X.; Li, W.; Li, L.; Zhang, Y.; Zhang, H.; Ji, Z.; et al. The Symbiodinium kawagutii genome illuminates dinoflagellate gene expression and coral symbiosis. Science 2015, 350, 691–694. [Google Scholar] [CrossRef] [Green Version]
- Smayda, T.J. Harmful algal blooms: Their ecophysiology and general relevance to phytoplankton blooms in the sea. Limnol. Oceanog. 1997, 42, 1137–1153. [Google Scholar] [CrossRef]
- Cullen, J.J.; Yang, X.; Macintyre, H.L. Nutrient limitation of marine photosynthesis. In Primary Productivity and Biogeochemical Cycles in the Sea; Falkowski, P.G., Ed.; Plenum Press: New York, NY, USA, 1992; Volume 43, pp. 69–88. [Google Scholar]
- Tang, E.P.Y. Why do dinoflagellates have lower growth rates? J. Phycol. 1996, 32, 80–84. [Google Scholar] [CrossRef]
- Jones, R.I. Mixotrophy in Planktonic protists as a spectrum of nutritional strategies. Mar. Microbial Food Webs 1994, 8, 87–96. [Google Scholar]
- Jones, R.I. Mixotrophy in planktonic protists: An overview. Freshw. Biol. 2000, 45, 219–226. [Google Scholar] [CrossRef]
- Hammer, A.C.; Pitchford, J.W. The role of mixotrophy in plankton bloom dynamics, and the consequences for productivity. ICES J. Mar. Sci. 2005, 62, 833–840. [Google Scholar] [CrossRef]
- Gaines, G. Heterotrophic nutrition. Biology of Dinoflagellates. In The biology of dinoflagellates; Taylor, F.J.R., Ed.; Blackwell: Oxford, UK, 1987; pp. 224–267. [Google Scholar]
- Skovgaard, A. Mixotrophy in Fragilidium subglobosum (Dinophyceae): Growth and grazing responses as functions of light intensity. Mar. Ecol. Prog. 1996, 143, 247–253. [Google Scholar] [CrossRef]
- Daugbjerg, N.; Hansen, G.; Larsen, J.; Moestrup, O. Phylogeny of some of the major genera of dinoflagellates based on ultrastructure and partial LSU rDNA sequence data, including the erection of three new genera of unarmoured dinoflagellates. Phycologia 2000, 39, 302–317. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Hu, Z.; Shang, L.; Deng, Y.; Tang, Y.Z. A strain of the toxic dinoflagellate Karlodinium veneficum isolated from the East China Sea is an omnivorous phagotroph. Harmful Algae 2020, 93, 101775. [Google Scholar] [CrossRef] [PubMed]
- Hallegraeff, G.; Mooney, B.; Evans, K.; Hosja, W. What Triggers Fish-Killing Karlodinium veneficum Dinoflagellate Blooms in the Swan Canning River System; Swan Canning Research and Innovation Program Final Report; Swan River Trust: Perth, Australia, 2010; p. 31. [Google Scholar]
- Lim, H.C.; Leaw, C.P.; Tan, T.H.; Kon, N.F.; Yek, L.H.; Hii, K.S.; Teng, S.T.; Razali, R.M.; Usup, G.; Iwataki, M.; et al. A bloom of Karlodinium australe (Gymnodiniales, Dinophyceae) associated with mass mortality of cage-cultured fishes in West Johor Strait, Malaysia. Harmful Algae 2014, 40, 51–62. [Google Scholar] [CrossRef]
- Adolf, J.E.; Bachvaroff, T.R.; Krupatkina, D.N.; Nonogaki, H.; Brown, P.J.P.; Lewitus, A.J.; Harvey, H.R.; Place, A.R. Species specificity and potential roles of Karlodinium micrum toxin. Afr. J. Mar. Sci. 2006, 28, 415–419. [Google Scholar] [CrossRef]
- Li, A.S.; Stoecker, D.K.; Adolf, J.E. Feeding, pigmentation, photosynthesis and growth of the mixotrophic dinoflagellate Gyrodinium galatheanum. Aquat. Microb. Ecol. 1999, 19, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Li, A.S.; Stoecker, D.K.; Coats, D.W. Mixotrophy in Gyrodinium galatheanum (Dinophyceae): Grazing responses to light intensity and inorganic nutrients. J. Phycol. 2000, 36, 33–45. [Google Scholar] [CrossRef]
- Li, A.S.; Stoecker, D.K.; Coats, D.W. Spatial and temporal aspects of Gyrodinium galatheanum in Chesapeake Bay: Distribution and mixotrophy. J. Plankton Res. 2000, 22, 2105–2124. [Google Scholar] [CrossRef]
- Nielsen, M.V. Growth and chemical composition of the toxic dinoflagellate Gymnodinium galatheanum in relation to irradiance, temperature and salinity. Mar. Ecol. Prog. 1996, 136, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Delgado, M.; Alcaraz, M. Interactions between red tide microalgae and herbivorous zooplankton: The noxious effects of Gyrodinium corsicum (Dinophyceae) on Acartia grani (Copepoda: Calanoida). J. Plankton Res. 1999, 21, 2361–2371. [Google Scholar] [CrossRef]
- Deeds, J.R.; Terlizzi, D.E.; Adolf, J.E.; Stoecker, D.K.; Place, A.R. Toxic activity from cultures of Karlodinium micrum (=Gyrodinium galatheanum) (Dinophyceae)—a dinoflagellate associated with fish mortalities in an estuarine aquaculture facility. Harmful Algae 2002, 1, 169–189. [Google Scholar] [CrossRef]
- Fernández-Tejedor, M.; Soubrier-Pedreño, M.Á.; Ma, D.F. Acute LD of a Gyrodinium corsicum natural population for Sparus aurata and Dicentrarchus labrax. Harmful Algae 2004, 3, 1–9. [Google Scholar] [CrossRef]
- Da Costa, M.R.; Franco, J.; Cacho, E.; Fernández, F. Toxin content and toxic effects of the dinoflagellate Gyrodinium corsicum (Paulmier) on the ingestion and survival rates of the copepods Acartia grani and Euterpina acutifrons. J. Exp. Mar. Biol. Ecol. 2005, 322, 177–183. [Google Scholar] [CrossRef]
- de Salas, M.F.; Bolch, C.J.S.; Hallegraeff, G.M. Karlodinium australe sp. nov. (Gymnodiniales, Dinophyceae), a new potentially ichthyotoxic unarmoured dinoflagellate from lagoonal habitats of south-eastern Australia. Phycologia 2005, 44, 640–650. [Google Scholar] [CrossRef]
- Garcés, E.; Fernandez, M.; Penna, A.; Lenning, K.V.; Gutierrez, A.; Camp, J.; Zapata, M. Characterization of NW Mediterranean Karlodinium spp. (Dinophyceae) strains using morphological, molecular, chemical, and physiological methodologies. J. Phycol. 2006, 42, 1096–1112. [Google Scholar] [CrossRef]
- Adolf, J.E.; Bachvaroff, T.R.; Deeds, J.R.; Place, A.R. Ichthyotoxic Karlodinium veneficum (Ballantine) J Larsen in the upper Swan River estuary (Western Australia): Ecological conditions leading to a fish kill. Harmful Algae 2015, 48, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Van Wagoner, R.M.; Deeds, J.R.; Tatters, A.O.; Place, A.R.; Tomas, C.R.; Wright, J.L.C. Structure and relative potency of several karlotoxins from Karlodinium veneficum. J. Nat. Prod. 2010, 73, 1360–1365. [Google Scholar] [CrossRef] [Green Version]
- Cai, P.; He, S.; Zhou, C.; Place, A.R.; Haq, S.; Ding, L.; Chen, H.; Jiang, Y.; Guo, C.; Xu, Y.; et al. Two new karlotoxins found in Karlodinium veneficum (strain GM2) from the East China Sea. Harmful Algae 2016, 58, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Krock, B.; Busch, J.A.; Tillmann, U.; Garcia-Camacho, F.; Sanchez-Miron, A.; Gallardo-Rodriguez, J.J.; Lopez-Rosales, L.; Andree, K.B.; Fernandez-Tejedor, M.; Witt, M.; et al. LC-MS/MS detection of karlotoxins reveals new variants in strains of the marine dinoflagellate Karlodinium veneficum from the Ebro Delta (NW Mediterranean). Mar. Drugs 2017, 15. [Google Scholar] [CrossRef] [Green Version]
- Mooney, B.D.; de Salas, M.; Hallegraeff, G.M.; Place, A.R. Survey for karlotoxin production in 15 species of gymnodinioid dinoflagellates (Kareniaceae, Dinophyta). J. Phycol. 2009, 45, 164–175. [Google Scholar] [CrossRef]
- Mooney, B.D.; Hallegraeff, G.M.; Place, A.R. Ichthyotoxicity of four species of gymnodinioid dinoflagellates (Kareniaceae, Dinophyta) and purified karlotoxins to larval sheepshead minnow. Harmful Algae 2010, 9, 557–562. [Google Scholar] [CrossRef]
- Andersen, A.; De, L.M.; Binzer, S.B.; Rasmussen, S.A.; Hansen, P.J.; Nielsen, K.F.; Jørgensen, K.; Larsen, T.O. HPLC-HRMS quantification of the ichthyotoxin karmitoxin from Karlodinium armiger. Mar Drugs 2017, 15, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, S.A.; Binzer, S.B.; Hoeck, C.; Meier, S.; de Medeiros, L.S.; Andersen, N.G.; Place, A.; Nielsen, K.F.; Hansen, P.J.; Larsen, T.O. Karmitoxin: An amine-containing polyhydroxy-polyene toxin from the marine dinoflagellate Karlodinium armiger. J. Nat. Prod. 2017, 80, 1287–1293. [Google Scholar] [CrossRef] [PubMed]
- Bergholtz, T.; Daugbjerg, N.; Moestrup, Ø.; Fernández-Tejedor, M. On the identity of Karlodinium veneficum and description of Karlodinium armiger sp. nov. (Dinophyceae), based on light and electron microscopy, nuclear-encoded LSU rDNA, and pigment composition. J. Phycol. 2005, 42, 170–193. [Google Scholar] [CrossRef]
- Toldrà, A.; Jauset-Rubio, M.; Andree, K.B.; Fernández-Tejedor, M.; Diogène, J.; Katakis, I.; O’Sullivan, C.K.; Campàs, M. Detection and quantification of the toxic marine microalgae Karlodinium veneficum and Karlodinium armiger using recombinase polymerase amplification and enzyme-linked oligonucleotide assay. Anal. Chim. Acta 2018, 1039, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Berge, T.; Hansen, P.J.; Moestrup, O. Prey size spectrum and bioenergetics of the mixotrophic dinoflagellate Karlodinium armiger. Aquat. Microb. Ecol. 2008, 50, 289–299. [Google Scholar] [CrossRef] [Green Version]
- Berge, T.; Poulsen, L.K.; Moldrup, M.; Daugbjerg, N.; Hansen, P.J. Marine microalgae attack and feed on metazoans. Isme J. 2012, 6, 1926–1936. [Google Scholar] [CrossRef]
- Leong, S.C.Y.; Lim, L.P.; Chew, S.M.; Kok, J.W.K.; Teo, S.L.M. Three new records of dinoflagellates in Singapore’s coastal waters, with observations on environmental conditions associated with microalgal growth in the Johor Straits. Raffles Bull. Zool. 2015, 24–36. [Google Scholar]
- De Salas, M.F.; Laza-Martínez, A.; Hallegraeff, G.M. Novel unarmored dinoflagellates from the toxigenic family Kareniaceae (Gymnodiniales): Five new species of Karlodinium and one new Takayama from the Australian sector of the southern ocean. J. Phycol. 2008, 44, 241–257. [Google Scholar] [CrossRef]
- Benico, G.; Takahashi, K.; Lum, W.M.; Yniguez, A.T.; Iwataki, M. The harmful unarmored dinoflagellate Karlodiniumin Japan and Philippines, with reference to ultrastructure and micropredation of Karlodinium azanzae sp. nov. (Kareniaceae, Dinophyceae). J. Phycol. 2020, 56, 1264–1282. [Google Scholar] [CrossRef]
- Escobar-Morales, S.; Hernandez-Becerril, D.U. Free-living marine planktonic unarmoured dinoflagellates from the Gulf of Mexico and the Mexican Pacific. Botanica Marina 2015, 58, 9–22. [Google Scholar] [CrossRef]
- Zingone, A.; Siano, R.; D’Alelio, D.; Sarno, D. Potentially toxic and harmful microalgae from coastal waters of the Campania region (Tyrrhenian Sea, Mediterranean Sea). Harmful Algae 2006, 5, 321–337. [Google Scholar] [CrossRef]
- Paulmier, G.; Berland, B.; Billard, C.; Nezan, E. Gyrodinium corsicum nov. sp. (Gymnodiniales, Dinophycean), responsible organism of the green water in salt-water lake of Diana (Corsica), in April 1994. Cryptogamie Algol. 1995, 16, 77–94. [Google Scholar]
- Cen, J.; Wang, J.; Huang, L.; Ding, G.; Qi, Y.; Cao, R.; Cui, L.; Lv, S. Who is the"murderer"of the bloom in coastal waters of Fujian, China, in 2019? J. Oceanol. Limnol. 2020, 38, 722–732. [Google Scholar] [CrossRef]
- Yang, Z.B.; Takayama, H.; Matsuoka, K.; Hodgkiss, I.J. Karenia digitata sp. nov. (Gymnodiniales, Dinophyceae), a new harmful algal bloom species from the coastal waters of west Japan and Hong Kong. Phycologia 2000, 39, 463–470. [Google Scholar] [CrossRef]
- Cen, J.; Wang, J.; Huang, L.; Lin, Y.; Ding, G.; Qi, Y.; Lv, S. Karlodinium elegans sp. nov. (Gymnodiniales, Dinophyceae), a novel species isolated from the East China Sea in a dinoflagellate bloom. J. Oceanol. Limnol. 2020, 1–17. [Google Scholar] [CrossRef]
- Nezan, E.; Siano, R.; Boulben, S.; Six, C.; Bilien, G.; Cheze, K.; Duval, A.; Le Panse, S.; Quere, J.; Chomerat, N. Genetic diversity of the harmful family Kareniaceae (Gymnodiniales, Dinophyceae) in France, with the description of Karlodinium gentienii sp. nov.: A new potentially toxic dinoflagellate. Harmful Algae 2014, 40, 75–91. [Google Scholar] [CrossRef] [Green Version]
- Ballantine, D. Two new marine species of Gymnodinium isolated from the Plymouth area. J. Mar. Biol. Assoc. UK 1956, 35, 467–474. [Google Scholar] [CrossRef] [Green Version]
- Hallett, C.S.; Valesini, F.J.; Clarke, K.R.; Hoeksema, S.D. Effects of a harmful algal bloom on the community ecology, movements and spatial distributions of fishes in a microtidal estuary. Hydrobiologia 2016, 763, 267–284. [Google Scholar] [CrossRef]
- Granéli, E.; Turner, J.T. Ecology of Harmful Algae; Springer: Berlin/Heidelberg, Germany, 2006; pp. 1–413. [Google Scholar]
- Zhu, X.; Zhou, C.; Meng, R.; Li, S.; Fang, K.; Luo, Z.; Xu, J.; He, S.; Luo, Q.; Yan, X. Biochemical characteristics support the recently described species Karlodinium zhouanum (Gymnodiniales, Dinophyceae). Phycol. Res. 2019, 68, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Wang, L.; Chan, L.; Lu, S.; Gu, H. Karlodinium zhouanum, a new dinoflagellate species from China, and molecular phylogeny of Karenia digitata and Karenia longicanalis (Gymnodiniales, Dinophyceae). Phycologia 2018, 57, 401–412. [Google Scholar] [CrossRef]
- Li, Z.; Shin, H.H. Morphology and phylogeny of an unarmored dinoflagellate, Karlodinium jejuense sp. nov. (Gymnodiniales), isolated from the northern East China Sea. Phycol. Res. 2018, 66, 318–328. [Google Scholar] [CrossRef]
- Sheng, J.; Malkiel, E.; Katz, J.; Adolf, J.E.; Place, A.R. A dinoflagellate exploits toxins to immobilize prey prior to ingestion. Proc. Natl Acad. Sci. USA 2010, 107, 2082–2087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berge, T.; Hansen, P.J.; Moestrup, O. Feeding mechanism, prey specificity and growth in light and dark of the plastidic dinoflagellate Karlodinium armiger. Aquat. Microb. Ecol. 2008, 50, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Stoecker, D.K. Mixotrophy among dinoflagellates. J. Eukaryot. Microbiol. 1999, 46, 397–401. [Google Scholar] [CrossRef]
- Schnepf, E.; Elbrächter, M. Nutritional strategies in dinoflagellates: A review with emphasis on cell biological aspects. Eur. J. Protistol. 1992, 28, 3–24. [Google Scholar] [CrossRef]
- Biecheler, B. Recherches sur les Péridiniens. Bull. Biol. Fr. Belg. 1952, 36, 1–149. [Google Scholar]
- Calado, A.J.; Moestrup, Ø. Feeding in Peridiniopsis berolinensis (Dinophyceae): New observations on tube feeding by an omnivorous, heterotrophic dinoflagellate. Phycologia 1997, 36, 47–59. [Google Scholar] [CrossRef]
- Hansen, P.J.; Calado, A.J. Phagotrophic mechanisms and prey selection in free-living dinoflagellates. J. Eukaryot. Microbiol. 1999, 46, 382–389. [Google Scholar] [CrossRef]
- Jacobson, D.M.; Anderson, D.M. Thecate heterophic dinoflagellates: Feeding behavior and mechanisms. J. Phycol. 1986, 22, 249–258. [Google Scholar] [CrossRef]
- Jeong, H.J.; Latz, M.I. Growth and grazing rates of the heterotrophic dinoflagellates Protoperidinium spp. on red tide dinoflagellates. Mar. Ecol. Prog. Ser. 1994, 106, 173–185. [Google Scholar] [CrossRef]
- Elbrächter, M. Food uptake mechanisms in phagotrophic dinoflagellates and classification. In The Biology of Free-Living Heterotrophic Flagellates; Patterson, D.J., Larsen, J., Eds.; Clarendon Press: Oxford, UK, 1991; Volume 45, pp. 303–312. [Google Scholar]
- Schnepf, E.; Deichgräber, G. “Myzocytosis”, a kind of endocytosis with implications to compartmentation in endosymbiosis. Naturwissenschaften 1984, 71, 218–219. [Google Scholar] [CrossRef]
- Spero, H.J. Phagotrophy in Gymnodinium fungiforme (Pyrrhophyta): The peduncle as an organelle of ingestion. J. Phycol. 1982, 18, 356–360. [Google Scholar] [CrossRef]
- Wilcox, L.W.; Wedemayer, G.J. Phagotrophy in the freshwater, photosynthetic dinoflagellate Amphidinium cryophilum. J. Phycol. 1991, 27, 600–609. [Google Scholar] [CrossRef]
- Schnepf, E.; Deichgräber, G.; Drebes, G. Food uptake and the fine structure of the dinophyte Paulsenella sp., an ectoparasite of marine diatoms. Protoplasma 1985, 124, 188–204. [Google Scholar] [CrossRef]
- Place, A.R.; Bowers, H.A.; Bachvaroff, T.R.; Adolf, J.E.; Deeds, J.R.; Sheng, J. Karlodinium veneficum —The little dinoflagellate with a big bite. Harmful Algae 2012, 14, 179–195. [Google Scholar] [CrossRef]
- Song, X.; Hu, Z.; Shang, L.; Leaw, C.P.; Lim, P.T.; Tang, Y.Z. Contact micropredation may play a more important role than exotoxicity does in the lethal effects of Karlodinium australe blooms: Evidence from laboratory bioassays. Harmful Algae 2020, 99, 101926. [Google Scholar] [CrossRef]
- Vogelbein, W.K.; Lovko, V.J.; Shields, J.D.; Reece, K.S.; Mason, P.L.; Haas, L.W.; Walker, C.C. Pfiesteria shumwayae kills fish by micropredation not exotoxin secretion. Nature 2002, 418, 967–970. [Google Scholar] [CrossRef]
- Lafferty, K.D.; Kuris, A.M. Trophic strategies, animal diversity and body size. Trends Ecol. Evol. 2002, 17, 507–513. [Google Scholar] [CrossRef]
- Spero, H.J. Chemosensory capabilities in the phagotrophic dinoflagellate Gymnodinium fungiforme. J. Phycol. 1985, 21, 181–184. [Google Scholar] [CrossRef]
- Sheng, J.; Malkiel, E.; Katz, J.; Adolf, J.; Belas, R.; Place, A.R. Digital holographic microscopy reveals prey-induced changes in swimming behavior of predatory dinoflagellates. Proc. Natl Acad. Sci. USA 2007, 104, 17512–17517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, P.J. Prey size selection, feeding rates and growth dynamics of heterotrophic dinoflagellates with special emphasis on Gyrodinium-spirale. Mar. Biol. 1992, 114, 327–334. [Google Scholar] [CrossRef]
- Schnepf, E.; Winter, S.; Mollenhauer, D. Gymnodinium aeruginosum (Dinophyta): A blue-green dinoflagellate with a vestigial, anucleate, cryptophycean endosymbiont. Plant Syst. Evol. 1989, 164, 75–91. [Google Scholar] [CrossRef]
- Skovgaard, A. Role of chloroplast retention in a marine dinoflagellate. Aquat. Microb. Ecol. 1998, 15, 293–301. [Google Scholar] [CrossRef]
- Wilcox, L.W.; Wedemayer, G.J. Gymnodinium acidotum Nygaard (Pyrrophyta), a dinoflagellate with an endosymbiotic cryptomonad. J. Phycol. 2010, 20, 236–242. [Google Scholar] [CrossRef]
- Hansen, P.J. Phagotrophic mechanisms and prey selection in mixotrophic phytoflagellates. Physiol. Ecol. Harmful Algal Blooms 1998, 525–537. [Google Scholar]
- Jakobsen, H.H.; Hansen, P.J.; Larsen, J. Growth and grazing responses of two chloroplast-retaining dinoflagellates: Effect of irradiance and prey species. Mar. Ecol. Prog. Ser. 2000, 201, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Hansen, P.J. The role of photosynthesis and food uptake for the growth of marine mixotrophic dinoflagellates. J. Eukaryot. Microbiol. 2011, 58, 203–214. [Google Scholar] [CrossRef]
- Adolf, J.E.; Stoecker, D.K.; Harding, L.W., Jr. The balance of autotrophy and heterotrophy during mixotrophic growth of Karlodinium micrum (Dinophyceae). J. Plankton Res. 2006, 28, 737–751. [Google Scholar] [CrossRef]
- Adolf, J.E.; Bachvaroff, T.; Place, A.R. Can cryptophyte abundance trigger toxic Karlodinium veneficum blooms in eutrophic estuaries? Harmful Algae 2008, 8, 119–128. [Google Scholar] [CrossRef]
- Bachvaroff, T.R.; Adolf, J.E.; Place, A.R. Strain variation in Karlodinium veneficum (Dinophyceae): Toxin profiles, pigments, and growth characteristics. J. Phycol. 2009, 45, 137–153. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Hu, Z.; Xu, N.; Tang, Y.Z. A comparative study on the allelopathy and toxicity of four strains of Karlodinium veneficum with different culturing histories. J. Plankton Res. 2019, 41, 17–29. [Google Scholar] [CrossRef]
- Stoecker, D.K.; Gustafson, D.E. Cell-surface proteolytic activity of photosynthetic dinoflagellates. Aquat. Microb. Ecol. 2003, 30, 175–183. [Google Scholar] [CrossRef]
- Solomon, C.M.; Glibert, P.M. Urease activity in five phytoplankton species. Aquat. Microb. Ecol. 2008, 52, 149–157. [Google Scholar] [CrossRef]
- Croft, M.T.; Warren, M.J.; Smith, A.G. Algae need their vitamins. Eukaryot. Cell 2006, 5, 1175–1183. [Google Scholar] [CrossRef] [Green Version]
- Hellebust, J.A. Excretion of some organic compounds by marine phytoplankton. Limnol. Oceanogr. 1965, 10, 192–206. [Google Scholar] [CrossRef]
- Kamjunke, N.; Tittel, J. Utilisation of leucine by several phytoplankton species. Limnologica 2008, 38, 360–366. [Google Scholar] [CrossRef] [Green Version]
- Tittel, J.; Wiehle, I.; Wannicke, N.; Kampe, H.; Poerschmann, J.; Meier, J.; Kamjunke, N. Utilisation of terrestrial carbon by osmotrophic algae. Aquat. Sci. 2009, 71, 46–54. [Google Scholar] [CrossRef]
- Beamud, S.G.; Karrasch, B.; Pedrozo, F.L.; Diaz, M.M. Utilisation of organic compounds by osmotrophic algae in an acidic lake of Patagonia (Argentina). Limnology 2014, 15, 163–172. [Google Scholar] [CrossRef]
- Dąbrowska, A.; Nawrocki, J.; Szeląg-Wasielewska, E. Appearance of aldehydes in the surface layer of lake waters. Environ. Monit. Assess. 2014, 186, 4569–4580. [Google Scholar] [CrossRef] [Green Version]
- Nygaard, K.; Tobiesen, A. Bacterivory in algae—A survival strategy during nutrient limitation. Limnol. Oceanogr. 1993, 38, 273–279. [Google Scholar] [CrossRef]
- Li, A.S.; Stoecker, D.K.; Coats, D.W.; Adam, E.J. Ingestion of fluorescently labeled and phycoerythrin-containing prey by mixotrophic dinoflagellates. Aquat. Microb. Ecol. 1996, 10, 139–147. [Google Scholar] [CrossRef]
- Adolf, J.E.; Stoecker, D.K.; Harding, L.W. Autotrophic growth and photoacclimation in Karlodinium micrum (Dinophyceae) and Storeatula major (Cryptophyceae). J. Phycol. 2003, 39, 1101–1108. [Google Scholar] [CrossRef]
- Li, J.; Glibert, P.M.; Alexander, J.A.; Molina, M.E. Growth and competition of several harmful dinoflagellates under different nutrient and light conditions. Harmful Algae 2012, 13, 112–125. [Google Scholar] [CrossRef]
- Claessen, D.; de Roos, A.M.; Persson, L. Population dynamic theory of size-dependent cannibalism. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2004, 271, 333–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martel, C.M.; Flynn, K.J. Morphological controls on cannibalism in a planktonic marine phagotroph. Protist 2008, 159, 41–51. [Google Scholar] [CrossRef]
- Spero, H.J.; Moree, M.D. Phagotrophic feeding and its importance to the life cycle of the holozoic dinoflagellate, Gymnodinium fungiforme. J. Phycol. 1981, 17, 43–51. [Google Scholar] [CrossRef]
- Schnepf, E.; Drebes, G. Chemotaxis and appetence of Paulsenella sp. (Dinophyta), an ectoparasite of the marine diatom Streptotheca thamesis Shrubsole. Planta 1986, 167, 337–343. [Google Scholar] [CrossRef]
- Feinstein, T.N.; Traslavina, R.; Sun, M.Y.; Lin, S.J. Effects of light on photosynthesis, grazing, and population dynamics of the heterotrophic dinoflagellate Pfiesteria piscicida (Dinophyceae). J. Phycol. 2002, 38, 659–669. [Google Scholar] [CrossRef]
- Jeong, H.J.; Lee, C.W.; Yih, W.H.; Kim, J.S. Fragilidium cf mexicanum, a thecate mixotrophic dinoflagellate which is prey for and a predator on co-occurring thecate heterotrophic dinoflagellate Protoperidinium cf. divergens. Mar. Ecol. Prog. Ser. 1997, 151, 299–305. [Google Scholar]
- Naustvoll, L.J. Growth and grazing by the thecate heterotrophic dinoflagellate Diplopsalis lemicula (Diplopsalidaceae, Dinophyceae). Phycologia 1998, 37, 1–9. [Google Scholar] [CrossRef]
- Margulis, L. Origins of Sex; Yale University Press: New Haven, CT, USA, 1986. [Google Scholar]
- Raven, J.A.; Beardall, J.; Flynn, K.J.; Maberly, S.C. Phagotrophy in the origins of photosynthesis in eukaryotes and as a complementary mode of nutrition in phototrophs: Relation to Darwin’s insectivorous plants. J. Exp. Bot. 2009, 60, 3975–3987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leles, S.G.; Mitra, A.; Flynn, K.J.; Stoecker, D.K.; Hansen, P.J.; Calbet, A.; McManus, G.B.; Sanders, R.W.; Caron, D.A.; Not, F.; et al. Oceanic protists with different forms of acquired phototrophy display contrasting biogeographies and abundance. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2017, 284, 20170664. [Google Scholar] [CrossRef] [Green Version]
- Margulis, L.; Schwartz, K.V. Five Kingdoms; Freeman, W.H. and Company: San Francisco, CA, USA, 1982; pp. 1–338.
- Porter, K.G. Phagotrophic phytoflagellates in microbial food webs. Hydrobiologia 1988, 159, 89–97. [Google Scholar] [CrossRef]
- Stoecker, D.K.; Tillmann, U.; Granéli, E. Phagotrophy in harmful algae. In Ecology of Harmful Algae; Granéli, E., Turner, J.T., Eds.; Springer: Berlin, Germany, 2006; pp. 177–188. [Google Scholar]
- Mitra, A.; Flynn, K.J.; Tillmann, U.; Raven, J.A.; Caron, D.; Stoecker, D.K.; Not, F.; Hansen, P.J.; Hallegraeff, G.; Sanders, R.; et al. Defining planktonic protist functional groups on mechanisms for energy and nutrient acquisition: Incorporation of diverse mixotrophic strategies. Protist 2016, 167, 106–120. [Google Scholar] [CrossRef] [PubMed]
- Stoecker, D.K.; Hansen, P.J.; Caron, D.A.; Mitra, A.; Annual, R. Mixotrophy in the marine plankton. Ann. Rev. Mar. Sci. 2017, 9, 311–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, A.; Flynn, K.J.; Burkholder, J.M.; Berge, T.; Calbet, A.; Raven, J.A.; Graneli, E.; Glibert, P.M.; Hansen, P.J.; Stoecker, D.K.; et al. The role of mixotrophic protists in the biological carbon pump. Biogeosciences 2014, 11, 995–1005. [Google Scholar] [CrossRef] [Green Version]
- Flynn, K.J.; Mitra, A.; Anestis, K.; Anschütz, A.A.; Calbet, A.; Ferreira, G.D.; Gypens, N.; Hansen, P.J.; John, U.; Martin, J.L.; et al. Mixotrophic protists and a new paradigm for marine ecology: Where does plankton research go now? J. Plankton Res. 2019, 4, 375–391. [Google Scholar] [CrossRef] [Green Version]
- Legrand, C.; Granéli, E.; Carlsson, P. Induced phagotrophy in the photosynthetic dinoflagellate Heterocapsa triquetra. Aquat. Microb. Ecol. 1998, 15, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Berge, T.; Hansen, P.J. Role of the chloroplast in the predatory dinoflagellate Karlodinium armiger. Mar. Ecol. Prog. 2016, 519, 41–54. [Google Scholar] [CrossRef]
- Naik, R.K.; Chitari, R.R.; Anil, A.C. Karlodinium veneficum in India: Effect of fixatives on morphology and allelopathy in relation to Skeletonema costatum. Curr. Sci. India. 2010, 99, 1112–1116. [Google Scholar]
- Tittel, J.; Bissinger, V.; Zippel, B.; Gaedke, U.; Bell, E.; Lorke, A.; Kamjunke, N. Mixotrophs combine resource use to outcompete specialists: Implications for aquatic food webs. Proc. Natl Acad. Sci. USA 2003, 100, 12776–12781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granéli, E.; Edvardsen, B.; Roelke, D.L.; Hagström, J.A. The ecophysiology and bloom dynamics of Prymnesium spp. Harmful Algae 2012, 14, 260–270. [Google Scholar] [CrossRef]
- Cabrerizo, M.J.; Manuel Gonzalez-Olalla, J.; Hinojosa-Lopez, V.J.; Peralta-Cornejo, F.J.; Carrillo, P. A shifting balance: Responses of mixotrophic marine algae to cooling and warming under UVR. N. Phytol. 2019, 221, 1317–1327. [Google Scholar] [CrossRef] [Green Version]
- Okada, M.; Huston, C.D.; Mann, B.J.; Petri, W.A.; Kita, K.; Nozaki, T. Proteomic analysis of phagocytosis in the enteric protozoan parasite Entamoeba histolytica. Eukaryot.Cell 2005, 4, 827–831. [Google Scholar] [CrossRef] [Green Version]
- Gotthardt, D.; Blancheteau, V.; Bosserhoff, A.; Ruppert, T.; Delorenzi, M.; Soldati, T. Proteomics fingerprinting of phagosome maturation and evidence for the role of a G alpha during uptake. Mol. Cell. Proteomics 2006, 5, 2228–2243. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, M.E.; DeSouza, L.V.; Samaranayake, H.; Pearlman, R.E.; Siu, K.W.M.; Klobutcher, L.A. The Tetrahymena thermophila phagosome proteome. Eukaryot. Cell 2006, 5, 1990–2000. [Google Scholar] [CrossRef] [Green Version]
- Shevchuk, O.; Batzilla, C.; Haegele, S.; Kusch, H.; Engelmann, S.; Hecker, M.; Haas, A.; Heuner, K.; Gloeckner, G.; Steinert, M. Proteomic analysis of Legionella-containing phagosomes isolated from Dicyostelium. Int. J. Med. Microbiol. 2009, 299, 489–508. [Google Scholar] [CrossRef]
- Boulais, J.; Trost, M.; Landry, C.R.; Dieckmann, R.; Levy, E.D.; Soldati, T.; Michnick, S.W.; Thibault, P.; Desjardins, M. Molecular characterization of the evolution of phagosomes. Mol. Syst. Biol. 2010, 6, 423. [Google Scholar] [CrossRef]
- Burns, J.A.; Paasch, A.; Narechania, A.; Kim, E. Comparative genomics of a bacterivorous green alga reveals evolutionary causalities and consequences of phago-mixotrophic mode of nutrition. Genome Biol. Evol. 2015, 7, 3047–3061. [Google Scholar] [CrossRef] [Green Version]
- Lie, A.A.Y.; Liu, Z.; Terrado, R.; Tatters, A.O.; Heidelberg, K.B.; Caron, D.A. Effect of light and prey availability on gene expression of the mixotrophic chrysophyte, Ochromonas sp. BMC Genomics 2017, 18, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]



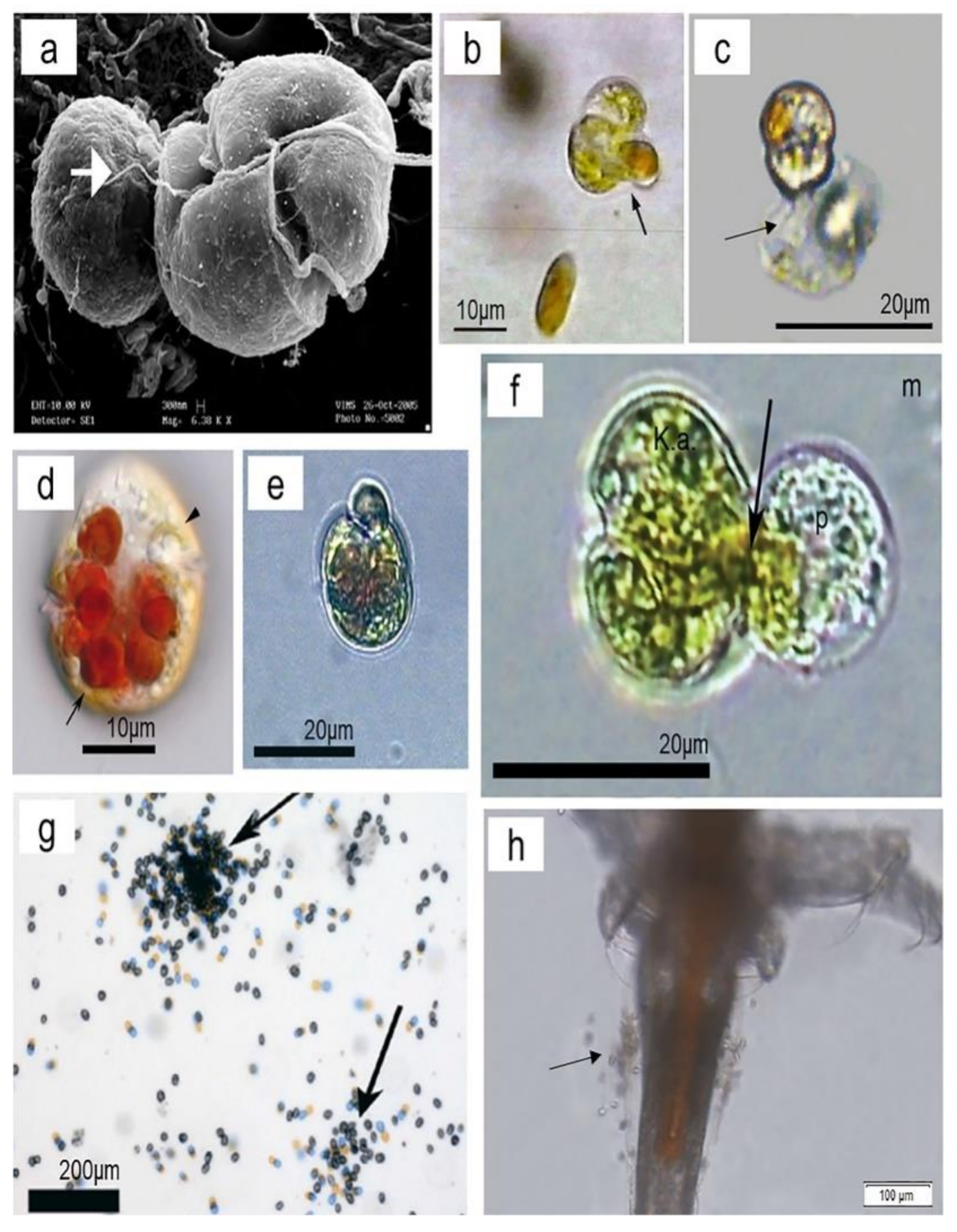{kind=link}
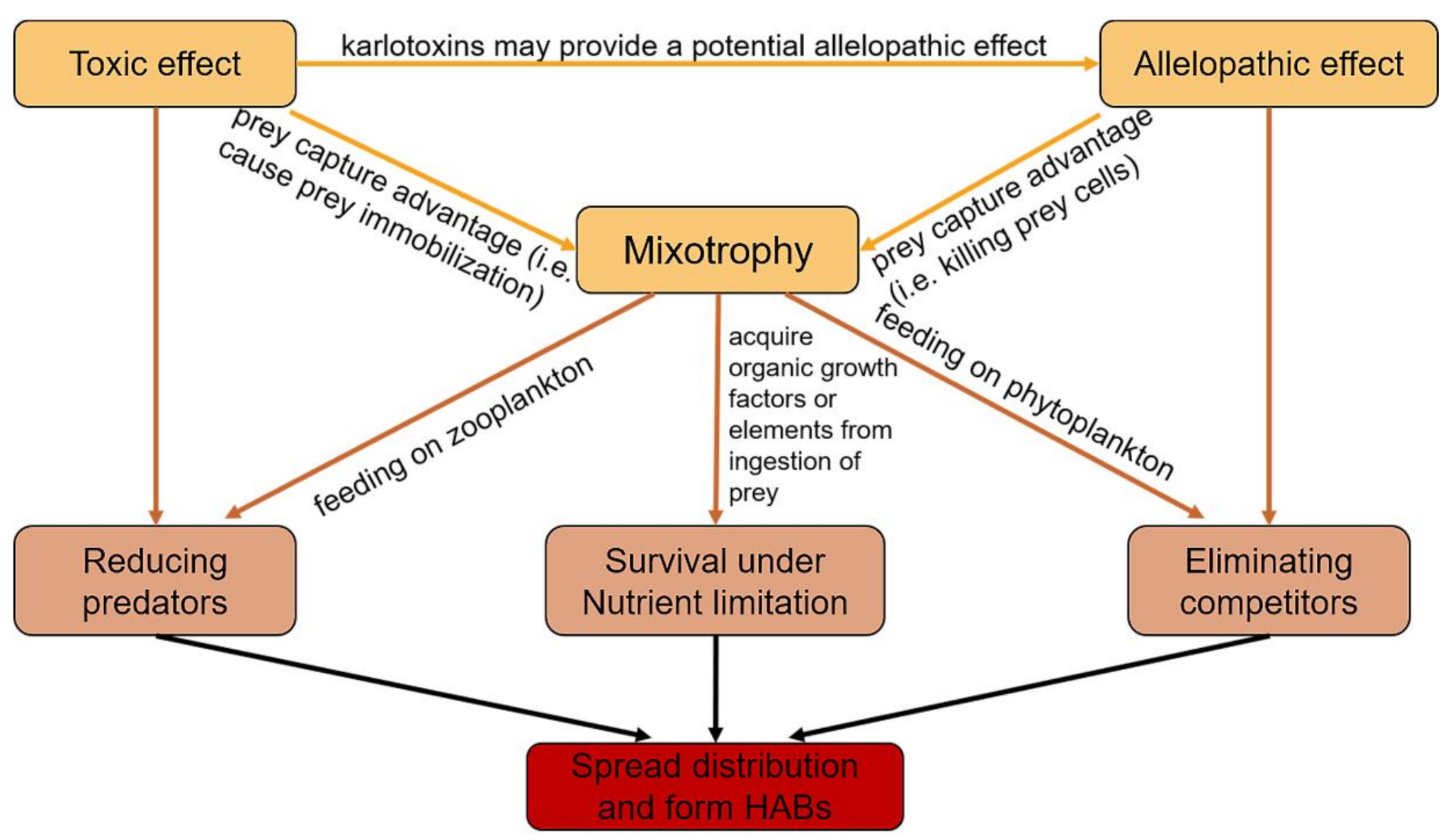{kind=link}
| Species | Former names and/or taxonomic synonyms | Distribution | Blooms | Toxicity | Autotrophy | Osmotrophy | Phagotrophy | Peduncle-like structure |
|---|---|---|---|---|---|---|---|---|
| K. armiger | \ | Alfacs Bay, Ebro Delta, NW Mediterranean [38] | Yes [29,39] | Yes (Karmitoxin) [37] | Yes | ? | Yes [40,41] | Have a peduncle [38] |
| K. australe | \ | North-eastern Tasmania, Port Phillip Bay (Victoria), South Australia and TuggerahLakes [28] and Singapore [42,43] | Yes [18] | Yes [18,28] | Yes [28] | ? | Yes [28] | Have a thick, tubular peduncle-like structure [28] |
| K. antarcticum | \ | Southern Ocean [43] | ? | No [34] | Yes [43] | ? | ? | Have a tube-shaped structure [43] |
| K. azanzae | \ | Manila Bay, Philippines [44] | ? | Yes [44] | Yes [44] | ? | Yes [44] | Have a peduncle [44] |
| K. ballantinum | \ | Mercury Passage, Tasmania, Australia, and Tyrrhenian coastal waters [43] and the Mexican Pacific [45] | ? | ? | Yes | ? | ? | Have a tube-shaped structure [43] |
| K. conicum | \ | Southern Ocean [43] | ? | Yes (KmTx) [34] | Yes | ? | ? | Have a tube-shaped structure [43] |
| K. corrugatum | \ | Southern Ocean [43] | ? | No [34] | Yes | ? | ? | ? |
| K. corsicum | Gyrodinium corsicum | Corsica (France), Tyrrhenian Sea and the Spanish Alfacs Bay of Mediterranean Sea [29,46] | Yes [24] | Yes [26] | Yes | ? | ? | Have a ventral plate [47] |
| K. decipiens | Karenia digitate | from coastal Tasmania southward to the north polar front, and western European Atlantic waters (Bilbao, Spain) [43] | ? | No [34] | Yes | ? | ? | Have a tube-shaped structure [43] |
| K. digitatum1 | Karenia digitata | Japan coastal waters of Hong Kong, Fujian and Guangdong’s Southern, China [48,49] | Yes [49] | Yes [49] | Yes | ? | ? | Have small finger-like extensions [49] |
| K. elegans | \ | Pingtan coastal water, East China Sea [50] | ? | No [50] | Yes [50] | ? | ? | Have a tube-like structure [50] |
| K. gentienii | \ | The Atlantic coast of Brittany [51] | Yes [51] | Yes [51] | Yes | ? | ? | Have a tube-shaped structure [51] |
| K. veneficum | Gymnodinium veneficum; Karlodinium micrum; Gymnodinium micrum; Gyrodinium galatheanum; Woloszynskia micra; Gyrodinium estuariale [38,52] | Cosmopolitan [16] | Yes [29,39,53] | Yes [54] | Yes [20] | Yes | Yes [16,20,21] | Have a peduncle [38] |
| K. vitiligo | Gymnodinium vitiligo ? K. veneficum [38] | ? | ? | ? | Yes | ? | ? | ? |
| K. zhouanum | K. jejuense | Widely spread over the coastal waters of China [55,56] | Possible [57] | ? | Yes [56] | ? | ? | Tube-like structure in intercingular region [56] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Hu, Z.; Tang, Y.Z. Plasticity and Multiplicity of Trophic Modes in the Dinoflagellate Karlodinium and Their Pertinence to Population Maintenance and Bloom Dynamics. J. Mar. Sci. Eng. 2021, 9, 51. https://doi.org/10.3390/jmse9010051
Yang H, Hu Z, Tang YZ. Plasticity and Multiplicity of Trophic Modes in the Dinoflagellate Karlodinium and Their Pertinence to Population Maintenance and Bloom Dynamics. Journal of Marine Science and Engineering. 2021; 9(1):51. https://doi.org/10.3390/jmse9010051
Chicago/Turabian StyleYang, Huijiao, Zhangxi Hu, and Ying Zhong Tang. 2021. "Plasticity and Multiplicity of Trophic Modes in the Dinoflagellate Karlodinium and Their Pertinence to Population Maintenance and Bloom Dynamics" Journal of Marine Science and Engineering 9, no. 1: 51. https://doi.org/10.3390/jmse9010051
APA StyleYang, H., Hu, Z., & Tang, Y. Z. (2021). Plasticity and Multiplicity of Trophic Modes in the Dinoflagellate Karlodinium and Their Pertinence to Population Maintenance and Bloom Dynamics. Journal of Marine Science and Engineering, 9(1), 51. https://doi.org/10.3390/jmse9010051